Low Oxygen Tension Maintains Multipotency, Whereas Normoxia Increases Differentiation of Mouse Bone Marrow Stromal Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of BMSC Cultured in 21% O2
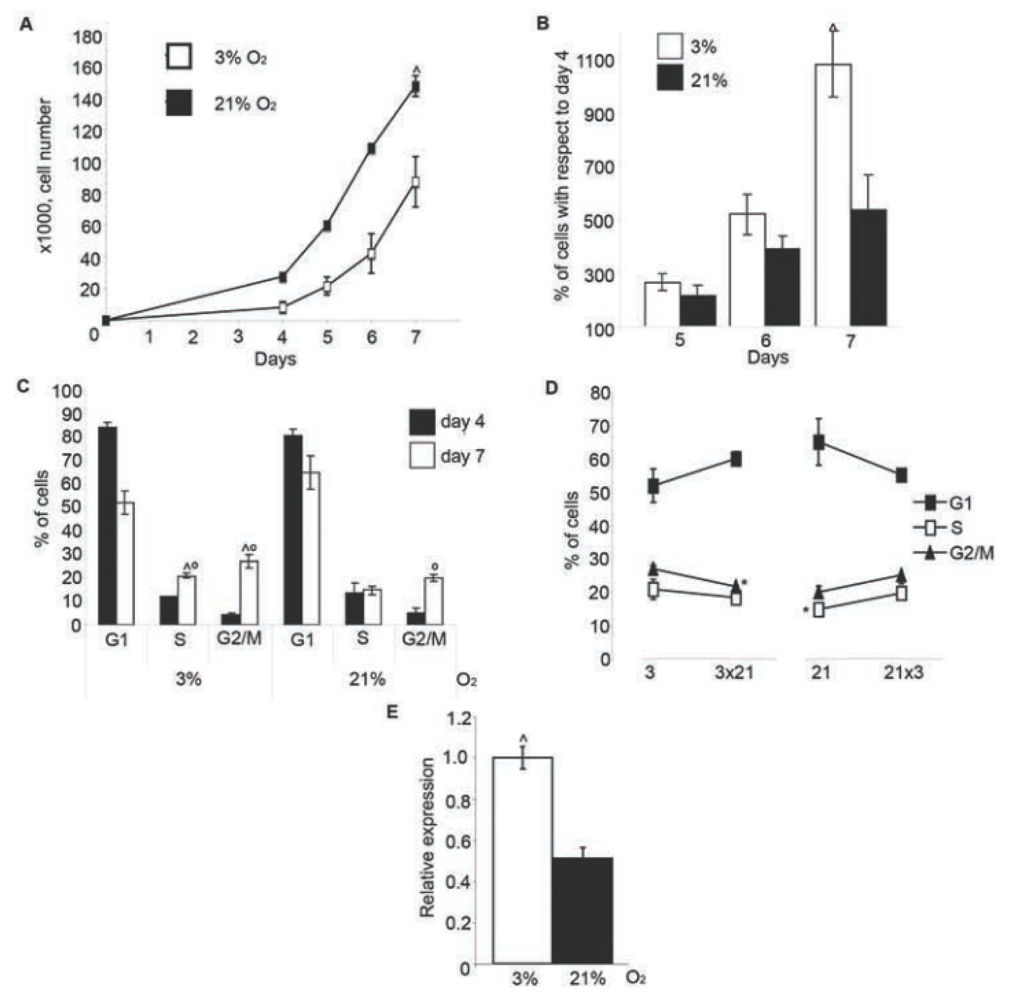
2.2. BMSC Proliferation Enhanced in 3% O2
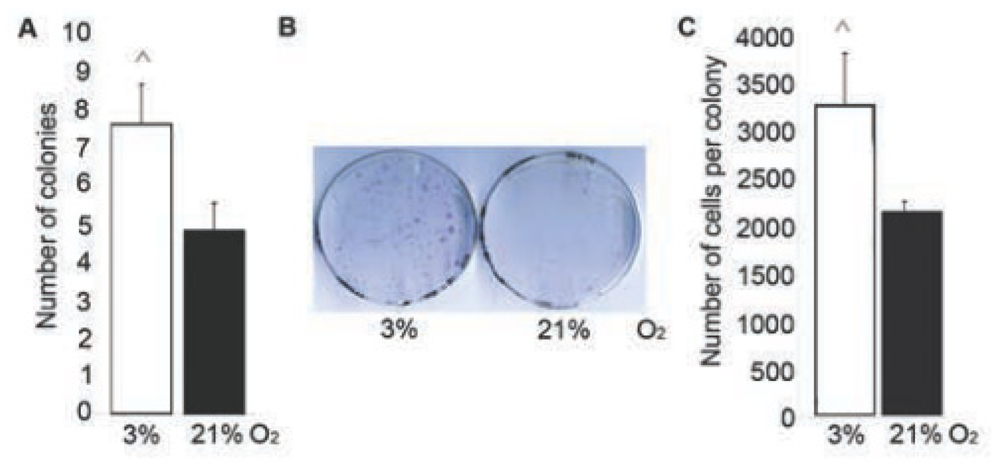
2.3. Number and Proliferation of MSC Increased at 3% O2
2.4. 3% O2 Inhibited Differentiation of BMSC
2.5. 3% O2 Enhanced Pluripotency Markers Expression in BMSC
2.6. 3% O2 Decreases the Cycling Fraction of BMSC
2.7. 3% O2 Affected BMSC Differentiation through ROS and p53 Independent Mechanisms
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. BMSC Isolation and Amplification
4.3. Differentiation
4.4. CFU-A
4.5. PKH-26 Staining
4.6. Total RNA Extraction
4.7. CDNA Synthesis and qPCR
4.8. CD Profiling
4.9. Cell Cycle Analysis
4.10. ROS Analysis
4.11. Statistical Analysis
Supplementary Information
ijms-14-02119-s001.pdfAcknowledgments
Abbreviations
| BMSC | bone marrow stromal cells |
| CFU-A | colony-forming unit assay |
| MSC | mesenchymal stem cells |
| NAC | N-acetylcysteine |
| ROS | reactive oxygen species |
References
- Rosova, I.; Dao, M.; Capoccia, B.; Link, D.; Nolta, J.A. Hypoxic preconditioning results in increased motility and improved therapeutic potential of human mesenchymal stem cells. Stem Cells 2008, 26, 2173–2182. [Google Scholar]
- Annabi, B.; Lee, Y.T.; Turcotte, S.; Naud, E.; Desrosiers, R.R.; Champagne, M.; Eliopoulos, N.; Galipeau, J.; Beliveau, R. Hypoxia promotes murine bone-marrow-derived stromal cell migration and tube formation. Stem Cells 2003, 21, 337–347. [Google Scholar]
- Ivanovic, Z.; Hermitte, F.; Brunet de la Grange, P.; Dazey, B.; Belloc, F.; Lacombe, F.; Vezon, G.; Praloran, V. Simultaneous maintenance of human cord blood SCID-repopulating cells and expansion of committed progenitors at low O2 concentration (3%). Stem Cells 2004, 22, 716–724. [Google Scholar]
- Danet, G.H.; Pan, Y.; Luongo, J.L.; Bonnet, D.A.; Simon, M.C. Expansion of human SCID-repopulating cells under hypoxic conditions. J. Clin. Invest 2003, 112, 126–135. [Google Scholar]
- Pan, Y.; Oprysko, P.R.; Asham, A.M.; Koch, C.J.; Simon, M.C. p53 cannot be induced by hypoxia alone but responds to the hypoxic microenvironment. Oncogene 2004, 23, 4975–4983. [Google Scholar]
