The Expression Profiling of the Lipoxygenase (LOX) Family Genes During Fruit Development, Abiotic Stress and Hormonal Treatments in Cucumber (Cucumis sativus L.)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Expression Profiling of the Cucumber LOX Genes During Vegetative and Reproductive Development
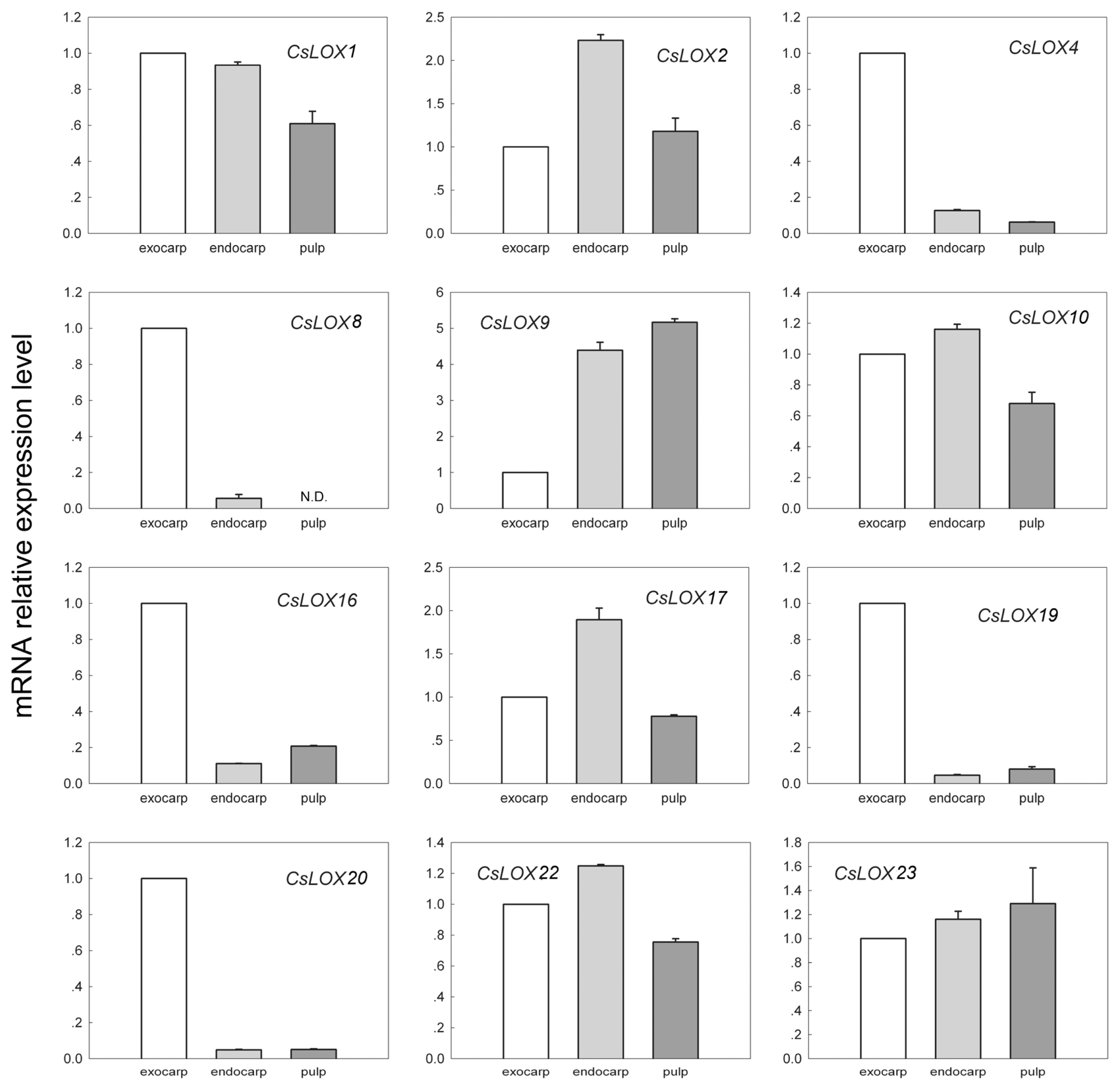
2.2. The Expression Patterns of Cucumber LOX During Fruit Development
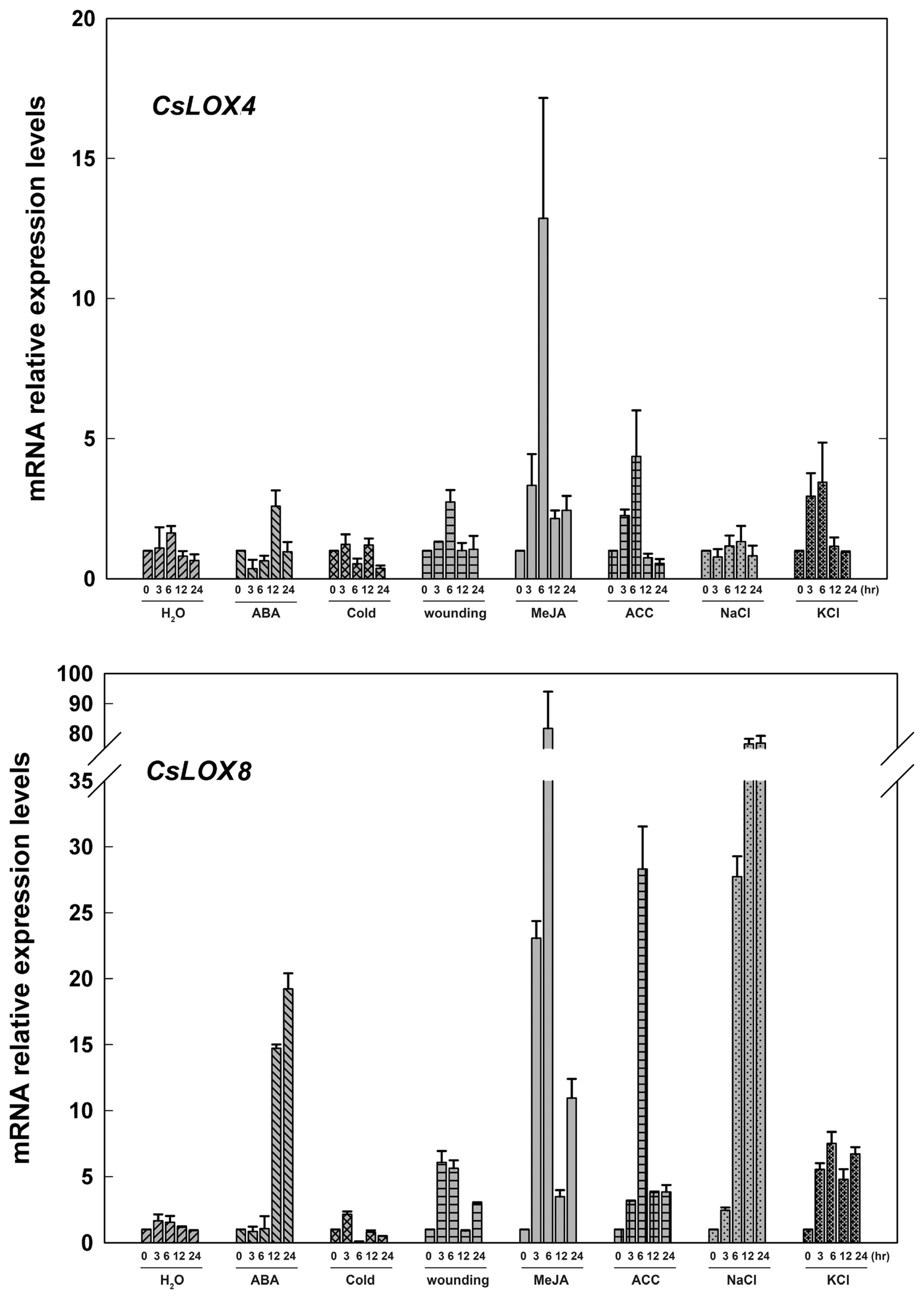
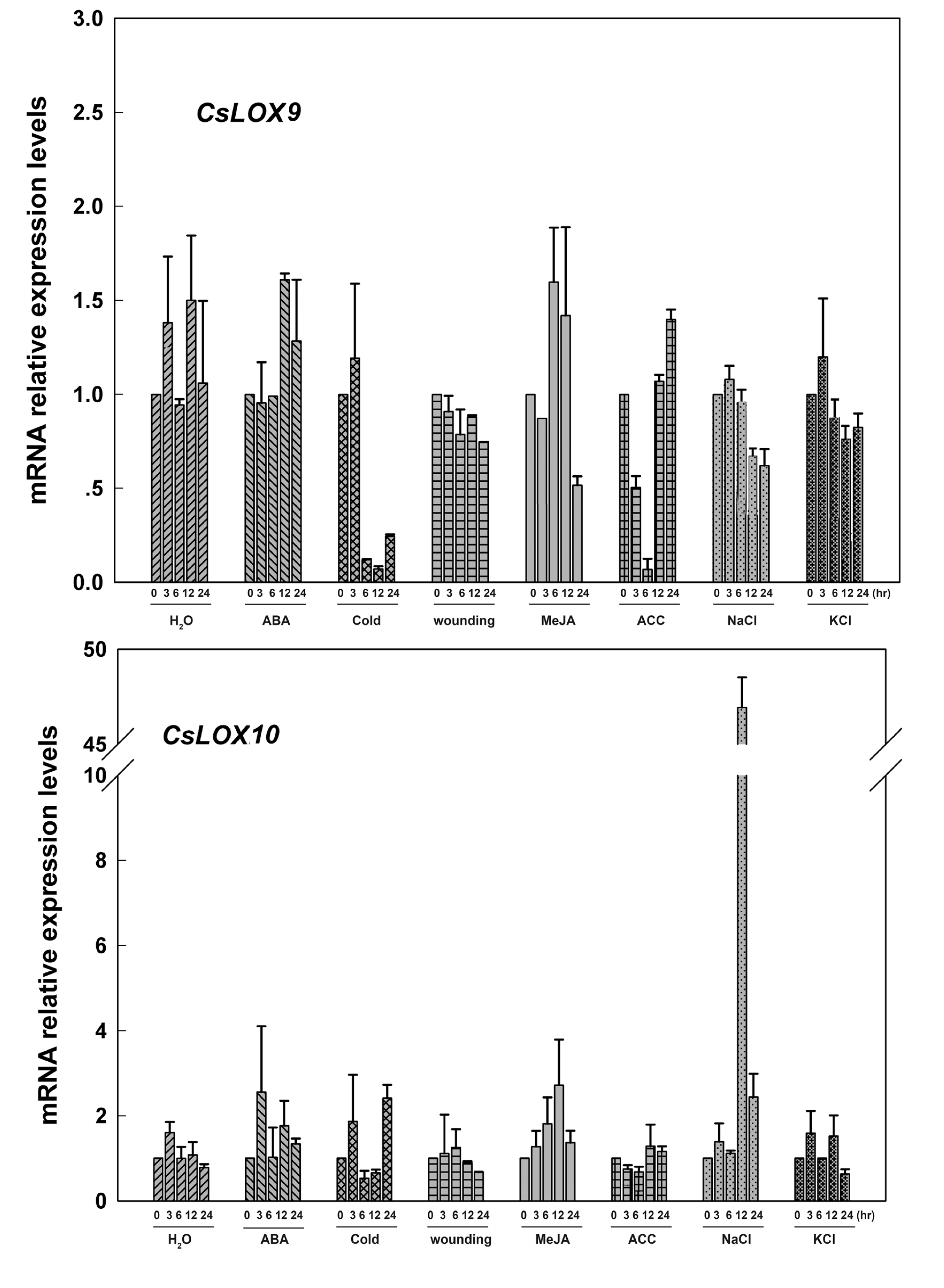
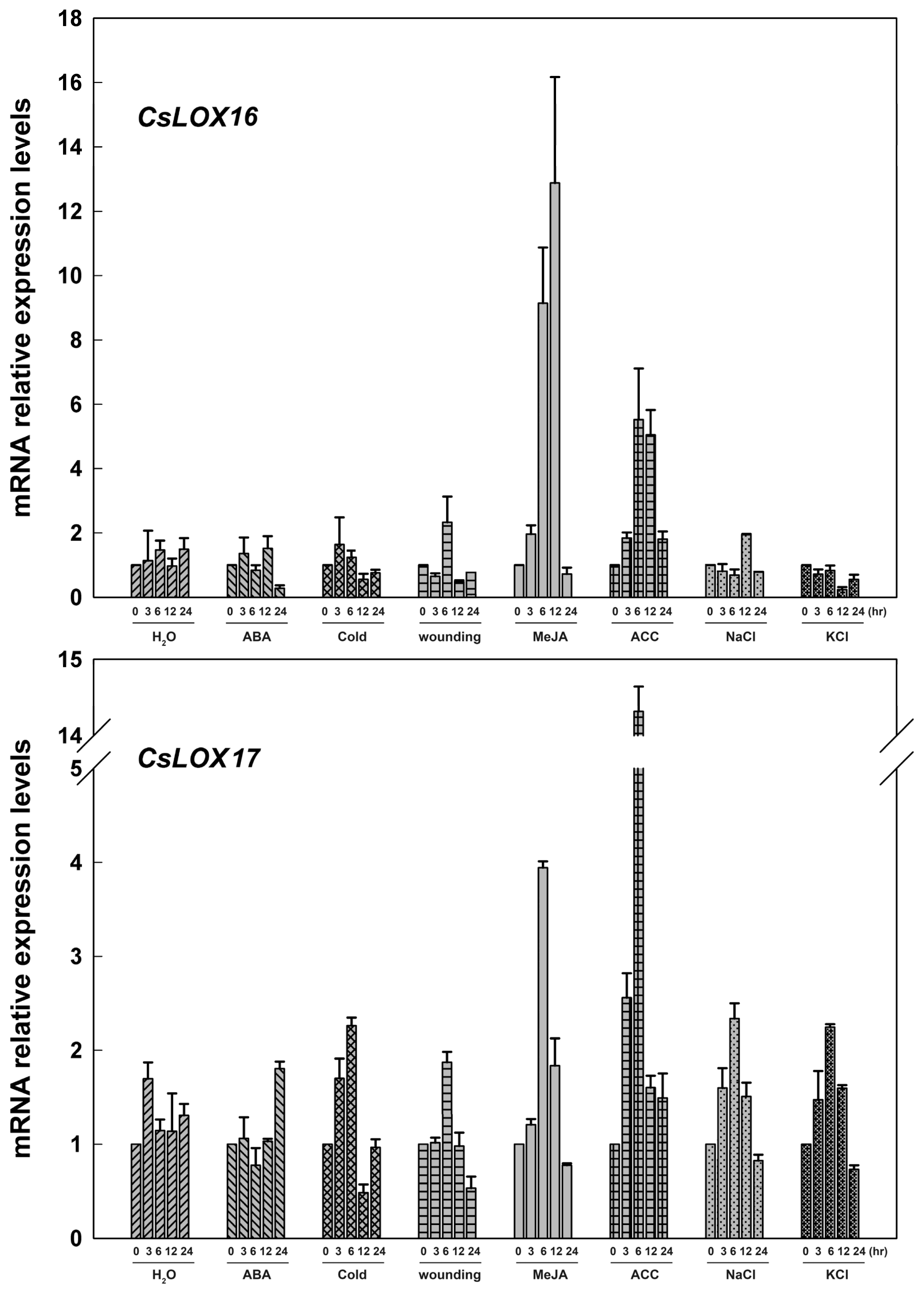
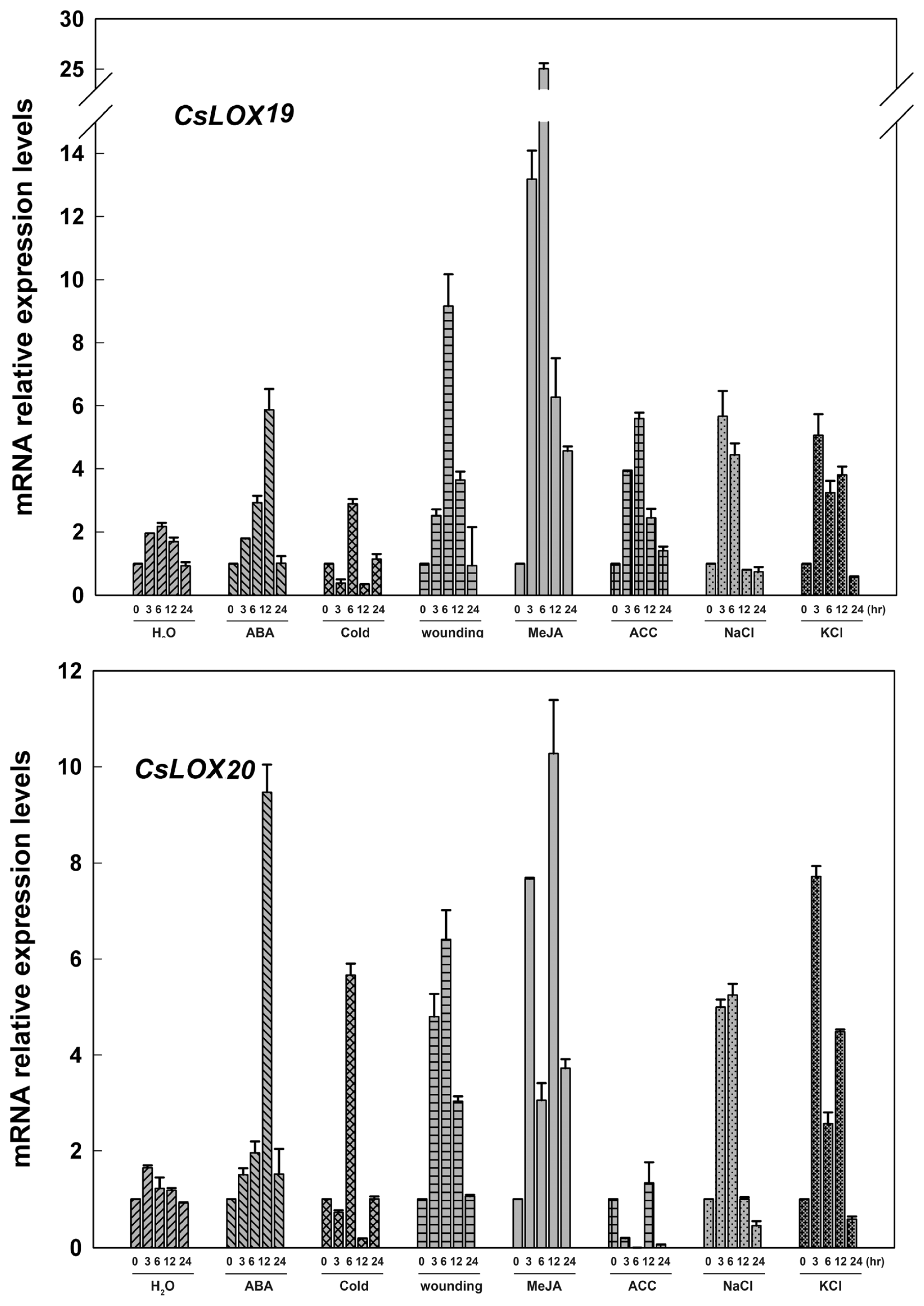
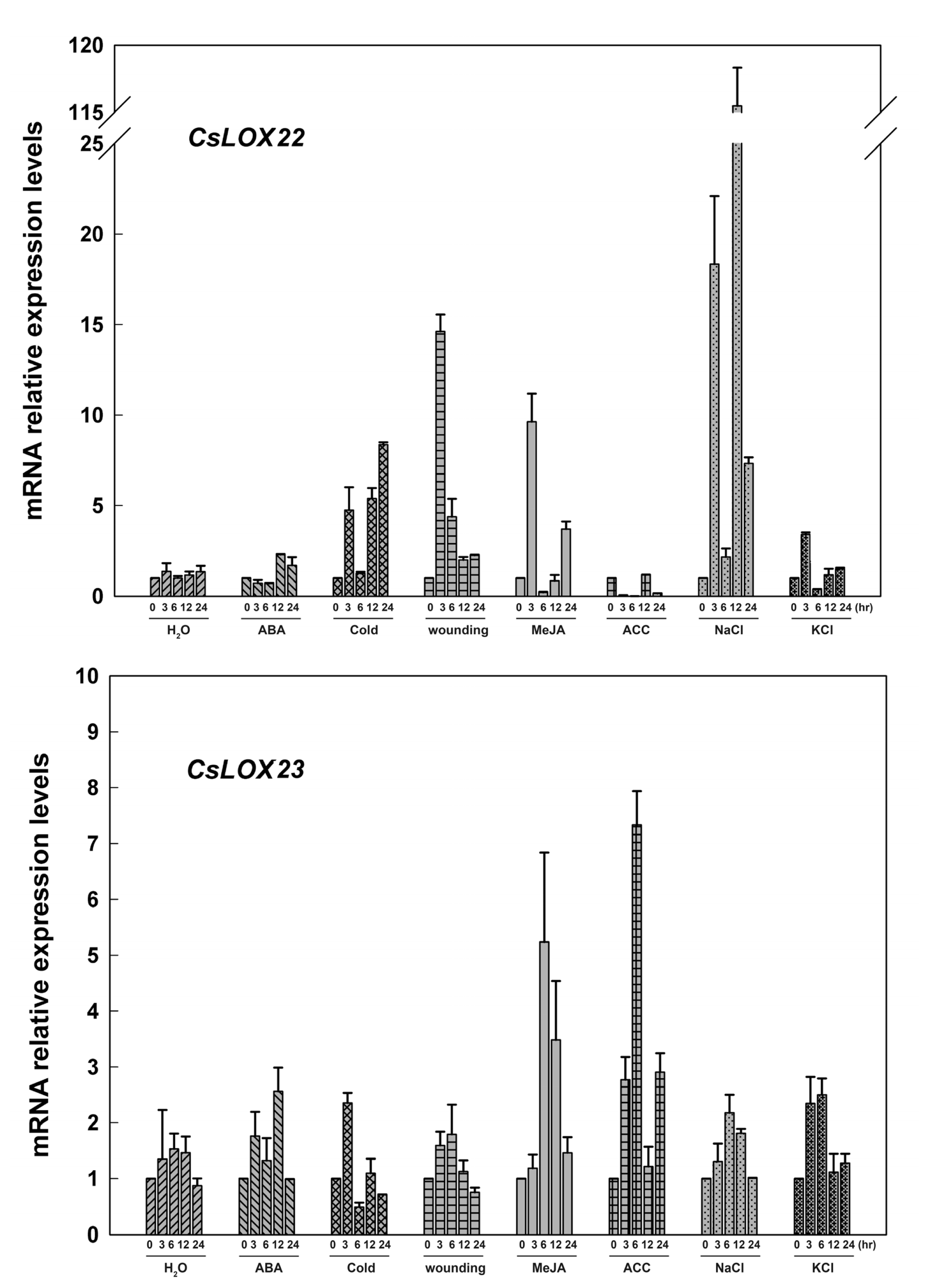
2.3. The Expression of Fruit CsLOX Genes in Response to Plant Hormones and Abiotic Stress Treatments
3. Discussion
3.1. Cucumber LOX in Fruit Development
3.2. Cucumber LOX Gene Expressions in Response to Plant-Growth Regulators and Abiotic Stresses
4. Methods
4.1. Plant Materials and Stress Treatments
4.2. RNA Isolation and Semi-Quantitative RT-PCR
4.3. Quantitative RT-PCR Analysis
5. Conclusions
Acknowledgements
Reference
- Brash, A.R. Lipoxygenases: occurrence, functions, catalysis, and acquisition of substrate. J. Biol. Chem 1999, 274, 23679–23682. [Google Scholar]
- Kolomiets, M.V.; Hannapel, D.J.; Chen, H.; Tymeson, M.; Gladon, R.J. Lipoxygenase is involved in the control of potato tuber development. Plant Cell 2001, 13, 613–626. [Google Scholar]
- Porta, H.; Rueda-Benítez, P.; Campos, F.; Colmenero-Flores, J.M.; Colorado, J.M.; Carmona, M.J.; Covarrubias, A.A.; Rocha-Sosa, M. Analysis of lipoxygenase mRNA accumulation in the common bean (Phaseolus vulgaris L.) during development and under stress conditions. Plant Cell Physiol 1999, 40, 850–858. [Google Scholar]
