SIG1, a Sigma Factor for the Chloroplast RNA Polymerase, Differently Associates with Multiple DNA Regions in the Chloroplast Chromosomes in Vivo

Abstract
:1. Introduction
2. Results
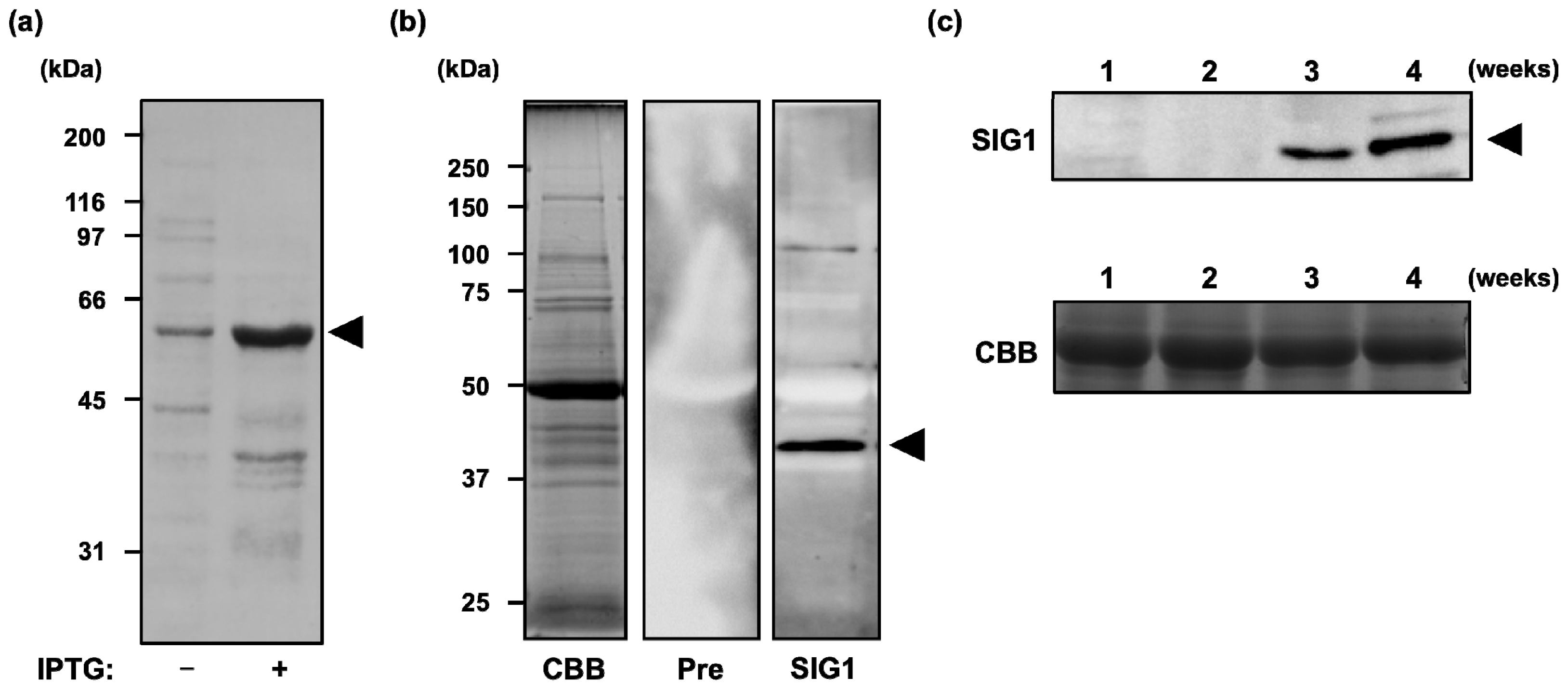
2.1. Accumulation of SIG1 Protein during Development of A. thaliana Seedlings
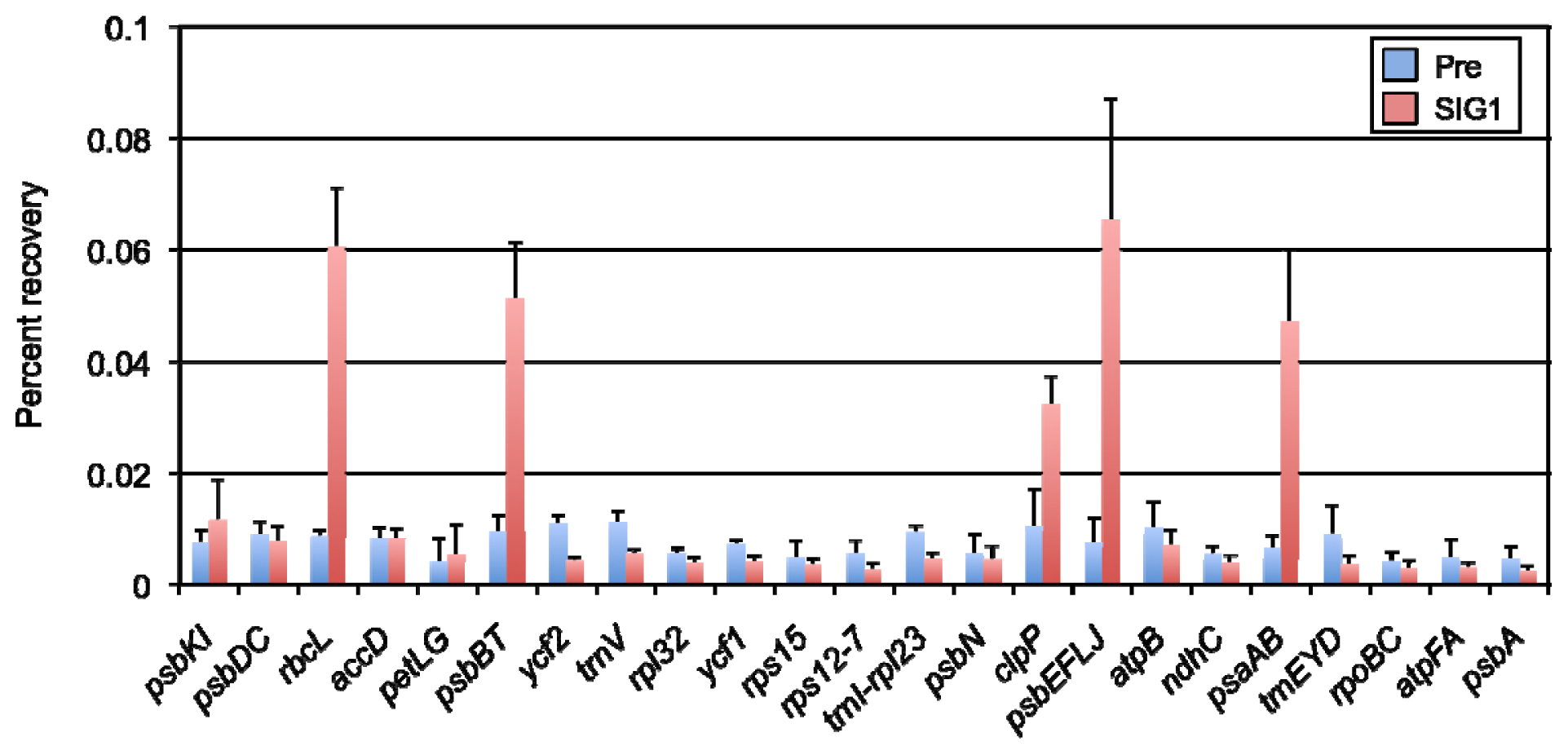
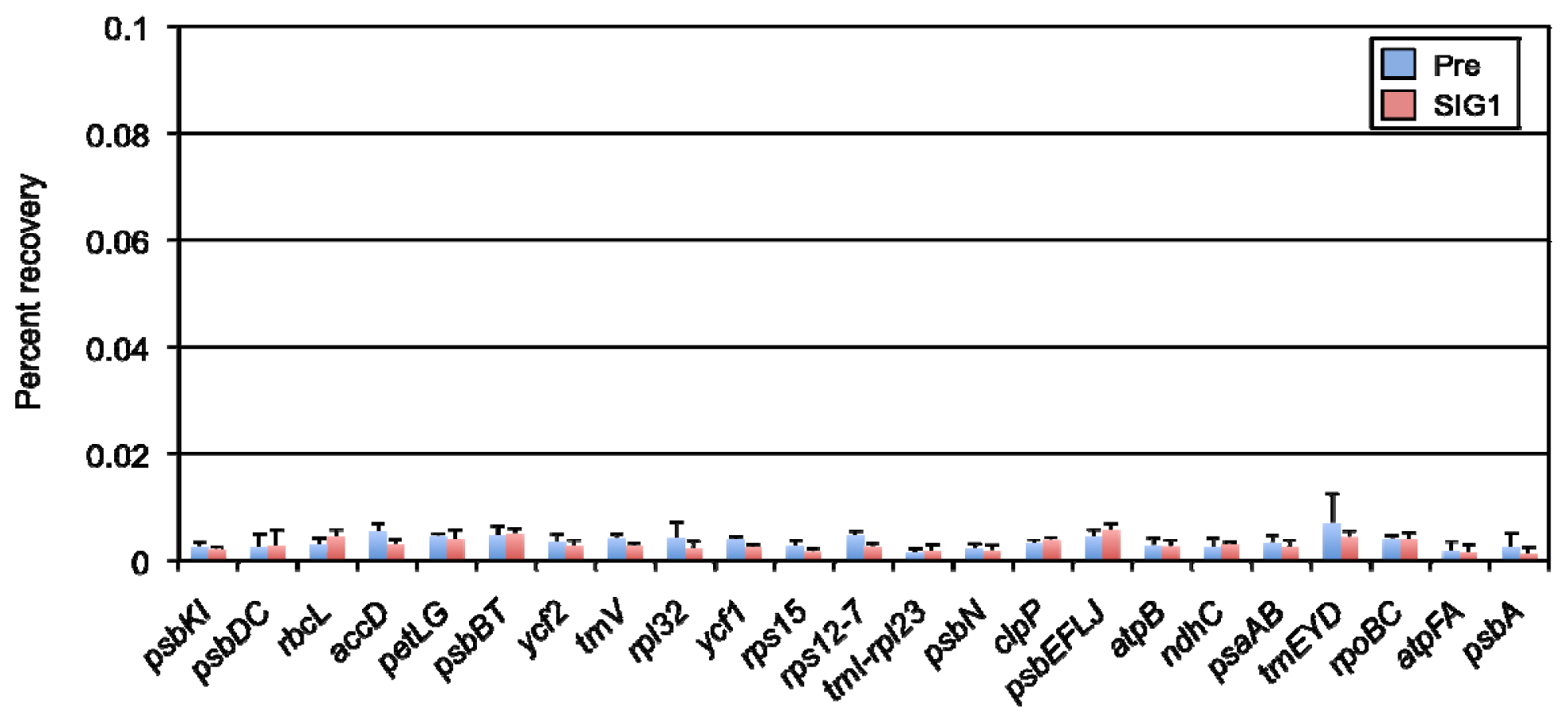
2.2. Identification of SIG1-Dependent Promoters in A. thaliana Chloroplasts
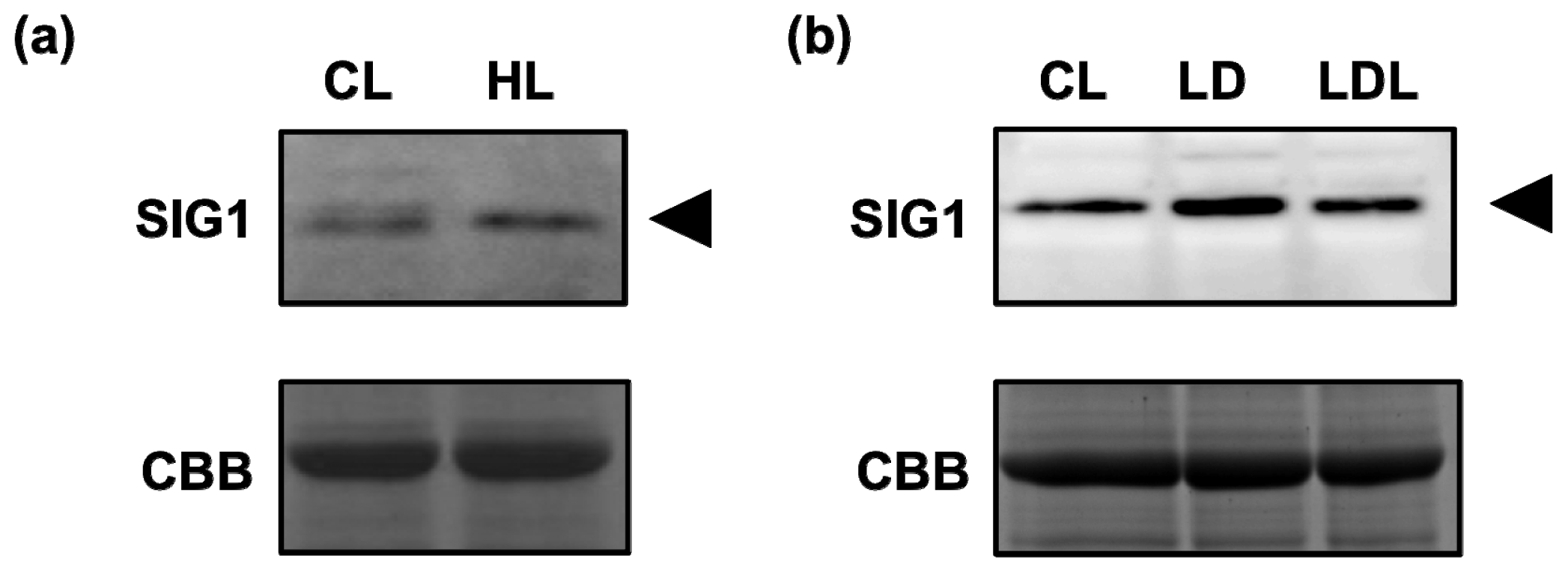
2.3. High-Light-Dependent Release of SIG1 from Its Target Promoters
2.4. Accumulation of SIG1 during the Change in Light Conditions
3. Discussion
4. Experimental Section
4.1. Plant Materials and Growth Conditions
4.2. Preparation of Polyclonal Antibody and Immunoblotting
4.3. Preparation of Leaf Extracts for ChIP Analysis
4.4. ChIP and Quantitative PCR Analysis
5. Conclusion
Acknowledgments
References
- Sugita, M.; Sugiura, M. Regulation of gene expression in chloroplasts of higher plants. Plant Mol. Biol 1996, 32, 315–326. [Google Scholar]
- Sato, S.; Nakamura, Y.; Kaneko, T.; Asamizu, E.; Tabata, S. Complete structure of the chloroplast genome of Arabidopsis thaliana. DNA Res 1999, 6, 283–290. [Google Scholar]
- Leister, D. Chloroplast research in the genomic age. Trends Genet 2003, 19, 47–56. [Google Scholar]
- Goldschmidt-Clermont, M. Coordination of nuclear and chloroplast gene expression in plant cells. Int. Rev. Cytol 1998, 177, 115–180. [Google Scholar]
- Hess, W.R.; Börner, T. Organellar RNA polymerases of higher plants. Int. Rev. Cytol 1999, 190, 1–59. [Google Scholar]
- Shiina, T.; Tsunoyama, Y.; Nakahira, Y.; Khan, M.S. Plastid RNA polymerases, promoters, and transcription regulators in higher plants. Int. Rev. Cytol 2005, 244, 1–68. [Google Scholar]
