Advanced Knowledge of Three Important Classes of Grape Phenolics: Anthocyanins, Stilbenes and Flavonols



 and
and
Abstract
:1. Introduction
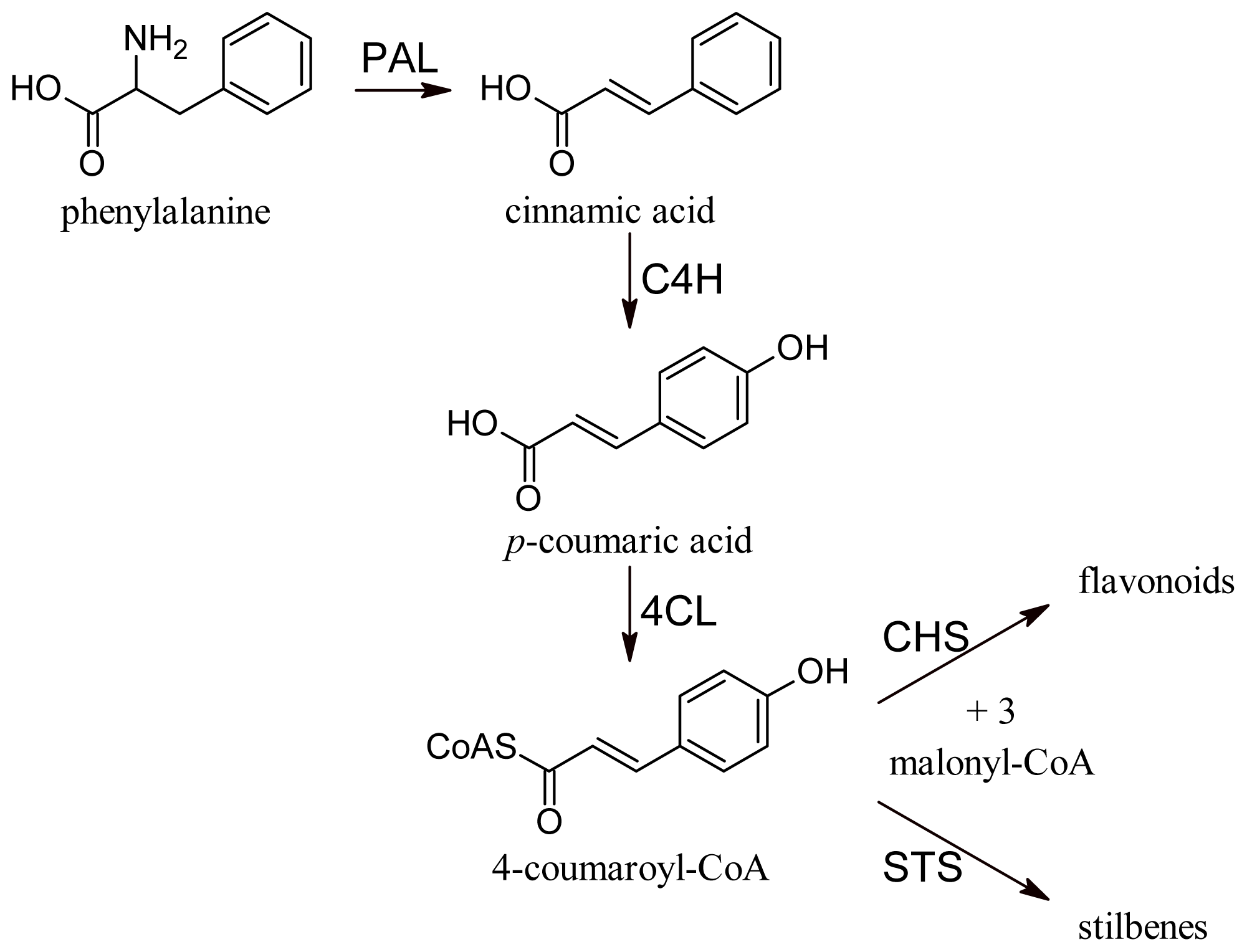
2. Biosynthesis of Grape Polyphenols
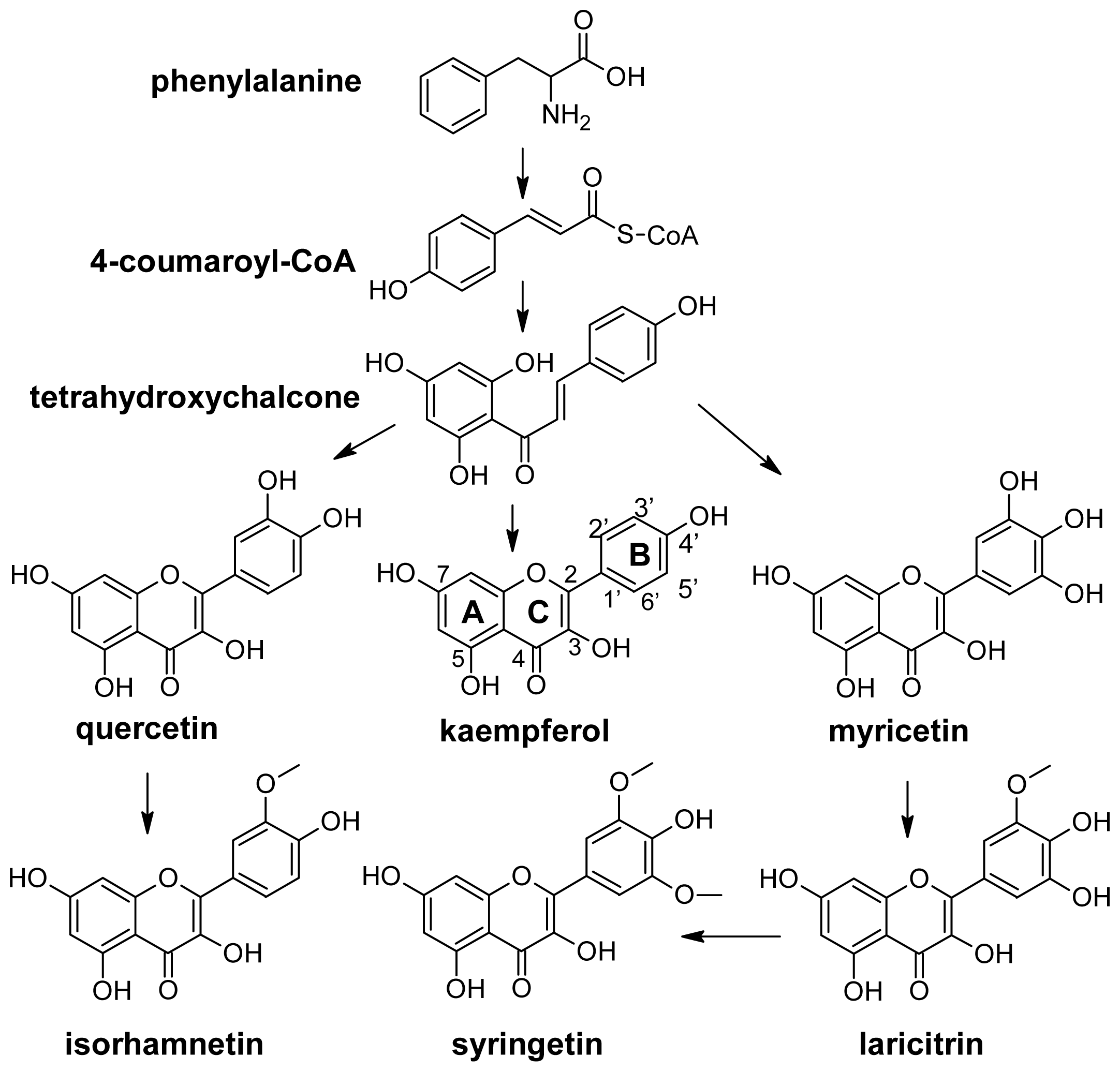
2.1. Flavonoids
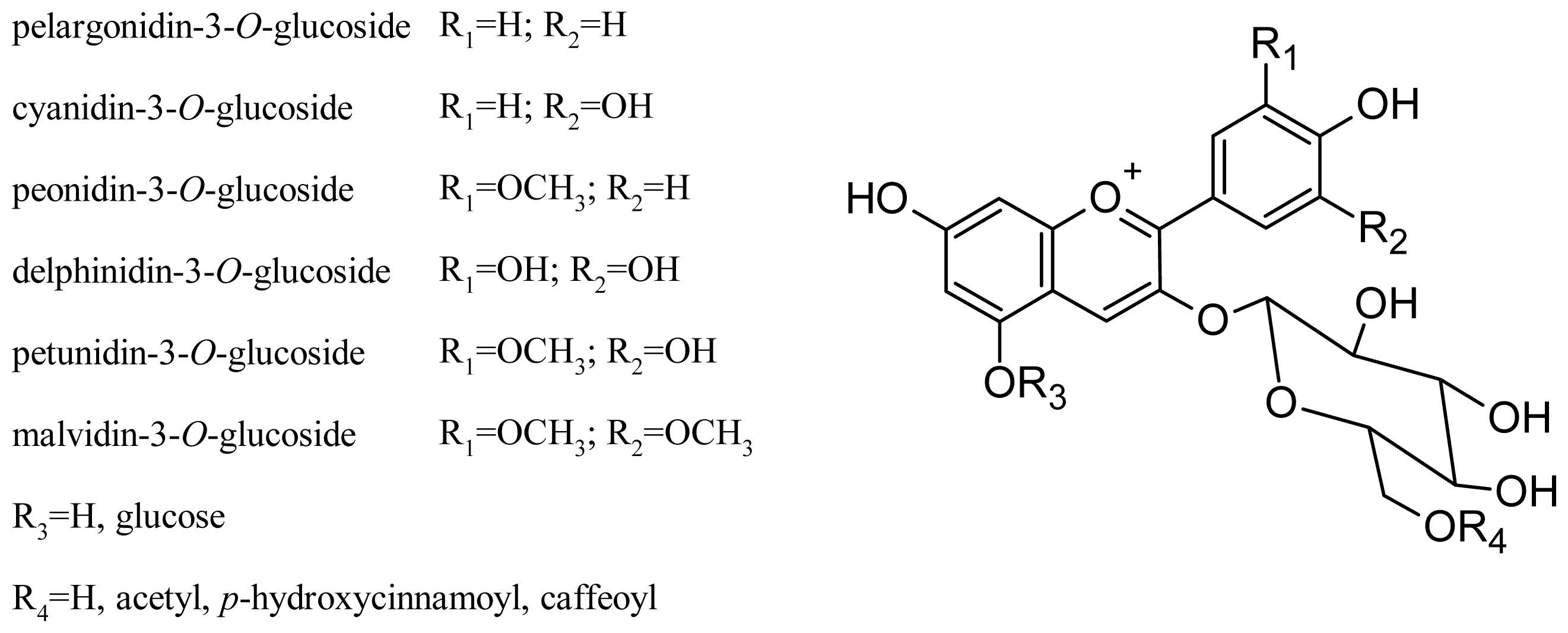
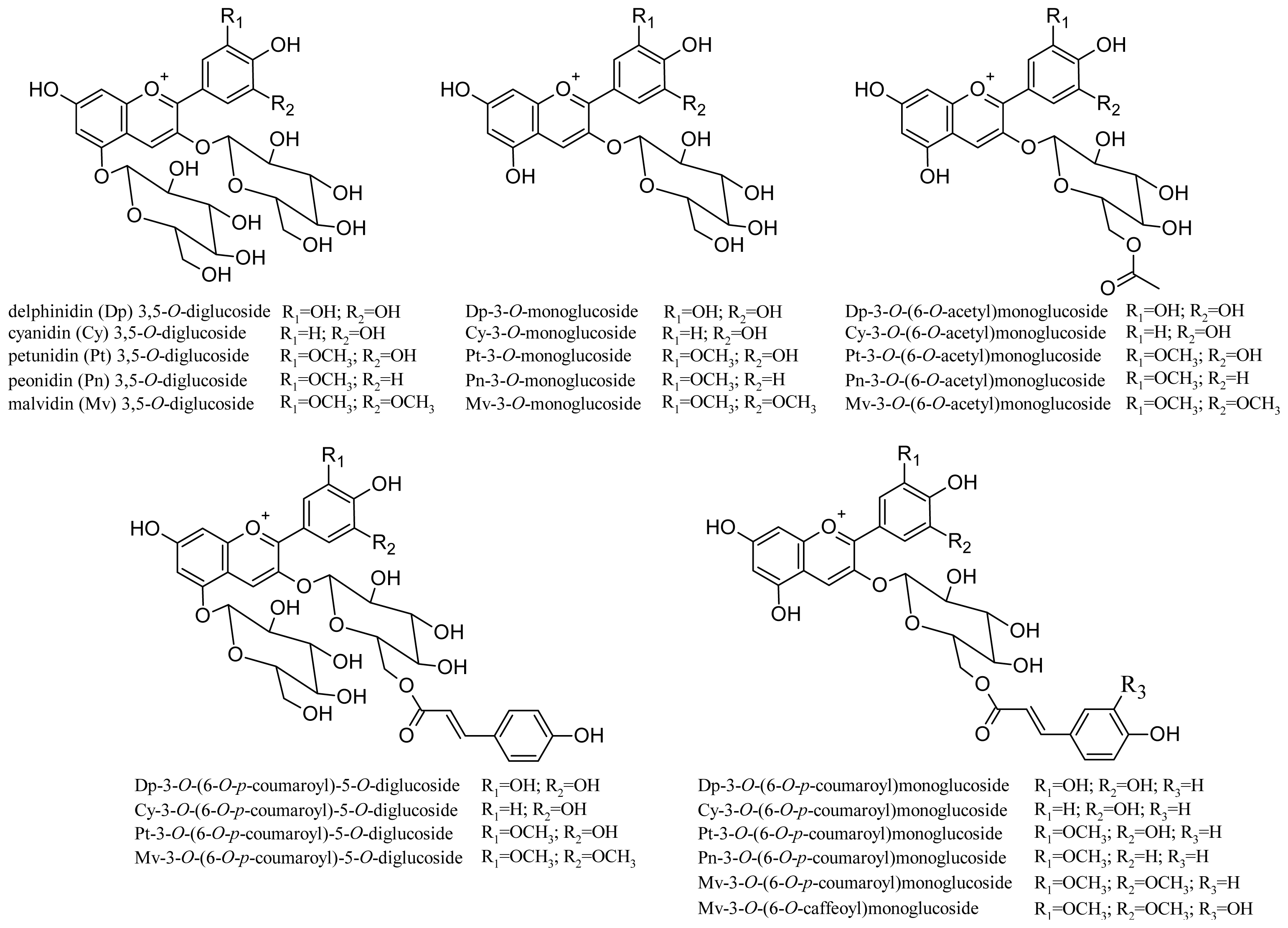
2.2. Anthocyanins and Flavonols
2.3. Stilbene Derivatives
2.4. Effects of Agrochemicals and Plant Activators on Grape Polyphenols
3. Chemistry and Proprieties of Anthocyanins
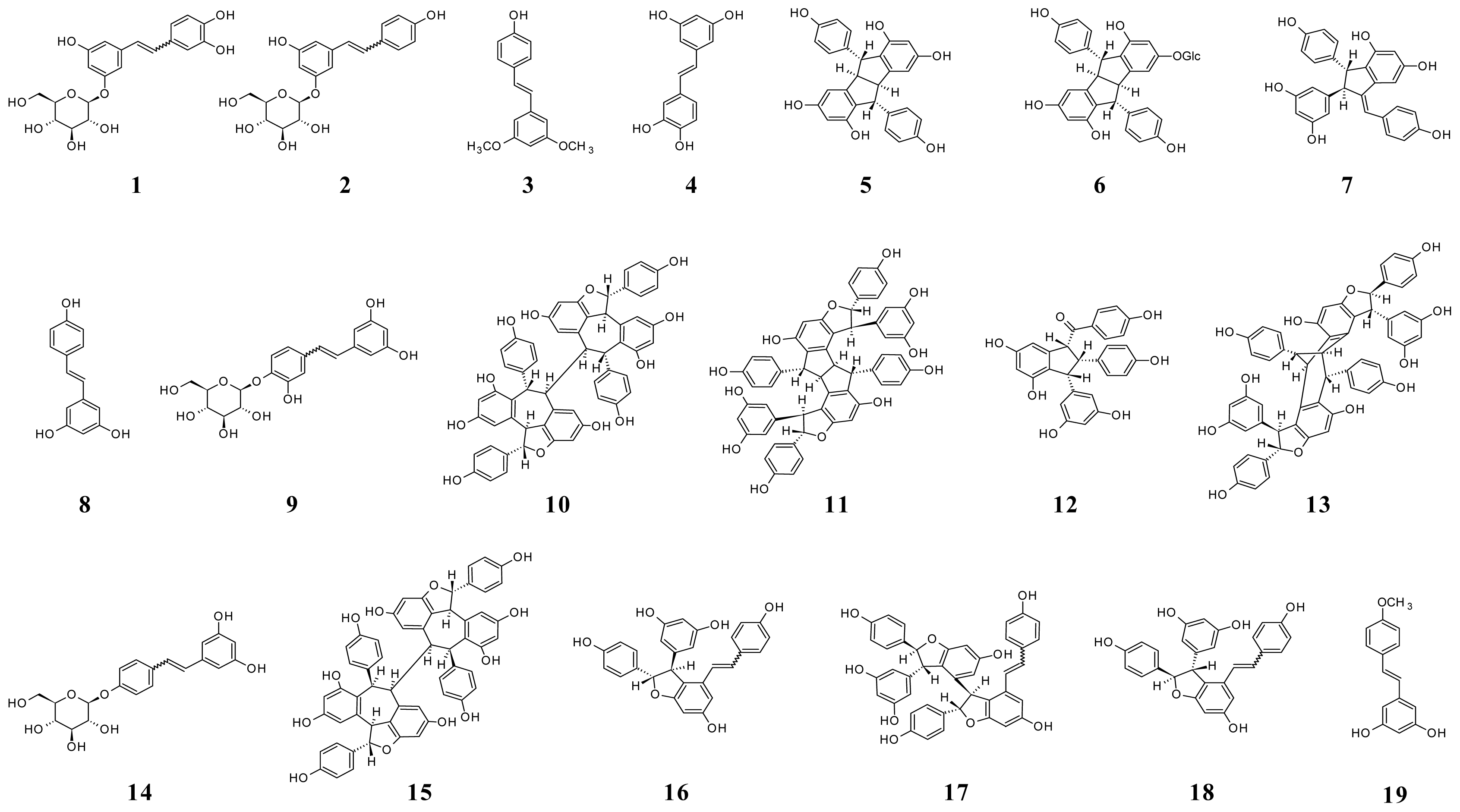
4. Chemistry and Proprieties of Stilbene Derivatives
5. Chemistry and Proprieties of Flavonols
6. Conclusions







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| m/z | Assignment |
|---|---|
| 287(F) | Cy |
| 301(F) | Pn |
| 303(F) | Dp |
| 317(F) | Pt |
| 331(F) | Mv |
| 449(M) | Cy + G |
| 463(M) | Pn + G |
| 465(M) | Dp + G |
| 479(M) | Pt + G |
| 493(M) | Mv + G |
| 617(F) | MvCy |
| 631(F) | MvPn |
| 633(F) | MvDp |
| 647(F) | MvPt |
| 661(F) | MvMv |
| 779(F) | MvCy + G |
| 793(F) | MvPn + G |
| 795(F) | MvDp + G |
| 809(F) | MvPt + G |
| 823(F) | MvMv + G |