- Siedow, J.N. Plant lipoxygenase: structure and function. Annu. Rev. Plant Biol 1991, 42, 145–188. [Google Scholar]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol 2002, 53, 275–297. [Google Scholar]
- Porta, H.; Rocha-Sosa, M. Plant lipoxygenases. Physiological and molecular features. Plant Physiol 2002, 130, 15–21. [Google Scholar]
- Liavonchanka, A.; Feussner, I. Lipoxygenases: occurrence, functions and catalysis. J. Plant Physiol 2006, 163, 348–357. [Google Scholar]
- Casey, R.; West, S.I.; Hardy, D.; Robinson, D.S.; Zecai, W.; Hughes, R.K. New frontiers in food enzymology: recombinant lipoxygenases. Trends Food Sci. Tech 1999, 10, 297–302. [Google Scholar]
- Schwab, W.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant J 2008, 54, 712–732. [Google Scholar]
- Buescher, R.; Buescher, R. Production and Stability of (E, Z)-2,6-Nonadienal, the Major Flavor Volatile of Cucumbers. J. Food Sci 2001, 66, 357–361. [Google Scholar]
- Thompson, J.E. The Molecular Basis for Membrane Deterioration During Senescence. In Senescence and Aging in Plants; Nooden, L.D., Leopold, A.C., Eds.; Academic Press: San Diego, CA USA, 1988. [Google Scholar]
- Rogiers, S.Y.; Kumar, G.N.M.; Knowles, N.R. Maturation and Ripening of Fruit of Amelanchier alnifolia Nutt. are Accompanied by Increasing Oxidative Stress. Ann. Bot 1998, 81, 203–211. [Google Scholar]
- Brennan, T.; Frenkel, C. Involvement of hydrogen peroxide in the regulation of senescence in pear. Plant Physiol 1977, 59, 411–416. [Google Scholar]
- Brennan, T.; Rychter, A.; Frenkel, C. Activity of enzymes involved in the turnover of hydrogen peroxide during fruit senescence. Bot. Gaz 1979, 140, 384–388. [Google Scholar]