- Hedtke, B.; Börner, T.; Weihe, A. Mitochondrial and chloroplast phage-type RNA polymerases in Arabidopsis. Science 1997, 277, 809–811. [Google Scholar]
- Hajdukiewicz, P.T.; Allison, L.A.; Maliga, P. The two RNA polymerases encoded by the nuclear and the plastid compartments transcribe distinct groups of genes in tobacco plastids. EMBO J 1997, 16, 4041–4048. [Google Scholar]
- Allison, L.A.; Simon, L.D.; Maliga, P. Deletion of rpoB reveals a second distinct transcription system in plastids of higher plants. EMBO J 1996, 15, 2802–2809. [Google Scholar]
- De Santis-Maciossek, G.; Kofer, W.; Bock, A.; Schoch, S.; Maier, R.M.; Wanner, G.; Rudiger, W.; Koop, H.U.; Herrmann, R.G. Targeted disruption of the plastid RNA polymerase genes rpoA, B and C1: molecular biology, biochemistry and ultrastructure. Plant J 1999, 18, 477–489. [Google Scholar]
- Allison, L.A. The role of sigma factors in plastid transcription. Biochimie 2000, 82, 537–548. [Google Scholar]
- Isono, K.; Shimizu, M.; Yoshimoto, K.; Niwa, Y.; Satoh, K.; Yokota, A.; Kobayashi, H. Leaf-specifically expressed genes for polypeptides destined for chloroplasts with domains of sigma70 factors of bacterial RNA polymerases in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1997, 94, 14948–14953. [Google Scholar]
- Tanaka, K.; Tozawa, Y.; Mochizuki, N.; Shinozaki, K.; Nagatani, A.; Wakasa, K.; Takahashi, H. Characterization of three cDNA species encoding plastid RNA polymerase sigma factors in Arabidopsis thaliana: Evidence for the sigma factor heterogeneity in higher plant plastids. FEBS Lett 1997, 413, 309–313. [Google Scholar]
- Fujiwara, M.; Nagashima, A.; Kanamaru, K.; Tanaka, K.; Takahashi, H. Three new nuclear genes, sigD, sigE and sigF, encoding putative plastid RNA polymerase sigma factors in Arabidopsis thaliana. FEBS Lett 2000, 481, 47–52. [Google Scholar]
- Lysenko, E.A. Plant sigma factors and their role in plastid transcription. Plant Cell Rep 2007, 26, 845–859. [Google Scholar]
