1. Introduction
Abiotic stress such as drought and salinity impose serious limitations to plant growth, development and crop productivity [
1]. In order to survive adverse conditions, plants have developed multiple strategies, including physiological, biochemical and molecular mechanisms to counteract damages caused by environmental conditions. At the molecular level, the expression of stress responsive genes has been classified into: the early- and the delayed-response genes. The first group of genes is induced very quickly and often transiently after stress perception, such as kinases and transcription factors. The delayed-response genes are activated more slowly and their expression is often sustained during the stress conditions, encoding stress proteins such as heat shock proteins (HSPs), late embryogenesis abundant (LEAs), aquaporins, enzymes for osmolyte metabolism, antioxidant proteins among others [
2–
5].
HSPs or molecular chaperones are produced in all organisms in response to elevated temperatures and other stress conditions; its induction is correlated with the acquisition of thermotolerance [
6–
8]. HSPs are involved in a variety of cellular processes including protein folding, assembly of oligomeric proteins, transport of proteins across membranes, stabilization of polypeptide strands and membranes, and prevention of protein inactivation [
8–
10]. In addition, HSPs participate in regulating the activation and function of target proteins, although themselves are not components of the target proteins [
10]. These proteins are classified in plants into five classes according to their molecular weight: HSP100, HSP90, HSP70, HSP60, and small HSP (sHSP) [
9,
11,
12].
sHSPs constitute a family of structurally diverse and abundant HSPs synthesized by plants, usually undetectable under normal conditions but they can be induced by stress conditions and developmental stimuli [
13]. The monomeric molecular mass of sHSPs ranges from 12–42 kDa, but in their native state the majority of sHSPs form homo- and hetero-oligomers of 12 to 32 subunits [
14], although there are also dimeric and tetrameric sHSPs [
12,
15]. sHSPs are defined by a core α-crystalline domain of about 100 amino acids that is conserved in all plant sHSPs. This domain is responsible for dimer formation (sHSP-sHSP), which is enriched in β-strands organized in a β-sheet sandwich, the basic structural unit of many sHSPs [
16]. Also, this α-crystalline domain is flanked by an
N-terminal region highly variable in size and sequence that has been suggested as a candidate site for substrate binding. On the other hand, the
C-terminal region is short and flexible, and it has been proposed as the site of contact that stabilizes the oligomeric structure [
12,
14,
17]. In the
Arabidopsis thaliana genome, at least 19 open reading frames (ORFs) encoding sHSPs have been identified [
18]. These proteins are grouped into six classes, based on their intracellular localization, three sHSP classes (CI, CII and CIII) are localized in the cytosol or nucleus, and other classes in plastids (class P), in the endoplasmic reticulum (class ER) and in the mitochondria (class M). In addition, a set of sHSP-like proteins have been described and grouped as: class CI related and class P related proteins [
18].
Cactus pear (
Opuntia spp.) is a perennial arborescent cactus widely distributed in the arid and semiarid regions of Mexico, where it plays a strategic role in subsistence agriculture. Cactus pear is edible as a fruit and vegetable, and is also used as forage crop and for medicinal purpose.
Opuntia plants exhibit Crassulacean Acid Metabolism (CAM) photosynthesis, in which CO
2 exchange occurs at night, when the water vapor pressure difference between the air and the transpiring surfaces is lower, resulting in high water use efficiency [
19]. These plants present several extraordinary morphological adaptations to reduce water loss, for example: the globular or racquet-like structures known as cladodes, which store large amounts of water, the replacement of leaves by thorns, thick cuticle cladode, low stomatal frequency, and sunken stomata [
20,
21]. Because of these adaptations and its tolerance to drought, this plant can cope with a variety of environmental conditions [
22,
23].
Recently, our group reported a cDNA library of
Opuntia streptacantha cladodes containing a collection of ESTs accumulated under abiotic stress [
24]. Among the genes involved in stress response obtained in this library, a small heat shock protein (sHSP) was identified. In the present study, we have approached the functional characterization of the
OpsHSP18 gene, encoding an 18.34 kDa sHSP, by analyzing a series of
Arabidopsis thaliana over-expressing lines
35S::OpsHSP18. Results reveal that
35S::OpsHSP18 over-expressing lines are tolerant to salt and osmotic stress, suggesting that OpsHSP18 protein has a role in plant abiotic stress tolerance.
3. Discussion
Plants are sessile organisms, which are daily exposed to unfavorable conditions such as heat, cold, salt, drought, among others, which limit their growth and development. Processes such as seed germination, seedling growth and vigor, vegetative growth, flowering and fruit set are adversely affected by high salt concentration, ultimately causing diminished economic yield and also reduced quality of production [
29]. Thus, plants have developed mechanisms such as the synthesis of heat shock proteins to adapt to those complex environments.
Recently, our research group generated a cDNA library from
Opuntia streptacantha plants under abiotic stress conditions [
24]. Among 329 unigenes of this cDNA library, 9% (29 unigenes) were involved in stress responsiveness, such as the heat shock proteins: HSP90, HSP70, HSP40, and sHSP. sHSPs are associated to the nuclei, cytoskeleton, and membranes, and they bind partially to denatured proteins, avoiding irreversible protein aggregation during stress conditions [
16].