- Chen, K.S.; Xu, C.J.; Lou, J.; Zhang, S.L.; Ross, G. Lipoxygenase in relation to the ripening and softening of Actinidia fruit. Acta Phytophysiol. Sinica 1999, 25, 138–144. [Google Scholar]
- Zhang, Y.; Chen, K.; Zhang, S.; Ferguson, I. The role of salicylic acid in postharvest ripening of kiwifruit. Postharvest Biol. Tech 2003, 28, 67–74. [Google Scholar]
- Wu, M.; Chen, K.S.; Zhang, S.L. Involvement of lipoxygenase in the postharvest ripening of peach fruit. Acta Hortic. Sinica 1999, 26, 227–231. [Google Scholar]
- Baldwin, E.; Nisperos-Carriedo, M.; Moshonas, M. Quantitative analysis of flavor and other volatiles and for certain constituents of two tomato cultivars during ripening. J. Am. Soc. Hortic. Sci 1991, 116, 265–269. [Google Scholar]
- Chen, G.; Hackett, R.; Walker, D.; Taylor, A.; Lin, Z.; Grierson, D. Identification of a specific isoform of tomato lipoxygenase (TomloxC) involved in the generation of fatty acid-derived flavor compounds. Plant Physiol 2004, 136, 2641–2. [Google Scholar]
- Echeverría, G.; Graell, J.; López, M.; Lara, I. Volatile production, quality and aroma-related enzyme activities during maturation of Fuji’ apples. Postharvest Biol. Tech 2004, 31, 217–227. [Google Scholar]
- Perez, A.G.; Sanz, C.; Olías, R.; Olías, J.M. Lipoxygenase and hydroperoxide lyase activities in ripening strawberry fruits. J. Agr. Food Chem 1999, 47, 249–253. [Google Scholar]
- Bannenberg, G.; Martínez, M.; Hamberg, M.; Castresana, C. Diversity of the Enzymatic Activity in the Lipoxygenase Gene Family of Arabidopsis thaliana. Lipids 2009, 44, 85–95. [Google Scholar]
- Melan, M.A.; Dong, X.; Endara, M.E.; Davis, K.R.; Ausubel, F.M.; Peterman, T.K. An Arabidopsis thaliana lipoxygenase gene can be induced by pathogens, abscisic acid, and methyl jasmonate. Plant Physiol 1993, 101, 441–450. [Google Scholar]