- Schweer, J.; Türkeri, H.; Kolpack, A.; Link, G. Role and regulation of plastid sigma factors and their functional interactors during chloroplast transcription—Recent lessons from Arabidopsis thaliana. Eur. J. Cell Biol 2010, 89, 940–946. [Google Scholar]
- Kestermann, M.; Neukirchen, S.; Kloppstech, K.; Link, G. Sequence and expression characteristics of a nuclear-encoded chloroplast sigma factor from mustard (Sinapis alba). Nucleic Acids Res 1998, 26, 2747–2753. [Google Scholar]
- Privat, I.; Hakimi, M.A.; Buhot, L.; Favory, J.J.; Lerbs-Mache, S. Characterization of Arabidopsis plastid sigma-like transcription factors SIG1, SIG2 and SIG3. Plant Mol. Biol 2003, 51, 385–399. [Google Scholar]
- Zghidi, W.; Merendino, L.; Cottet, A.; Mache, R.; Lerbs-Mache, S. Nucleus-encoded plastid sigma factor SIG3 transcribes specifically the psbN gene in plastids. Nucleic Acids Res 2007, 35, 455–464. [Google Scholar]
- Favory, J.J.; Kobayshi, M.; Tanaka, K.; Peltier, G.; Kreis, M.; Valay, J.G.; Lerbs-Mache, S. Specific function of a plastid sigma factor for ndhF gene transcription. Nucleic Acids Res 2005, 33, 5991–5999. [Google Scholar]
- Nagashima, A.; Hanaoka, M.; Shikanai, T.; Fujiwara, M.; Kanamaru, K.; Takahashi, H.; Tanaka, K. The multiple-stress responsive plastid sigma factor, SIG5, directs activation of the psbD blue light-responsive promoter (BLRP) in Arabidopsis thaliana. Plant Cell Physiol 2004, 45, 357–368. [Google Scholar]
- Kanamaru, K.; Nagashima, A.; Fujiwara, M.; Shimada, H.; Shirano, Y.; Nakabayashi, K.; Shibata, D.; Tanaka, K.; Takahashi, H. An Arabidopsis sigma factor (SIG2)-dependent expression of plastid-encoded tRNAs in chloroplasts. Plant Cell Physiol 2001, 42, 1034–1043. [Google Scholar]