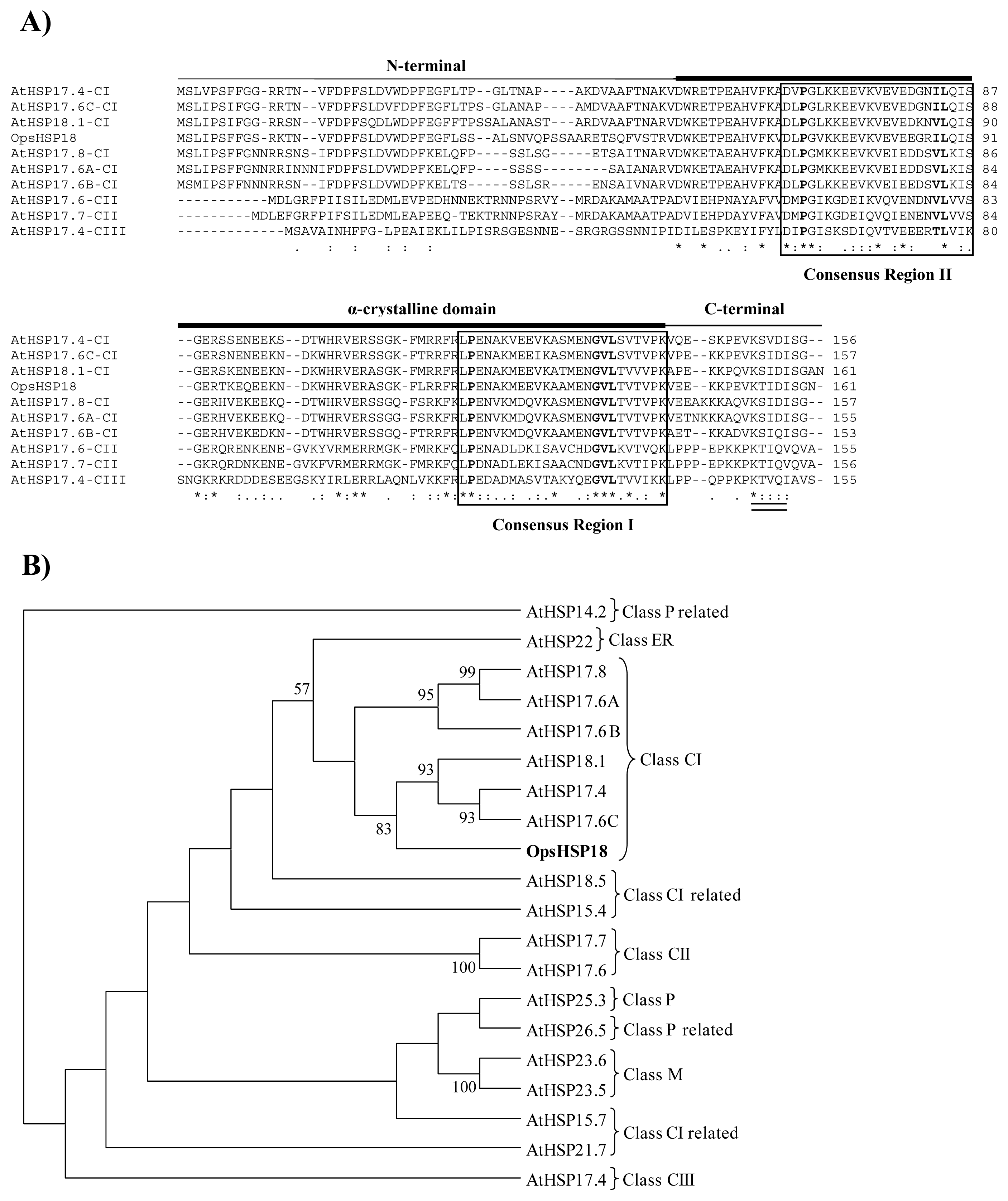
In this study, the small heat shock protein (OpsHSP18) was selected for functional characterization. Cactus pear OpsHSP18 protein displays characteristic features of the sHSP family, such as the α-crystalline domain, which has been reported to be involved in substrate binding and, in the interaction between the subunits of oligomeric complexes formed by sHSPs [
30]. This domain is flanked by an
N-terminal region of variable size and sequence, which has been proposed to have a role in unfolded substrate binding and specificity, and finally the
C-terminal region which has been suggested to confer stability to the oligomeric structure [
14].
In the
Arabidopsis thaliana genome, 19 genes encoding sHSP proteins have been described and classified into six groups, the first three groups of sHSP belong to proteins that are located in the cytoplasm and nucleus (classes CI, CII, and CIII), and those located in mitochondria, plastids and endoplasmic reticulum (classes M, P, and ER, respectively) forming the remaining groups [
18]. Thus, based on the alignments and phylogenetic analyzes, the
O. streptacantha OpsHSP18 protein was grouped within class CI sHSP. Regarding Arabidopsis, it was shown that the AtHSP17.4, AtHSP18.1, and AtHSP17.6C proteins presented the highest percentage of identity with the cactus pear OpsHSP18 protein. Particularly, this conservation was found in the α-crystalline domain and the
C-terminal region, while the
N-terminal region of these sHSPs was the most variable.
Several studies have demonstrated that sHSPs play certain roles in plant development such as embryogenesis, seed maturation, seed imbibition and germination, pollen development and fruit maturation under normal and stress conditions [