| 941(M) | MvCy + 2G |
| 955(M) | MvPn + 2G |
| 957(M) | MvDp + 2G |
| 971(M) | MvPt + 2G |
| 985(M) | MvMv + 2G |
| 1087(M) | MvCy + G·pCG |
| 1101(M) | MvPn + G·pCG |
| 1103(M) | MvDp + G·pCG |
| 1117(M) | MvPt + G·pCG |
| 1131(M) | MvMv + G·pCG |
| 1271(F) | MvMvCy + 2G |
| 1285(F) | MvMvPn + 2G |
| 1287(F) | MvMvDp + 2G |
| 1301(F) | MvMvPt + 2G |
| 1315(F) | MvMvMv + 2G |
| 1417(F) | MvMvCy + G·pCG |
| 1431(F) | MvMvPn + G·pCG |
| 1433(F) | MvMvDp + G·pCG |
| 1433(M) | MvMvCy + 3G |
| 1447(F) | MvMvPt + G·pCG |
| 1447(M) | MvMvPn + 3G |
| 1449(M) | MvMvDp + 3G |
| 1461(F) | MvMvMv + G·pCG |
| 1463(M) | MvMvPt + 3G |
| 1477(M) | MvMvMv + 3G |
| 1579(M) | MvMvCy + 2G·pCG |
| 1593(M) | MvMvPn + 2G·pCG |
| 1595(M) | MvMvDp + 2G·pCG |
| 1609(M) | MvMvPt + 2G·pCG |
| 1623(M) | MvMvMv + 2G·pCG |
| Anthocyanin | m/z (M+) | Cultivar |
|---|---|---|
| Cy-3-O-pentoside | 419 | Casavecchia |
| Pg-3-O-glucoside | 433 | Concord, Salvador, Rubired |
| Cy-3-O-(6-O-acetyl)pentoside | 461 | Casavecchia |
| Cy-3-O-(6-O-p-coumaryl)pentoside | 565 | Casavecchia |
| Dp-3-O-glucoside-pyruvic acid | 533 | Isabelle |
| Dp-3-O-(6-O-p-coumaryl)glucoside-pyruvic acid | 679 | Isabelle |