- Ishizaki, Y.; Tsunoyama, Y.; Hatano, K.; Ando, K.; Kato, K.; Shinmyo, A.; Kobori, M.; Takeba, G.; Nakahira, Y.; Shiina, T. A nuclear-encoded sigma factor, Arabidopsis SIG6, recognizes sigma-70 type chloroplast promoters and regulates early chloroplast development in cotyledons. Plant J 2005, 42, 133–144. [Google Scholar]
- Hanaoka, M.; Kanamaru, K.; Takahashi, H.; Tanaka, K. Molecular genetic analysis of chloroplast gene promoters dependent on SIG2, a nucleus-encoded sigma factor for the plastid-encoded RNA polymerase, in Arabidopsis thaliana. Nucleic Acids Res 2003, 31, 7090–7098. [Google Scholar]
- Nagashima, A.; Hanaoka, M.; Motohashi, R.; Seki, M.; Shinozaki, K.; Kanamaru, K.; Takahashi, H.; Tanaka, K. DNA microarray analysis of plastid gene expression in an Arabidopsis mutant deficient in a plastid transcription factor sigma, SIG2. Biosci. Biotechnol. Biochem 2004, 68, 694–704. [Google Scholar]
- Loschelder, H.; Schweer, J.; Link, B.; Link, G. Dual temporal role of plastid sigma factor 6 in Arabidopsis development. Plant Physiol 2006, 142, 642–650. [Google Scholar]
- Tsunoyama, Y.; Ishizaki, Y.; Morikawa, K.; Kobori, M.; Nakahira, Y.; Takeba, G.; Toyoshima, Y.; Shiina, T. Blue light-induced transcription of plastid-encoded psbD gene is mediated by a nuclear-encoded transcription initiation factor, AtSig5. Proc. Natl. Acad. Sci. USA 2004, 101, 3304–3309. [Google Scholar]
- Aparicio, O.M.; Geisberg, J.V.; Struhl, K. Chromatin Immunoprecipitation for Determining the Association of Proteins with Specific Genomic Sequences in vivo. In Current Protocols in Molecular Biology; Ausubel, F.A., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., Struhl, K., Eds.; John Wiley & Sons: New York, NY, USA, 2004; pp. 21.3.1–21.3.32. [Google Scholar]
- Kuo, M.H.; Allis, C.D. In vivo cross-linking and immunoprecipitation for studying dynamic Protein: DNA associations in a chromatin environment. Methods 1999, 19, 425–433. [Google Scholar]