12,
26,
31–
34].
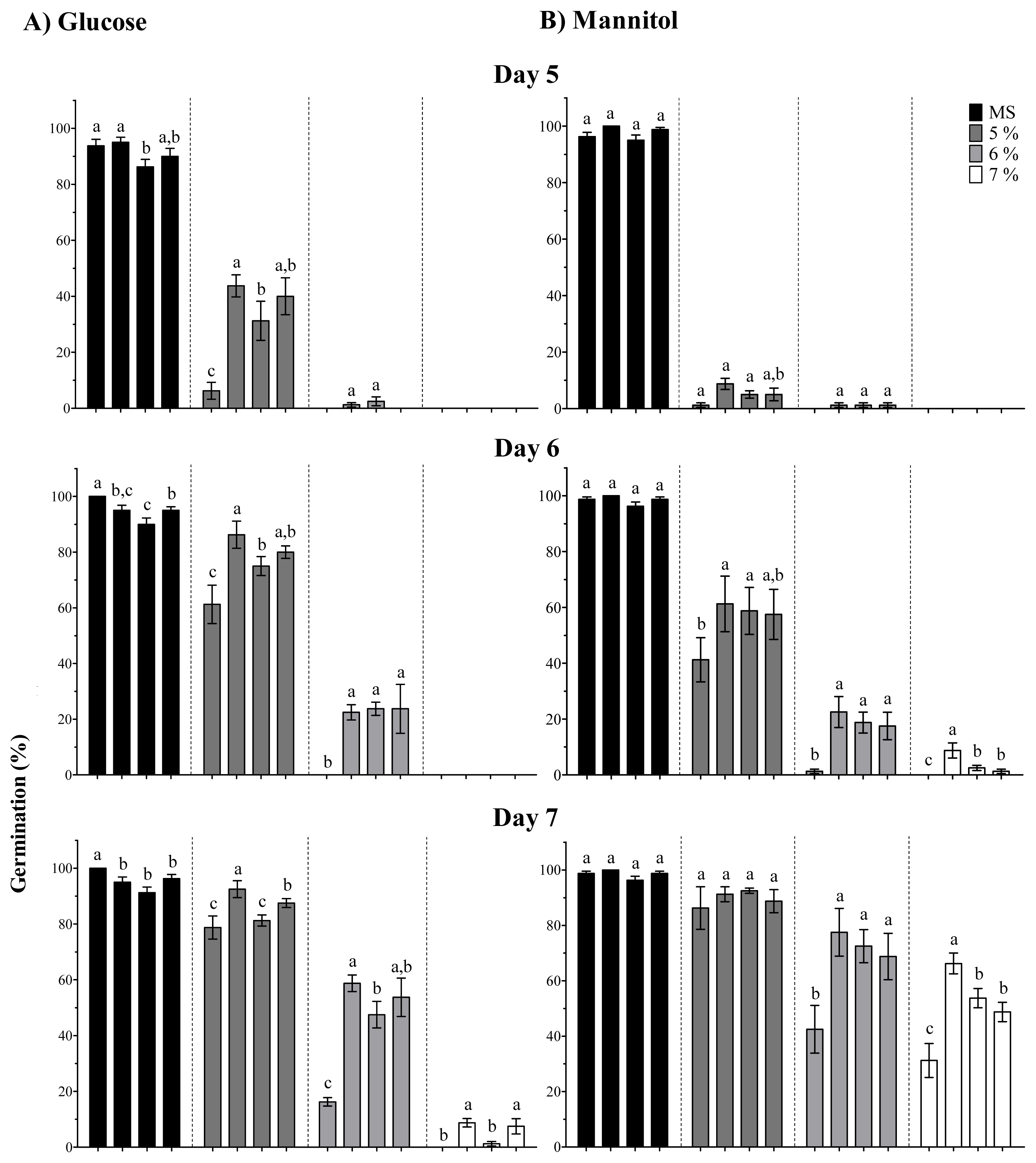
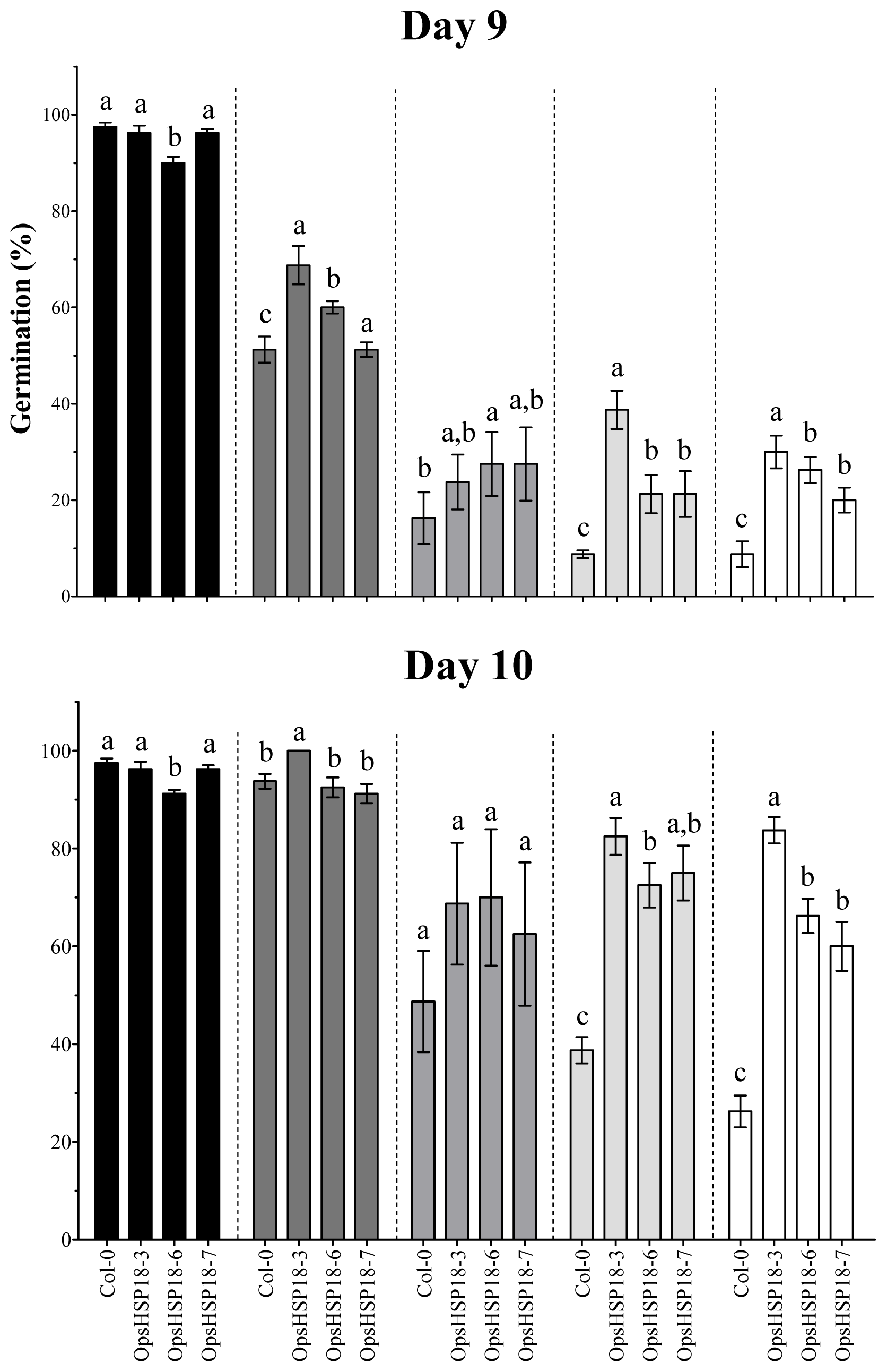
In order to gain insight into the function of the OpsHSP18 gene under abiotic stress, A. thaliana over-expressing lines were generated. When seed germination experiments were conducted under salt and osmotic stress, all three over-expressing lines exhibited higher germination percentages than the WT. Furthermore, over-expression of the cactus pear OpsHSP18 gene shows a positive effect on seedling development under stress, in which, Arabidopsis seedlings displayed a greater number of green cotyledons under salt and glucose conditions. Thus, our data suggest that over-expression of OpsHSP18 gene in A. thaliana might influence seed vigor under stressful germination conditions.
In general, plant reproductive organs are more sensitive to stress conditions than vegetative organs, so that class CI sHSP can facilitate either acquisition of desiccation tolerance, or they function in the rehydration mechanism of embryos during seed germination [
33,
35]. Wehmeyer in 1996 [
31] found that AtHSP17.4 and AtHSP17.6C proteins that belong to the class CI sHSP of Arabidopsis are expressed in seed maturation. In addition, the expression of
AtHSP17.4,
AtHSP17.6C and
AtHSP18.1 appear to be induced under heat stress conditions at temperatures of 35 or 40 °C for 2 h [
36], and at 37 °C for 90 min [
37]. AtHSP17.6 protein encodes a class CII sHSP from
A. thaliana, it was detected in mid-maturation, being the transcript more abundant in dry seeds, and a rapid decline was reported 6 days after germination [
38]. On the other hand, high levels of
AtHSP17.6A transcripts accumulated in siliques along with acquisition of desiccation tolerance by seeds.