| Pn-3-O-glucoside-acetaldehyde | 487 | Isabelle, Pallagrello |
| Mv-3-O-glucoside-acetaldehyde | 517 | Isabelle |
| Pt-3-O-(6-caffeoyl)-5-O-diglucoside | 803 | Isabelle, Casavecchia |
| Dp-3-O-(6-acetyl)-5-O-diglucoside | 669 | Isabelle |
| Dp-3-O-(6-feruloyl)-5-O-diglucoside | 803 | Isabelle, Casavecchia |
| Pn-3-O-(6-O-p-coumaryl)-5-O-diglucoside | 771 | Concord, Salvador, Isabelle, Casavecchia |
Conflicts of Interest
References
- Flamini, R. Mass spectrometry in grape and wine chemistry. Part I: Polyphenols. Mass Spectrom. Rev 2003, 22, 218–250. [Google Scholar]
- He, J.; Giusti, M.M. Anthocyanins: Natural colorants with health-promoting properties. Annu. Rev. Food Sci. Technol 2010, 1, 163–187. [Google Scholar]
- Niessen, W.M.A.; Tinke, A.P. Liquid chromatography-mass spectrometry. General principles and instrumentation. J. Chromatogr. A 1995, 703, 37–57. [Google Scholar]
- De Hoffmann, E. Tandem mass spectrometry: A primer. J. Mass Spectrom 1996, 31, 125–137. [Google Scholar]
- Abian, J. The coupling of gas and liquid chromatography with mass spectrometry. J. Mass Spectrom 1999, 34, 157–168. [Google Scholar]