- Prikryl, J.; Watkins, K.P.; Friso, G.; van Wijk, K.J.; Barkan, A. A member of the Whirly family is a multifunctional RNA- and DNA-binding protein that is essential for chloroplast biogenesis. Nucleic Acids Res 2008, 36, 5152–5165. [Google Scholar]
- Newell, C.A.; Gray, J.C. Binding of lac repressor-GFP fusion protein to lac operator sites inserted in the tobacco chloroplast genome examined by chromatin immunoprecipitation. Nucleic Acids Res 2010, 38, e145. [Google Scholar]
- Yagi, Y.; Ishizaki, Y.; Nakahira, Y.; Tozawa, Y.; Shiina, T. Eukaryotic-type plastid nucleoid protein pTAC3 is essential for transcription by the bacterial-type plastid RNA polymerase. Proc. Natl. Acad. Sci. USA 2012, 109, 7541–7546. [Google Scholar]
- Lahiri, S.D.; Allison, L.A. Complementary expression of two plastid-localized sigma-like factors in maize. Plant Physiol 2000, 123, 883–894. [Google Scholar]
- Tozawa, Y.; Teraishi, M.; Sasaki, T.; Sonoike, K.; Nishiyama, Y.; Itaya, M.; Miyao, A.; Hirochika, H. The plastid sigma factor SIG1 maintains photosystem I activity via regulated expression of the psaA operon in rice chloroplasts. Plant J 2007, 52, 124–132. [Google Scholar]
- Hakimi, M.A.; Privat, I.; Valay, J.G.; Lerbs-Mache, S. Evolutionary conservation of C-terminal domains of primary sigma(70)-type transcription factors between plants and bacteria. J. Biol. Chem 2000, 275, 9215–9221. [Google Scholar]
- Zoschke, R.; Liere, K.; Börner, T. From seedling to mature plant: Arabidopsis plastidial genome copy number, RNA accumulation and transcription are differentially regulated during leaf development. Plant J 2007, 50, 710–722. [Google Scholar]
- Shikanai, T.; Shimizu, K.; Ueda, K.; Nishimura, Y.; Kuroiwa, T.; Hashimoto, T. The chloroplast clpP gene, encoding a proteolytic subunit of ATP-dependent protease, is indispensable for chloroplast development in tobacco. Plant Cell Physiol 2001, 42, 264–273. [Google Scholar]