AtHSP17.6 (class CII) transcripts were less abundant than
AtHSP17.6A transcripts in seeds, but a strong and transient induction of
AtHSP17.6 transcript (CII) was observed after imbibition [
38]. This suggests a putative role of AtHSP17.6 (CII) in protecting proteins during rehydration, and a divergence in function between the two cytosolic AtHSP17.6 proteins.
Recently, the characterization of
NnHSP17.5 gene from
Nelumbo nucifera, which encodes a cytoplasmic class CII sHSP was reported [
34]. It was found that during seed germination the expression levels of
NnHSP17.5 declined rapidly within 3 d after imbibition, suggesting a role in early stages of seed germination. This gene was also found to be strongly regulated under heat and oxidative stress [
34].
On the other hand, we found that the over-expression of
OpsHSP18 gene in Arabidopsis increases significantly the germination rate under ABA treatments compared with WT. It has been suggested that ABI3 transcription factor may be necessary to induce several sHSP during seed development [
31,
32]. Our data show an ABA insensitive phenotype, since the over-expressing lines are less affected in the germination process under ABA treatments than the WT. This observation opens new questions about the role of sHSP and ABA signaling.
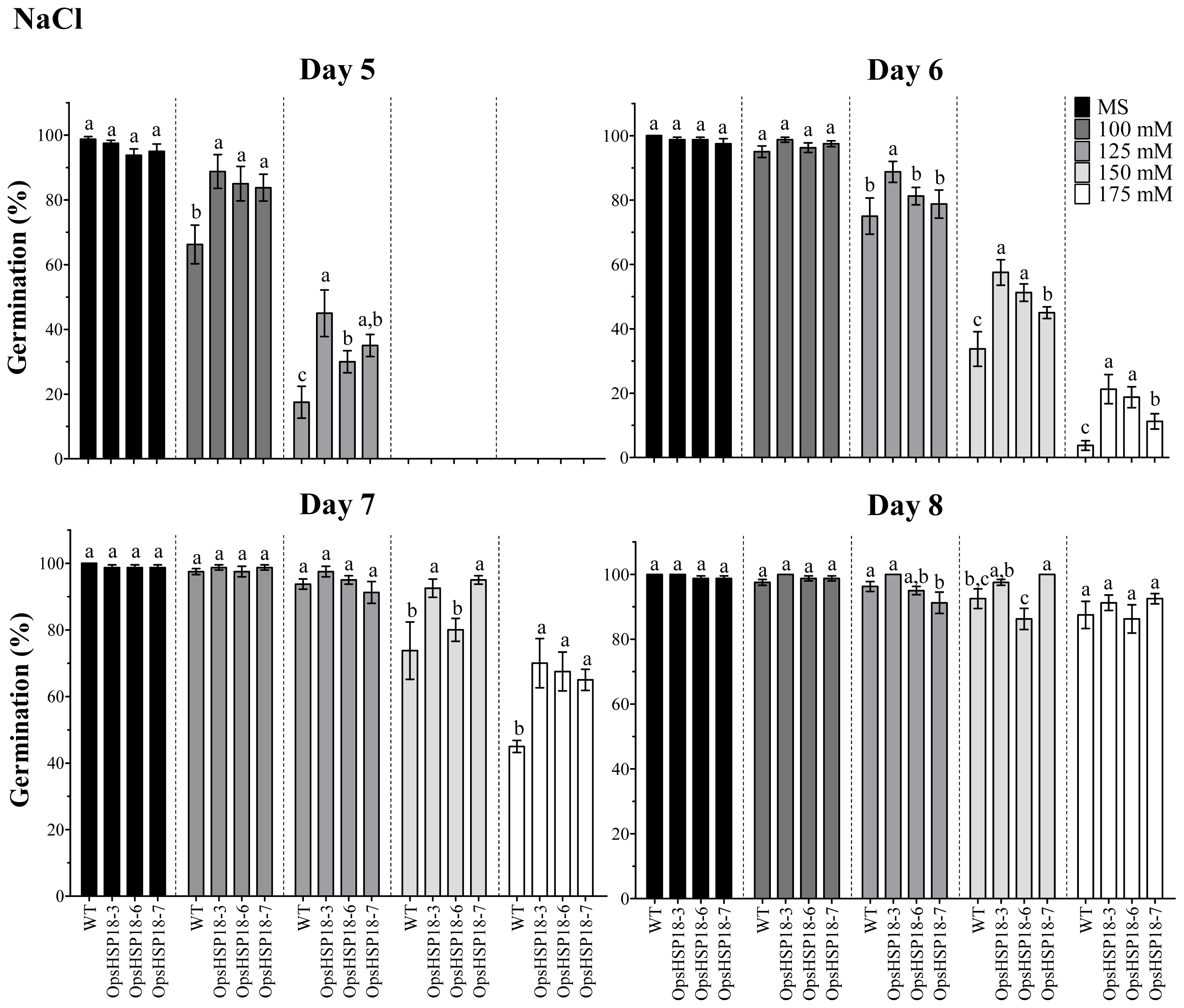
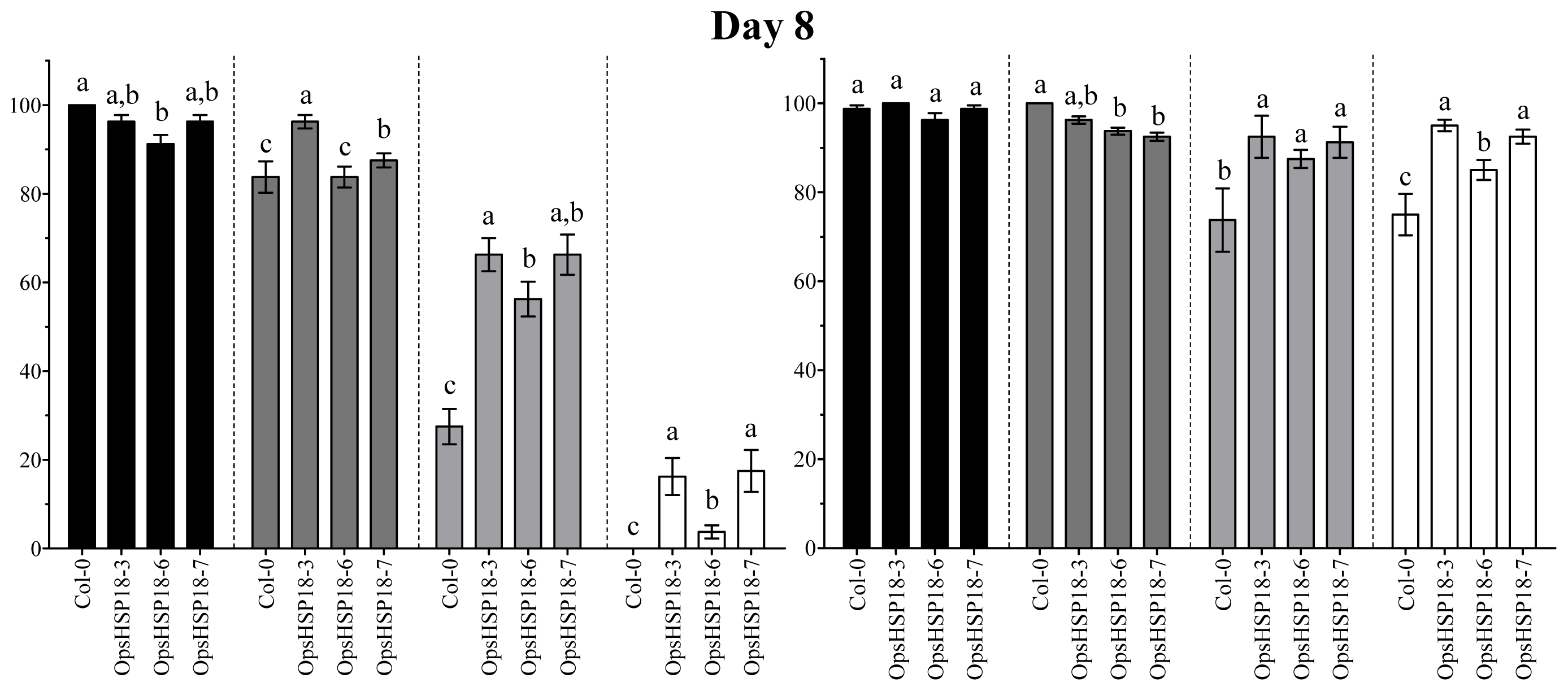
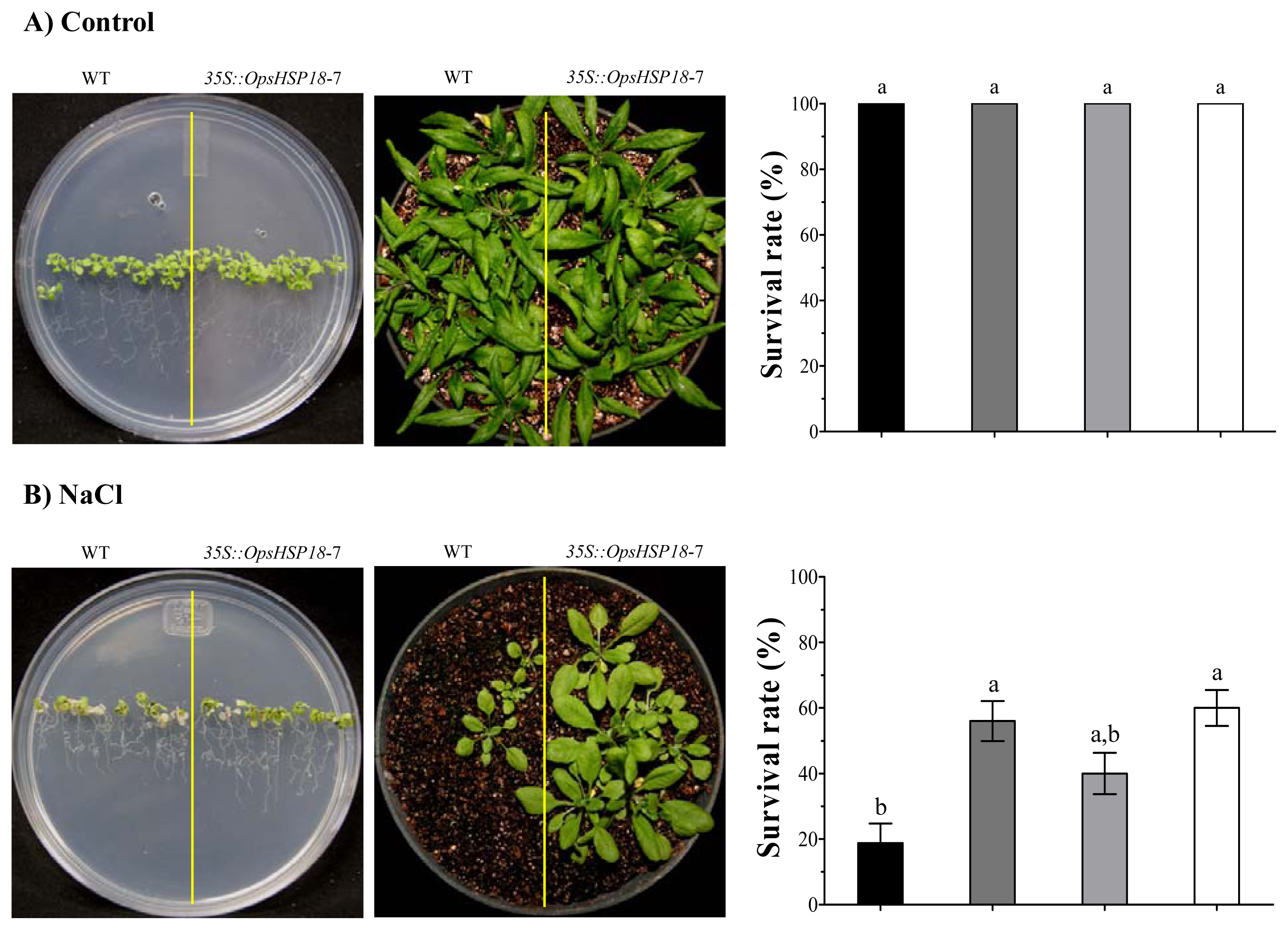
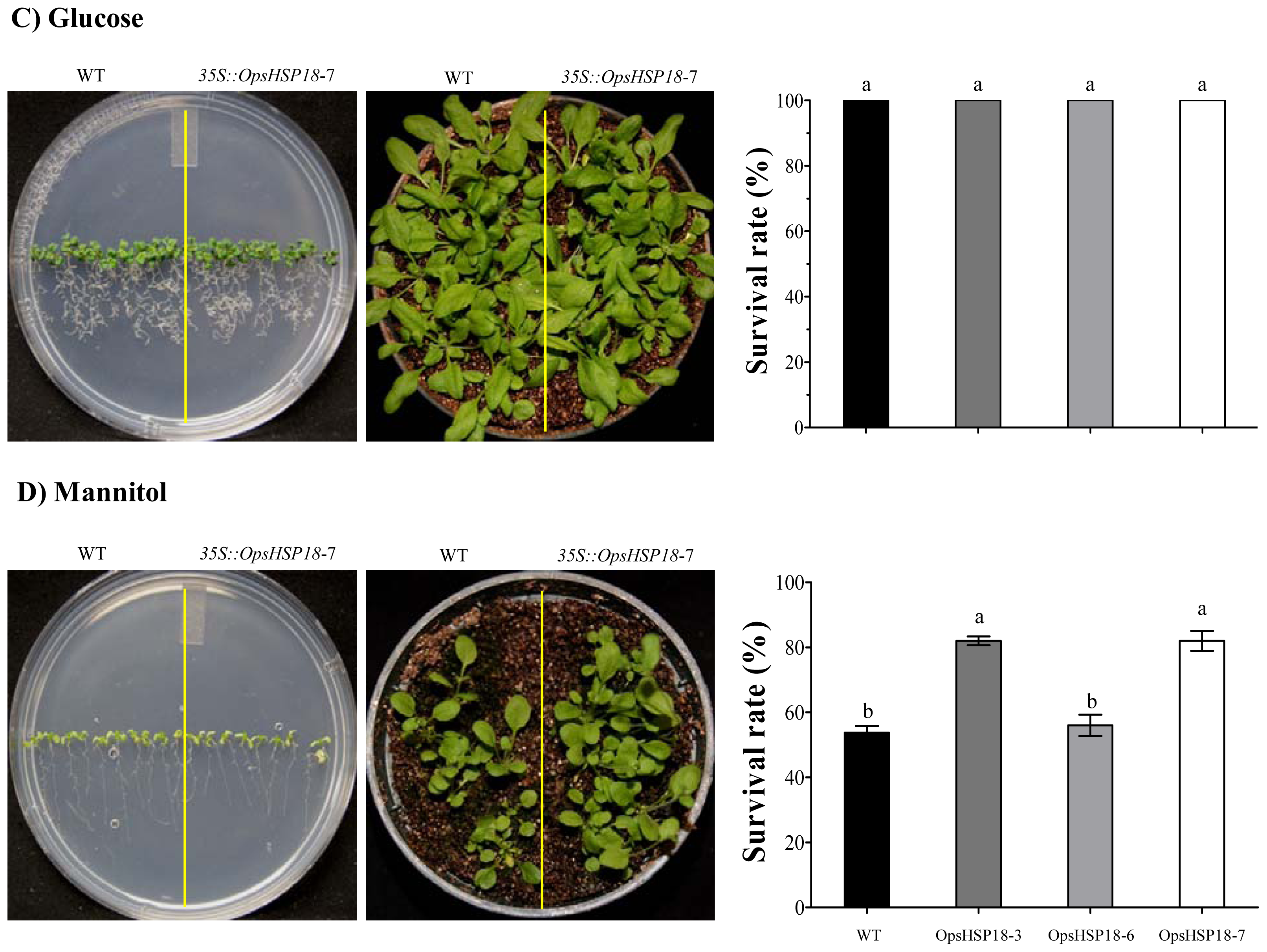
In addition, the over-expression of OpsHSP18 gene confers tolerance to salt and osmotic (mannitol) stress, increasing the survival rate of Arabidopsis transgenic lines after stress recovery. Differences in expression levels of the over-expressing lines were observed, in which a correlation between higher expression levels of lines 35S::OpsHSP18-3 and -7, and their tolerance to abiotic stress was obtained.
Different studies have shown that over-expression of
sHSPs confer tolerance to stress conditions. In transgenic rice, the over-expression of the
sHSP17.7 gene confers tolerance to heat and UV-B [
39], and drought stress [
40].
NnHSP17.5 over-expression in Arabidopsis transgenic plants showed an increase in the thermotolerance compared with WT plants at 40 °C for 60 or 75 min. The authors suggested that
NnHSP17.5 might play a protective role of proteins under stress conditions, such as superoxide dismutase (SOD), since SOD enzyme activity was found to be higher in transgenic lines than in WT plants [