- Prockop, D.J. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar]
- Peister, A.; Mellad, J.A.; Larson, B.L.; Hall, B.M.; Gibson, L.F.; Prockop, D.J. Adult stem cells from bone marrow (MSCs) isolated from different strains of inbred mice vary in surface epitopes, rates of proliferation, and differentiation potential. Blood 2004, 103, 1662–1668. [Google Scholar]
- Phinney, D.G.; Kopen, G.; Isaacson, R.L.; Prockop, D.J. Plastic adherent stromal cells from the bone marrow of commonly used strains of inbred mice: Variations in yield, growth, and differentiation. J. Cell Biochem 1999, 72, 570–585. [Google Scholar]
- Ivanovic, Z. Hypoxia or in situ normoxia: The stem cell paradigm. J. Cell. Physiol 2009, 219, 271–275. [Google Scholar]
- Grayson, W.L.; Zhao, F.; Izadpanah, R.; Bunnell, B.; Ma, T. Effects of hypoxia on human mesenchymal stem cell expansion and plasticity in 3D constructs. J. Cell. Physiol 2006, 207, 331–339. [Google Scholar]
- Fehrer, C.; Brunauer, R.; Laschober, G.; Unterluggauer, H.; Reitinger, S.; Kloss, F.; Gully, C.; Gassner, R.; Lepperdinger, G. Reduced oxygen tension attenuates differentiation capacity of human mesenchymal stem cells and prolongs their lifespan. Aging Cell 2007, 6, 745–757. [Google Scholar]
- Ren, H.; Cao, Y.; Zhao, Q.; Li, J.; Zhou, C.; Liao, L.; Jia, M.; Zhao, Q.; Cai, H.; Han, Z.C.; et al. Proliferation and differentiation of bone marrow stromal cells under hypoxic conditions. Biochem. Biophys. Res. Commun 2006, 347, 12–21. [Google Scholar]
- D’Ippolito, G.; Diabira, S.; Howard, G.A.; Roos, B.A.; Schiller, P.C. Low oxygen tension inhibits osteogenic differentiation and enhances stemness of human MIAMI cells. Bone 2006, 39, 513–522. [Google Scholar]
- Potier, E.; Ferreira, E.; Andriamanalijaona, R.; Pujol, J.P.; Oudina, K.; Logeart-Avramoglou, D.; Petite, H. Hypoxia affects mesenchymal stromal cell osteogenic differentiation and angiogenic factor expression. Bone 2007, 40, 1078–1087. [Google Scholar]
- Lennon, D.P.; Edmison, J.M.; Caplan, A.I. Cultivation of rat marrow-derived mesenchymal stem cells in reduced oxygen tension: Effects on in vitro and in vivo osteochondrogenesis. J. Cell. Physiol 2001, 187, 345–355. [Google Scholar]