- Bell, E.; Creelman, R.A.; Mullet, J.E. A chloroplast lipoxygenase is required for wound-induced jasmonic acid accumulation in Arabidopsis. Proc. Natl. Acad. Sci 1995, 92, 8675–8679. [Google Scholar]
- Caldelari, D.; Wang, G.; Farmer, E.E.; Dong, X. Arabidopsis lox3 lox4 double mutants are male sterile and defective in global proliferative arrest. Plant Mol. Biol 2011, 75, 25–33. [Google Scholar]
- Ferrie, B.J.; Beaudoin, N.; Burkhart, W.; Bowsher, C.G.; Rothstein, S.J. The cloning of two tomato lipoxygenase genes and their differential expression during fruit ripening. Plant Physiol 1994, 106, 109–118. [Google Scholar]
- Griffiths, A.; Barry, C.; Alpuche-Solis, A.G.; Grierson, D. Ethylene and developmental signals regulate expression of lipoxygenase genes during tomato fruit ripening. J. Exp. Bot 1999, 50, 793–798. [Google Scholar]
- Heitz, T.; Bergey, D.R.; Ryan, C.A. A gene encoding a chloroplast-targeted lipoxygenase in tomato leaves is transiently induced by wounding, systemin, and methyl jasmonate. Plant Physiol. 1997, 114, 1085–1093. [Google Scholar]
- Mariutto, M.; Duby, F.; Adam, A.; Bureau, C.; Fauconnier, M.L.; Ongena, M.; Thonart, P.; Dommes, J. The elicitation of a systemic resistance by Pseudomonas putida BTP1 in tomato involves the stimulation of two lipoxygenase isoforms. BMC Plant Biol 2011, 11. [Google Scholar] [CrossRef]
- León, J.; Royo, J.; Vancanneyt, G.; Sanz, C.; Silkowski, H.; Griffiths, G.; Sánchez-Serrano, J.J. Lipoxygenase H1 gene silencing reveals a specific role in supplying fatty acid hydroperoxides for aliphatic aldehyde production. J. Biol. Chem 2002, 277, 416–423. [Google Scholar]