34]. Furthermore, over-expression of
NnHSP17.5 in Arabidopsis transgenic lines confers tolerance to heat, salinity, drought, and osmotic stress [
34].
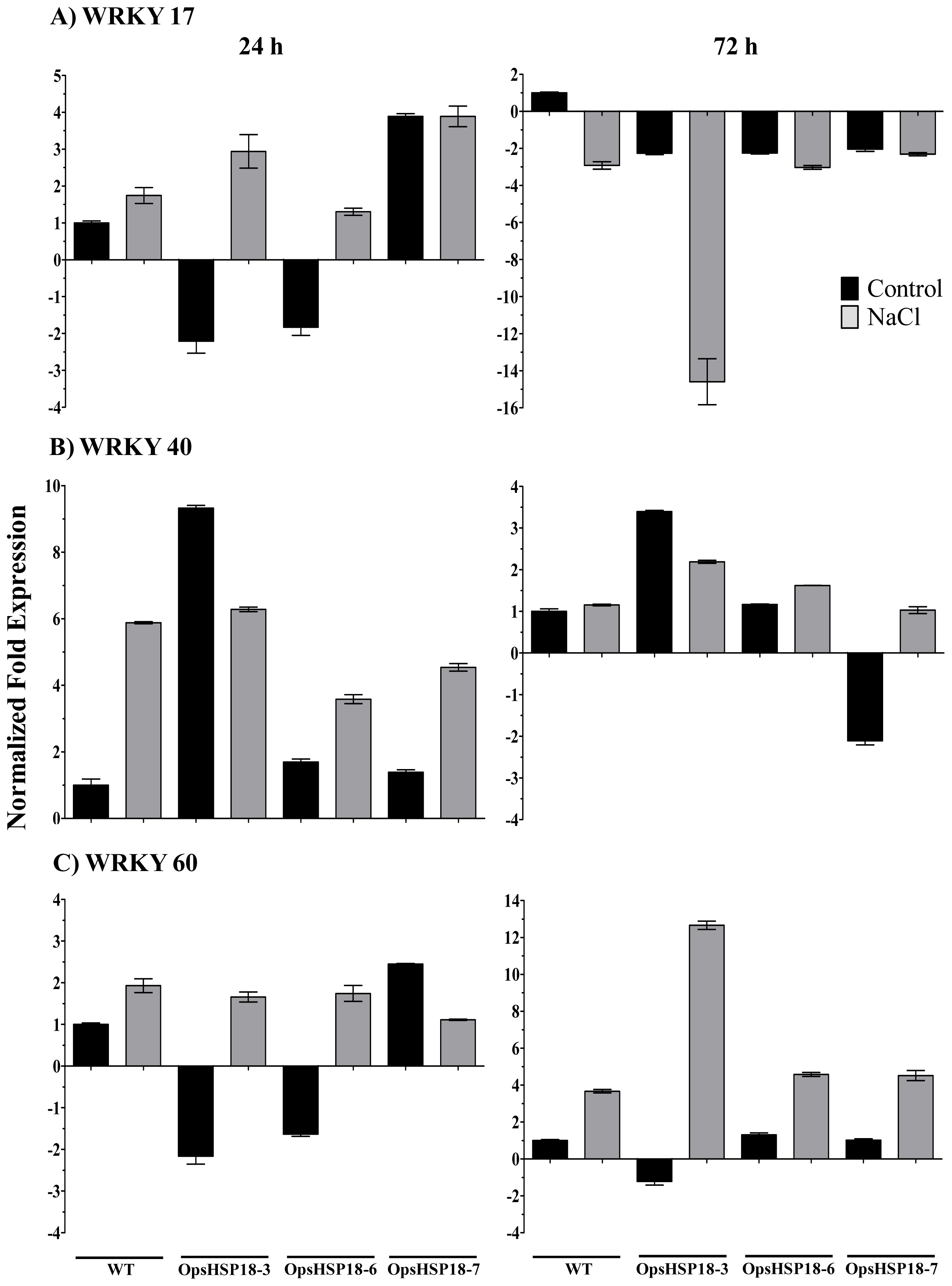
Finally, transcription factors play an important role in abiotic stress tolerance. In the last few years, changes in expression levels of some
WRKY transcription factors (
WRKY-17, -18, -25, -33, -40, and -60) have been described under salt stress [
41,
42]. In addition, some sHSP contain W-boxes on their promoter regions [
43], suggesting the transcriptional activation of
sHSP genes by
WRKY transcription factors. We observed a notable induction of the transcription factors
WRKY-17, -40 and -60 in the WT plants after 24 h of under salt stress. However, in the
35S::OpsHSP18 over-expressing lines, the expression level of these transcription factors was altered under stress. In particular,
WRKY-17 and -60 were more induced in the
35S::OpsHSP18-3 line than in the WT under stress; perhaps one of the mechanisms that lead stress tolerance in this line during germination under salt stress might be related to
WRKY expression.