- Malladi, P.; Xu, Y.; Chiou, M.; Giaccia, A.J.; Longaker, M.T. Effect of reduced oxygen tension on chondrogenesis and osteogenesis in adipose-derived mesenchymal cells. Am. J. Physiol. Cell Physiol 2006, 290, C1139–C1146. [Google Scholar]
- Fink, T.; Abildtrup, L.; Fogd, K.; Abdallah, B.M.; Kassem, M.; Ebbesen, P.; Zachar, V. Induction of adipocyte-like phenotype in human mesenchymal stem cells by hypoxia. Stem Cells 2004, 22, 1346–1355. [Google Scholar]
- Valorani, M.G.; Germani, A.; Otto, W.R.; Harper, L.; Biddle, A.; Khoo, C.P.; Lin, W.R.; Hawa, M.I.; Tropel, P.; Patrizi, M.P.; et al. Hypoxia increases Sca-1/CD44 co-expression in murine mesenchymal stem cells and enhances their adipogenic differentiation potential. Cell Tissue Res 2010, 341, 111–120. [Google Scholar]
- Berniakovich, I.; Trinei, M.; Stendardo, M.; Migliaccio, E.; Minucci, S.; Bernardi, P.; Pelicci, P.G.; Giorgio, M. p66Shc-generated oxidative signal promotes fat accumulation. J. Biol. Chem 2008, 283, 34283–34293. [Google Scholar]
- Cicalese, A.; Bonizzi, G.; Pasi, C.E.; Faretta, M.; Ronzoni, S.; Giulini, B.; Brisken, C.; Minucci, S.; Di Fiore, P.P.; Pelicci, P.G. The tumor suppressor p53 regulates polarity of self-renewing divisions in mammary stem cells. Cell 2009, 138, 1083–1095. [Google Scholar]
- Chandel, N.S.; Vander Heiden, M.G.; Thompson, C.B.; Schumacker, P.T. Redox regulation of p53 during hypoxia. Oncogene 2000, 19, 3840–3848. [Google Scholar]
- Denker, A.E.; Nicoll, S.B.; Tuan, R.S. Formation of cartilage-like spheroids by micromass cultures of murine C3H10T1/2 cells upon treatment with transforming growth factor-beta 1. Differentiation 1995, 59, 25–34. [Google Scholar]
- da Meirelles, L.S.; Nardi, N.B. Murine marrow-derived mesenchymal stem cell: Isolation, in vitro expansion, and characterization. Br. J. Haematol 2003, 123, 702–711. [Google Scholar]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp. Hematol 1976, 4, 267–274. [Google Scholar]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar]
- Okuyama, H.; Krishnamachary, B.; Zhou, Y.F.; Nagasawa, H.; Bosch-Marce, M.; Semenza, G.L. Expression of vascular endothelial growth factor receptor 1 in bone marrow-derived mesenchymal cells is dependent on hypoxia-inducible factor 1. J. Biol. Chem 2006, 281, 15554–15563. [Google Scholar]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell 1994, 79, 1147–1156. [Google Scholar]