- Royo, J.; Vancanneyt, G.; Pérez, A.G.; Sanz, C.; Störmann, K.; Rosahl, S.; Sánchez-Serrano, J.J. Characterization of three potato lipoxygenases with distinct enzymatic activities and different organ-specific and wound-regulated expression patterns. J. Biol. Chem 1996, 271, 21012–21019. [Google Scholar]
- Podolyan, A.; White, J.; Jordan, B.; Winefield, C. Identification of the lipoxygenase gene family from Vitis vinifera and biochemical characterisation of two 13-lipoxygenases expressed in grape berries of Sauvignon Blanc. Funct. Plant Biol 2010, 37, 767–784. [Google Scholar]
- Zhang, B.; Chen, K.; Bowen, J.; Allan, A.; Espley, R.; Karunairetnam, S.; Ferguson, I. Differential expression within the LOX gene family in ripening kiwifruit. J. Exp. Bot 2006, 57, 3825–3836. [Google Scholar]
- Höhne, M.; Nellen, A.; Schwennesen, K.; Kindl, H. Lipid body lipoxygenase characterized by protein fragmentation, cDNA sequence and very early expression of the enzyme during germination of cucumber seeds. Eur. J. Biochem 1996, 241, 6–11. [Google Scholar]
- Matsui, K.; Nishioka, M.; Ikeyoshi, M.; Matsumura, Y.; Mori, T.; Kajiwara, T. Cucumber root lipoxygenase can act on acyl groups in phosphatidylcholine. Biochim. Biophys. Acta Lipids Lipid Metabol 1998, 1390, 8–20. [Google Scholar]
- Matsui, K.; Minami, A.; Hornung, E.; Shibata, H.; Kishimoto, K.; Ahnert, V.; Kindl, H.; Kajiwara, T.; Feussner, I. Biosynthesis of fatty acid derived aldehydes is induced upon mechanical wounding and its products show fungicidal activities in cucumber. Phytochemistry 2006, 67, 649–657. [Google Scholar]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J; Wang, X.; Xie, B.; Ni, P. The genome of the cucumber, Cucumis sativus L. Nature Genet. 2009, 41, 1275–1281. [Google Scholar]