- Flamini, R.; Traldi, P. Grape; Polyphenols, Wine. Mass Spectrometry in Grape & Wine Chemistry; John Wiley & Sons Inc: Hoboken, NJ, USA, 2010; pp. 163–225. [Google Scholar]
- Flamini, R.; de Rosso, M.; de Marchi, F.; Dalla Vedova, A.; Panighel, A.; Gardiman, M.; Maoz, I.; Bavaresco, L. An innovative approach to grape metabolomics: Stilbene profiling by suspect screening analysis. Metabolomics 2013. [Google Scholar] [CrossRef]
- Castillo-Muñoz, N.; Gómez-Alonso, S.; García-Romero, E.; Hermosn-Gutiérrez, I. Flavonol profiles of Vitis vinifera red grapes and their single-cultivar wines. J. Agric. Food Chem 2007, 55, 992–1002. [Google Scholar]
- Sparvoli, F.; Martin, C.; Scienza, A.; Gavazzi, G.; Tonelli, C. Cloning and molecular analysis of structural genes involved in flavonoid and stilbene biosynthesis in grape (Vitis vinifera L.). Plant Mol. Biol 1994, 24, 743–755. [Google Scholar]
- Borkowski, T.; Szymusiak, H.; Gliszczyńska-Swiglo, A.; Tyrakowska, B. The effect of 3-O-β-glucosylation on structural transformations of anthocyanidins. Food Res. Int 2005, 38, 1031–1037. [Google Scholar]