- Schweer, J.; Geimer, S.; Meurer, J.; Link, G. Arabidopsis mutants carrying chimeric sigma factor genes reveal regulatory determinants for plastid gene expression. Plant Cell Physiol 2009, 50, 1382–1386. [Google Scholar]
- Schweer, J.; Türkeri, H.; Link, B.; Link, G. AtSIG6, a plastid sigma factor from Arabidopsis, reveals functional impact of cpCK2 phosphorylation. Plant J 2010, 62, 192–202. [Google Scholar]
- Shimizu, M.; Kato, H.; Ogawa, T.; Kurachi, A.; Nakagawa, Y.; Kobayashi, H. Sigma factor phosphorylation in the photosynthetic control of photosystem stoichiometry. Proc. Natl. Acad. Sci. USA 2010, 107, 10760–10764. [Google Scholar]
- Türkeri, H.; Schweer, J.; Link, G. Phylogenetic and functional features of the plastid transcription kinase cpCK2 from Arabidopsis signify a role of cysteinyl SH-groups in regulatory phosphorylation of plastid sigma factors. FEBS J 2012, 279, 395–409. [Google Scholar]
- Johnson, C.; Boden, E.; Arias, J. Salicylic acid and NPR1 induce the recruitment of trans-activating TGA factors to a defense gene promoter in Arabidopsis. Plant Cell 2003, 15, 1846–1858. [Google Scholar]
- Hanaoka, M.; Tanaka, K. Dynamics of RpaB-promoter interaction during high light stress, revealed by chromatin immunoprecipitation (ChIP) analysis in Synechococcus elongatus PCC 7942. Plant J 2008, 56, 327–335. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward (5′ to 3′) | Reverse (5′ to 3′) | |
|---|---|---|
| psbKI | TTGATCATTACATAGAAT | AACAAAAATTGGTGTTCT |
| psbDC | AATAAAATCAAAAATTTTG | AGCGATCCTCCTATTCA |
| rbcL | ATGAAAGAATATACAATAA | AAGTCCCTCCCTACAAG |
| accD | ATCCTTCTTTTCATTTAG | AGAGCTTCTGGCCTCTA |
| petLG | TGAATTGAGTTCTTTTTA | GAAGGGACTCAATAAAA |
| psbBT | TTGGTACTTATCGGATAT | GGAAATACCCCTTTATCA |