- Liu, S.; Liu, X.; Jiang, L. Genome-wide identification, phylogeny and expression analysis of the lipoxygenase gene family in cucumber. Genet. Mol. Res. GMR 2011, 10, 2613–2636. [Google Scholar]
- Pena-Cortes, H.; Willmitzer, L.; Sánchez-Serrano, J.J. Abscisic acid mediates wound induction but not developmental-specific expression of the proteinase inhibitor II gene family. Plant Cell 1991, 3, 963–972. [Google Scholar]
- Penninckx, I.A.M.A.; Thomma, B.P.H.J.; Buchala, A.; Métraux, J.P.; Broekaert, W.F. Concomitant activation of jasmonate and ethylene response pathways is required for induction of a plant defensin gene in Arabidopsis. Plant Cell 1998, 10, 2103–2114. [Google Scholar]
- Berrocal-Lobo, M.; Molina, A.; Solano, R. Constitutive expression of ETHYLENE-RESPONSEFACTOR1 in Arabidopsis confers resistance to several necrotrophic fungi. Plant J 2002, 29, 23–32. [Google Scholar]
- Lorenzo, O.; Piqueras, R.; Sánchez-Serrano, J.J.; Solano, R. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell 2003, 15, 165–178. [Google Scholar]
- Lester, G.E.; Jifon, J.L.; Rogers, G. Supplemental foliar potassium applications during muskmelon fruit development can improve fruit quality, ascorbic acid, and beta-carotene contents. J. Am. Soc. Hortic. Sci 2005, 130, 649–653. [Google Scholar]
- Hartz, T.; Johnstone, P.; Francis, D.; Miyao, E. Processing tomato yield and fruit quality improved with potassium fertigation. HortScience 2005, 40, 1862–1867. [Google Scholar]
- Vellosillo, T.; Martínez, M.; López, M.A.; Vicente, J.; Cascón, T.; Dolan, L.; Hamberg, M.; Castresana, C. Oxylipins produced by the 9-lipoxygenase pathway in Arabidopsis regulate lateral root development and defense responses through a specific signaling cascade. Plant Cell 2007, 19, 831–846. [Google Scholar]