- Ford, C.M.; Boss, P.K.; Hoj, P.B. Cloning and characterization of Vitis vinifera UDP-glucose: Flavonoid 3-O-glucosyltransferase, a homologue of the enzyme encoded by the maize Bronze-1 locus that may primarily serve to glucosylate anthocyanidins in vivo. J. Biol. Chem. 1998, 273, 9224–9233. [Google Scholar]
- Gomez, C.; Terrier, N.; Torregrosa, L.; Vialet, S.; Alexandre, F.-L.; Verriès, C.; Souquet, J.-M.; Mazauric, J.-P.; Klein, M.; Cheynier, V.; et al. Grapevine MATE-type proteins act as vacuolar H+-dependent acylated anthocyanin transporters. Plant Physiol 2009, 150, 402–415. [Google Scholar]
- Ageorges, A.; Fernandez, L.; Vialet, S.; Merdinoglu, D.; Terrier, N.; Romieu, C. Four specific isogenes of the anthocyanin metabolic pathway are systematically co-expressed with the red colour of grape berries. Plant Sci 2006, 170, 372–383. [Google Scholar]
- Castellarin, S.D.; di Gaspero, G. Transcriptional control of anthocyanin biosynthetic genes in extreme phenotypes for berry pigmentation of naturally occurring grapevines. BMC Plant Biol 2007, 7, 46. [Google Scholar]