| ycf2 | GCCAATTCCAATAGACTT | TGATTCCTCCTAAATTGC |
| trnV-16S rRNA | ATGGCTCGAATCCGTAGT | TCCCCCATCAAGAAATAG |
| rpl32 | ATTATTTAAATGAGTACT | TCAAAAATGAAAAAAAAT |
| ycf1 | TTTAATAGGGAACCTCAA | AAACCTCCCTTTTTTCTT |
| rps15 | GATACCAATTATAGCGGA | AAAAAGAAATCCTTCCCC |
| rps12-7 | GTATGGATATGTAAAATACA | TTGTAGGGTGGATCTCG |
| trnI-rpl23 | ATCCCACTGAATTGAATTG | TTAGTGGGGATCCTCGT |
| psbN | TTTACCATATTCGGAATT | TATTATAGAATTGAAAGA |
| clpP | TAGTTTTATTCATTCTCT | GAAATGAAAAAAAAAGAG |
| psbEFLJ | ATTATGTAACACCCCATT | ACTGAACTCCAGATATTC |
| atpB | AGGTTTCAGTTAGTTGA | AATAAAAAAAATATGTTAAA |
| ndhC | CTATTAAGTAATAAGTGTA | AGACGAACTCCTATGAA |
| psaAB | CATAATAGATCCGAACACT | TGAGTCCTCCTCTTTCC |
| trnEYD | AATATAAAAAGAAAGTATAT | ATACTTGCTCAACCGC |
| rpoBC | TTCCAATTGAATATAGTC | CTTTTTTGAATTTCCCAT |
| atpFA | ATAAGTCTCATTATTATTA | ATAATCTCCTCTTCTAG |
| psbA | GTGGATTCGCTTCTAATT | GGTAAAATCCTTGGTTTA |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hanaoka, M.; Kato, M.; Anma, M.; Tanaka, K. SIG1, a Sigma Factor for the Chloroplast RNA Polymerase, Differently Associates with Multiple DNA Regions in the Chloroplast Chromosomes in Vivo. Int. J. Mol. Sci. 2012, 13, 12182-12194. https://doi.org/10.3390/ijms131012182
Hanaoka M, Kato M, Anma M, Tanaka K. SIG1, a Sigma Factor for the Chloroplast RNA Polymerase, Differently Associates with Multiple DNA Regions in the Chloroplast Chromosomes in Vivo. International Journal of Molecular Sciences. 2012; 13(10):12182-12194. https://doi.org/10.3390/ijms131012182
Chicago/Turabian StyleHanaoka, Mitsumasa, Maiko Kato, Misato Anma, and Kan Tanaka. 2012. "SIG1, a Sigma Factor for the Chloroplast RNA Polymerase, Differently Associates with Multiple DNA Regions in the Chloroplast Chromosomes in Vivo" International Journal of Molecular Sciences 13, no. 10: 12182-12194. https://doi.org/10.3390/ijms131012182