- Bernlohr, D.A.; Angus, C.W.; Lane, M.D.; Bolanowski, M.A.; Kelly, T.J., Jr. Expression of specific mRNAs during adipose differentiation: Identification of an mRNA encoding a homologue of myelin P2 protein. Proc. Natl. Acad. Sci. USA 1984, 81, 5468–5472. [Google Scholar]
- Cook, K.S.; Hunt, C.R.; Spiegelman, B.M. Developmentally regulated mRNAs in 3T3-adipocytes: Analysis of transcriptional control. J. Cell Biol 1985, 100, 514–520. [Google Scholar]
- Nichols, J.; Zevnik, B.; Anastassiadis, K.; Niwa, H.; Klewe-Nebenius, D.; Chambers, I.; Scholer, H.; Smith, A. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell 1998, 95, 379–391. [Google Scholar]
- Rosfjord, E.; Rizzino, A. The octamer motif present in the Rex-1 promoter binds Oct-1 and Oct-3 expressed by EC cells and ES cells. Biochem. Biophys. Res. Commun 1994, 203, 1795–1802. [Google Scholar]
- Kim, J.H.; Song, S.Y.; Park, S.G.; Song, S.U.; Xia, Y.; Sung, J.H. Primary involvement of NADPH oxidase 4 in hypoxia-induced generation of reactive oxygen species in adipose-derived stem cells. Stem Cells Dev 2012, 21, 2212–2221. [Google Scholar]
- Wilson, A.; Laurenti, E.; Oser, G.; van der Wath, R.C.; Blanco-Bose, W.; Jaworski, M.; Offner, S.; Dunant, C.F.; Eshkind, L.; Bockamp, E.; et al. Hematopoietic stem cells reversibly switch from dormancy to self-renewal during homeostasis and repair. Cell 2008, 135, 1118–1129. [Google Scholar]
- Fuchs, E.; Segre, J.A. Stem cells: A new lease on life. Cell 2000, 100, 143–155. [Google Scholar]
- Migliaccio, E.; Giorgio, M.; Mele, S.; Pelicci, G.; Reboldi, P.; Pandolfi, P.P.; Lanfrancone, L.; Pelicci, P.G. The p66shc adaptor protein controls oxidative stress response and life span in mammals. Nature 1999, 402, 309–313. [Google Scholar]
- Jurgensen, J.S.; Rosenberger, C.; Wiesener, M.S.; Warnecke, C.; Horstrup, J.H.; Grafe, M.; Philipp, S.; Griethe, W.; Maxwell, P.H.; Frei, U.; et al. Persistent induction of HIF-1alpha and -2alpha in cardiomyocytes and stromal cells of ischemic myocardium. FASEB J 2004, 18, 1415–1417. [Google Scholar]
- Stevens, H.E. Allergic and inflammatory aspects of chronic rhinosinusitis. J. Otolaryngol 1991, 20, 395–399. [Google Scholar]
- Vaupel, P.; Kallinowski, F.; Okunieff, P. Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: A review. Cancer Res 1989, 49, 6449–6465. [Google Scholar]