4. Experimental Section
4.1. Plant Material and Growth Conditions
Seeds of Arabidopsis thaliana Columbia ecotype (Col-0) and seeds of the 35S::OpsHSP18 over-expressing lines generated in this study (see below) were sterilized with 20% (v/v) commercial sodium hypochlorite (6% free chlorine) solution for 5 min, and rinsed five times in sterile distilled water. Aseptic seeds were germinated in plastic Petri dishes containing Murashige and Skoog (MS) 0.5× medium supplemented with 1.4% agar and 1.5% sucrose. Plates were kept at 4 °C for 3 days and then, incubated at 22 ± 2 °C for 10 days in a growth chamber under a 16 h light/8 h dark photoperiod. Plants were grown to maturity in soil pots in a growth chamber at 22 ± 2 °C with a 16 h light/8 h dark photoperiod.
4.2. Inverse Polymerase Chain Reaction (IPCR)
Genomic DNA was isolated from O. streptacantha seedlings using the DNeasy Plant Mini Kit (Qiagen, Hilden, Germany), according to the manufacturer’s instructions. Then, 0.5 μg of genomic DNA were digested with 20 units of EcoRI at 37 °C, in an overnight incubation. The enzyme was then heat inactivated (75 °C for 20 min), and self-ligation of cohesive ends was conducted 16 °C (overnight incubation) with 0.2 units of T4 DNA ligase in a total volume of 100 μL. PCR was performed with 1 μL of the ligation mixture, 1 μL of InvFwOpsHSP18 5′-GATATCTCTGGGAACTAAAGGGG-3′ and InvRvOpsHSP18 5′-AAAGAAGCTTGGAATTAGCGAC-3′ primers (20 μM each one), 10× Advantage 2 PCR buffer, 50× dNTPs (10 mM each one) and 1 unit of 50× Advantage 2 polymerase mix (Clontech, Mountain View, CA, USA) in a total volume of 50 μL. PCR conditions were as follows: 94 °C for 5 min; 94 °C for 2 min, 58 °C for 2 min, and 72 °C for 3 min (35 cycles); and 72 °C for 10 min. A second amplification was done using 1 μL of the previous PCR reaction. The PCR products were cloned into pCR®4-TOPO® vector (Invitrogen, Carlsbad, CA, USA) and sequenced in the Macrogen Company (Seoul, Korea).
4.3. Generation of Arabidopsis thaliana Over-Expressing Lines
The ORF sequence (486 pb) of the
OpsHSP18 gene was amplified from a cDNA library of
O. streptacantha cladodes generated under abiotic stress conditions using the primers Fw
OpsHSP18 5′-ACCATGTCGCTAATTCCAAG-3′ and Rv
OpsHSP18 5′-CCTTTAGTTCCCAGAGATATC-3′ [
24]. The amplified PCR product was cloned into pCR
®8/GW/TOPO
® (Invitrogen, Carlsbad, CA, USA) entry vector, and subcloned into the pMDC32 expression vector by site-specific recombination using Gateway LR Clonase II Enzyme Mix (Invitrogen, Carlsbad, CA, USA). Then, the construct was transformed into
Agrobacterium tumefaciens GV2260 strain by electroporation and, was introduced into
A. thaliana Col-0 plants by floral dip method [
44]. Transgenic lines carrying the
OpsHSP18 gene (
35S::OpsHSP18) were selected on MS 0.5× medium containing hygromycin 50 mg/mL. The T2 generation of transgenic plants was transferred into soil pots and grown in growth chambers under controlled conditions to produce seeds. Homozygous transgenic lines (T3) were used for subsequent analysis of seed germination and tolerance assays under abiotic stress conditions.