- Ben-Hayyim, G.; Gueta-Dahan, Y.; Avsian-Kretchmer, O.; Weichert, H.; Feussner, I. Preferential induction of a 9-lipoxygenase by salt in salt-tolerant cells of Citrus sinensis L. Osbeck. Planta 2001, 212, 367–375. [Google Scholar]
- Liu, C.; He, Q.; Ai, X. Changes of some aromatic compounds and relative factors during fruit development of cucumber. China Vegetables 2006, 1, 9–12. [Google Scholar]
- Creelman, R.A.; Mullet, J.E. Biosynthesis and action of jasmonates in plants. Ann. Rev. Plant Biol 1997, 48, 355–381. [Google Scholar]
- Nemchenko, A.; Kunze, S.; Feussner, I.; Kolomiets, M. Duplicate maize 13-lipoxygenase genes are differentially regulated by circadian rhythm, cold stress, wounding, pathogen infection, and hormonal treatments. J. Exp. Bot 2006, 57, 3767–3779. [Google Scholar]
- Bate, N.J.; Rothstein, S.J. C6-volatiles derived from the lipoxygenase pathway induce a subset of defense-related genes. Plant J 1998, 16, 561–569. [Google Scholar]
- Parchmann, S.; Gundlach, H.; Mueller, M.J. Induction of 12-oxo-phytodienoic acid in wounded plants and elicited plant cell cultures. Plant Physiol 1997, 115, 1057–1064. [Google Scholar]
- Sheng, J.; Luo, Y.; Wainwright, H. Studies on lipoxygenase and the formation of ethylene in tomato. J. Hortic. Sci. Biotechnol 2000, 75, 69–71. [Google Scholar]
- Zhang, Y.; Chen, K.S.; Chen, Q.J.; Zhang, S.L.; Ren, Y.P. Effects of Acetylsalicylic Acid (ASA) and Ethylene Treatments on Ripening and Softening of Postharvest Kiwifruit. Acta Bot. Sin 2003, 45, 1447–1452. [Google Scholar]
- Tuteja, N. Abscisic acid and abiotic stress signaling. Plant Signal. Behav 2007, 2, 135–138. [Google Scholar]