- Braidot, E.; Petrussa, E.; Bertolini, A.; Peresson, C.; Ermacora, P.; Loi, N.; Terdoslavich, M.; Passamonti, S.; Macri, F.; Vianello, A. Evidence for a putative flavonoid translocator similar to mammalian bilitranslocase in grape berries (Vitis vinifera L.) during ripening. Planta 2008, 228, 203–213. [Google Scholar]
- Goto-Yamamoto, N.; Wan, G.H.; Masaki, K.; Kobayashi, S. Structure and transcription of three chalcone synthase genes of grapevine (Vitis vinifera). Plant Sci 2002, 162, 867–872. [Google Scholar]
- Schmidlin, L.; Poutaraud, A.; Claudel, P.; Mestre, P.; Prado, E.; Santos-Rosa, M.; Wiedemann-Merdinoglu, S.; Karst, F.; Merdinoglu, D.; Hugueney, P. A stress-inducible resveratrol O-methyltransferase involved in the biosynthesis of pterostilbene in grapevine. Plant Physiol 2008, 148, 1630–1639. [Google Scholar]
- Hall, D.; de Luca, V. Mesocarp localization of a bi-functional resveratrol/hydroxycinnamic acid glucosyltransferase of Concord grape (Vitis labrusca). Plant J 2007, 49, 579–591. [Google Scholar]
- He, F.; Mu, L.; Yan, G.-L.; Liang, N.-N.; Pan, Q.-H.; Wang, J.; Reeves, M.J.; Duan, C.-Q. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules 2010, 15, 9057–9091. [Google Scholar]