4.4. RNA Isolation, RT-PCR and Real-Time qRT-PCR Analyses
Total RNA was isolated from leaves of A. thaliana Col-0 (WT) and 35S::OpsHSP18 transgenic plants, using the Concert™ Plant RNA Reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. cDNA synthesis were performed using the SuperScript™ First Strand Synthesis System for reverse transcriptase polymerase chain reaction (RT-PCR; Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. One microliter of the RT reaction was used as template for PCR assays to amplify the OpsHSP18 gene using the following primers: FwOpsHSP18 5′-ACCATGTCGCTAATTCCAAG-3′ and RvOpsHSP18 5′-CCTTTAGTTCCCAGAGATATC-3′. As loading control, the A. thaliana Actin8 (At1g49240) transcript was amplified using the FwAct 5′-GCCAGTGGTCGTACAACCG-3′ and RvAct 5′-CACGACCAGCAAGGTCGAGACG-3′. PCR amplifications were performed in 25 μL reaction mixtures containing 20 mM Tris-HCl (pH 8.4), 50 mM KCl, 3.5 mM MgCl2, 200 μM dNTPs, 0.2 μM each primer, 2.5 U Taq polymerase (Invitrogen, Carlsbad, CA, USA) and 1 μL RT reaction as template. The amplifications were carried out with different numbers of cycles to ensure a linear response in the PCR reaction. Amplification conditions for both genes were as follows: 5 min at 94 °C followed by 28–30 cycles (depending on the transcript) of 30 s at 94 °C; 45 s at 58 °C; 45 s at 72 °C; and 5 min at 72 °C. PCR analyses were repeated twice, using 3 biological replicates, obtaining similar results. Regarding real-time RT-PCR, experiments were performed in a 10 μL reaction mixture made up of 5 μL of Power SYBR® Green RT-PCR Mix (2×), 200 nM of each oligonucleotide, 50 ng of RNA template and 0.08 μL of RT Enzyme Mix (125×) for one-step RT-PCR following manufacturer’s suggestions (Applied Biosystems), and using the StepOne Real-Time PCR Detection System and StepOne Software v2.1 (Applied Biosystems). The thermal cycling conditions consisted of 30 min at 48 °C (cDNA synthesis), 10 min at 95 °C (activation of AmpliTaq Gold® DNA polymerase), followed by 40 cycles for PCR cycling of 15 s at 95 °C for denature and anneal/extend of 1 min at 60 °C. For each sample, two biological replicates (n = 2) were analyzed with their respective technical replicates. Quantification of WRKY-17 (At2g24570), -40 (At1g80840) and -60 (At2g25000) gene expression was based on a cycle threshold value and transcript data were normalized to the Actin8 (At1g49240) gene values. Relative expression levels of WRKY-17, -40, and -60 in WT and the overexpressing lines with or without salt treatments, were calculated based on corresponding levels in WT plants without salt treatment. Absence of contaminant genomic DNA was confirmed by reactions in which no RT Enzyme Mix reverse transcriptase was added, and with primers for the Actin8 gene which were designed in flanking exons. The primers used were: FwWRKY17 5′-GAGAAATAGAGGGGTTGGTTTTG-3′ and RvWRKY17 5′-CATCATTTTCTTACATGACACCAC-3′ for WRKY-17 transcript, FwWRKY40 5′-AAATCAGCCCTCCCAAGAAACG-3′ and RvWRKY40 5′-CTTCACGACAGTCTCTTCTCTCTGC-3′ for WRKY-40 transcript, and FwWRKY60 5′-GGTGGGCTTGAACCAGTTGAGG-3′ and RvWRKY60 5′-AATCTCCCGGAAATAGCAGTCG-3′ for WRKY-60 transcript.
4.5. Germination Assays under Stress and Hormone Treatments
Seeds of
A. thaliana ecotype Col-0 and of the
35S::OpsHSP18 over-expressing lines (T3) were germinated under different stress conditions and in presence of the phytohormone abscisic acid (ABA). The effect of salt stress on germination was evaluated on MS 0.5× medium supplemented with 0, 100, 125, 150, and 175 mM NaCl. The effect of osmotic stress on germination was assessed on MS 0.5× medium without sucrose and supplemented with glucose (0, 277, 333, and 388 mM) or mannitol (0, 274, 329, and 383 mM). These concentrations were equivalent to 0%, 5%, 6%, and 7% of each sugar [
45]. In addition, seeds were germinated in presence of different concentrations of ABA (0, 3, 5, 7, and 9 μM). The germination assays were carried out using 20 seeds of the control (Col-0) and of the transgenic lines (
35S::OpsHSP18-3, -6, and -7) per treatment. Data are mean ± SE (
n = 20) from four biological replicates. The seeds were regarded as germinated when the radicle emerged from the seed coat. In addition, green cotyledon number was scored after 21 days of NaCl, glucose, mannitol and ABA treatments.
4.6. Estimation of Seedling Survival Rates under Stress Treatments
Seeds of A. thaliana Col-0 and of the 35S::OpsHSP18-3, -6, and -7 transgenic lines (T3 homozygous lines) were germinated in vitro in MS 0.5× Petri dishes in a growth chamber (22 ± 2 °C; 16 h light/8 h dark photoperiod). Subsequently, 10 day-old seedlings (ten plants of WT and ten plants of each one of the transgenic lines) were transferred to MS 1× medium supplemented with 0 and 150 mM NaCl for 14 days. In addition, for osmotic stress assays, seedlings were grown on MS 1× plates (without sucrose) containing 388 mM (7%) and 274 mM (5%) of glucose and mannitol, respectively, for 21 days. After these periods of saline and osmotic stress, the seedlings were transferred to pots, grown under controlled conditions (22 ± 2 °C; 16 h light/8 h dark photoperiod) and irrigated every 3 days, to observe post-stress recovery. The survival rate of plants was calculated by counting the number of plants that survived in each pot, after a period of 21 days. Data are mean ± SE (n = 10) from five biological replicates. The phenotype of each line was documented photographically.
4.7. In Silico Analysis of the OpsHSP18 Protein
The molecular weight and the isoelectric point of the OpsHSP18 protein was predicted using the Compute pI/Mw tool of the Expasy proteomics server at the Swiss Institute of Bioinformatics [
46]. The search of homologous sequences of the OpsHSP18 protein were conducted using the blast and keyword search tools in the Phytozome database [
47] and the BLAST program (BlastN and BlastX) in two databases (GenBank non-redundant (nr) and EST) of the National Center for Biotechnology Information [
48]. Putative protein domains were identified using the Motif Scan tool [
49] at the Expasy proteomics server. Multiple protein sequence alignments were carried out using the CLUSTAL W and the T-Coffee programs at the European Bioinformatics Institute [
50]. The phylogenetic tree was created by Neighbor joining method using the application of the PHYLIP (Phylogeny Inference Package) 3.67 package [
28]. For this, the amino acid sequences of the OpsHSP18 protein and the 19 sHSPs of
A. thaliana [
18] were aligned using MUSCLE version 4.0 version of the EBI database using default values. Then, the aligned sequences were subjected to re-sampling with replacement (1000 bootstrap) using the Seqboot program of PHYLIP package and a distance matrix was calculated with Protdist program using the Henikoff/Tillier [
51]. Probability Matrix from Blocks (PMB). The resultant matrices were transformed into multiple trees by the Neighbor program and summarized by the program Consense (both programs of the PHYLIP 3.67 package). The majority rule consensus tree was edited with the MEGA version 5.0 program [
52].
4.8. Statistical Analysis
The data of seed germination and survival experiments of WT and 35S::OpsHSP18-3, -6, and -7 transgenic lines (T3) were evaluated by One-way ANOVA analysis and Tukey test using GraphPad Software. The data are presented as the mean ± standard error. Differences at p ≤ 0.05 were considered significant.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