- Maccarrone, M.; Veldink, G.A.; Vliegenthart, J.F.G. Modulation of soybean lipoxygenase expression and membrane oxidation by water deficit. FEBS Lett 1995, 371, 223–226. [Google Scholar]
- Jia, W.; Wang, Y.; Zhang, S.; Zhang, J. Salt-stress-induced ABA accumulation is more sensitively triggered in roots than in shoots. J. Exp. Bot 2002, 53, 2201–2206. [Google Scholar]









© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yang, X.-Y.; Jiang, W.-J.; Yu, H.-J. The Expression Profiling of the Lipoxygenase (LOX) Family Genes During Fruit Development, Abiotic Stress and Hormonal Treatments in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2012, 13, 2481-2500. https://doi.org/10.3390/ijms13022481
Yang X-Y, Jiang W-J, Yu H-J. The Expression Profiling of the Lipoxygenase (LOX) Family Genes During Fruit Development, Abiotic Stress and Hormonal Treatments in Cucumber (Cucumis sativus L.). International Journal of Molecular Sciences. 2012; 13(2):2481-2500. https://doi.org/10.3390/ijms13022481
Chicago/Turabian StyleYang, Xue-Yong, Wei-Jie Jiang, and Hong-Jun Yu. 2012. "The Expression Profiling of the Lipoxygenase (LOX) Family Genes During Fruit Development, Abiotic Stress and Hormonal Treatments in Cucumber (Cucumis sativus L.)" International Journal of Molecular Sciences 13, no. 2: 2481-2500. https://doi.org/10.3390/ijms13022481
APA StyleYang, X. -Y., Jiang, W. -J., & Yu, H. -J. (2012). The Expression Profiling of the Lipoxygenase (LOX) Family Genes During Fruit Development, Abiotic Stress and Hormonal Treatments in Cucumber (Cucumis sativus L.). International Journal of Molecular Sciences, 13(2), 2481-2500. https://doi.org/10.3390/ijms13022481