- Chow, D.C.; Wenning, L.A.; Miller, W.M.; Papoutsakis, E.T. Modeling pO(2) distributions in the bone marrow hematopoietic compartment. I. Krogh’s model. Biophys. J 2001, 81, 675–684. [Google Scholar]
- Parmar, K.; Mauch, P.; Vergilio, J.A.; Sackstein, R.; Down, J.D. Distribution of hematopoietic stem cells in the bone marrow according to regional hypoxia. Proc. Natl. Acad. Sci. USA 2007, 104, 5431–5436. [Google Scholar]
- Rochefort, G.Y.; Delorme, B.; Lopez, A.; Herault, O.; Bonnet, P.; Charbord, P.; Eder, V.; Domenech, J. Multipotential mesenchymal stem cells are mobilized into peripheral blood by hypoxia. Stem Cells 2006, 24, 2202–2208. [Google Scholar]
- Goldstein, L.J.; Gallagher, K.A.; Bauer, S.M.; Bauer, R.J.; Baireddy, V.; Liu, Z.J.; Buerk, D.G.; Thom, S.R.; Velazquez, O.C. Endothelial progenitor cell release into circulation is triggered by hyperoxia-induced increases in bone marrow nitric oxide. Stem Cells 2006, 24, 2309–2318. [Google Scholar]
- Bezy, O.; Vernochet, C.; Gesta, S.; Farmer, S.R.; Kahn, C.R. TRB3 blocks adipocyte differentiation through the inhibition of C/EBPbeta transcriptional activity. Mol. Cell. Biol 2007, 27, 6818–6831. [Google Scholar]






© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Berniakovich, I.; Giorgio, M. Low Oxygen Tension Maintains Multipotency, Whereas Normoxia Increases Differentiation of Mouse Bone Marrow Stromal Cells. Int. J. Mol. Sci. 2013, 14, 2119-2134. https://doi.org/10.3390/ijms14012119
Berniakovich I, Giorgio M. Low Oxygen Tension Maintains Multipotency, Whereas Normoxia Increases Differentiation of Mouse Bone Marrow Stromal Cells. International Journal of Molecular Sciences. 2013; 14(1):2119-2134. https://doi.org/10.3390/ijms14012119
Chicago/Turabian StyleBerniakovich, Ina, and Marco Giorgio. 2013. "Low Oxygen Tension Maintains Multipotency, Whereas Normoxia Increases Differentiation of Mouse Bone Marrow Stromal Cells" International Journal of Molecular Sciences 14, no. 1: 2119-2134. https://doi.org/10.3390/ijms14012119
APA StyleBerniakovich, I., & Giorgio, M. (2013). Low Oxygen Tension Maintains Multipotency, Whereas Normoxia Increases Differentiation of Mouse Bone Marrow Stromal Cells. International Journal of Molecular Sciences, 14(1), 2119-2134. https://doi.org/10.3390/ijms14012119