- Ruiz-García, Y.; Romero-Cascales, I.; Gil-Muñoz, R.; Fernández-Fernández, J.I.; López-Roca, J.M.; Gómez-Plaza, E. Improving grape phenolic content and wine chromatic characteristics through the use of two different elicitors: Methyl jasmonate versus benzothiadiazole. J. Agric. Food Chem 2012, 60, 1283–1290. [Google Scholar]
- Ruiz-García, Y.; Gil-Muñoz, R.; López-Roca, J.M.; Martínez-Cutillas, A.; Romero-Cascales, I.; Gómez-Plaza, E. Increasing the phenolic compound content of grapes by preharvest application of abcisic acid and a combination of methyl jasmonate and benzothiadiazole. J. Agric. Food Chem 2013, 61, 3978–3983. [Google Scholar]
- Iriti, M.; Vitalini, S.; di Tommaso, G.; D’Amico, S.; Borgo, M.; Faoro, F. New chitosan formulation prevents grapevine powdery mildew infection and improves polyphenol content and free radical scavenging activity of grape and wine. Aust. J. Grape Wine Res 2011, 17, 263–269. [Google Scholar]
- Ochoa-Villarreal, M.; Vargas-Arispuro, I.; Islas-Osuna, M.A.; González-Aguilar, G.; Martínez-Téllez, M.Á. Pectin-derived oligosaccharides increase color and anthocyanin content in Flame Seedless grapes. J. Sci. Food Agric 2011, 91, 1928–1930. [Google Scholar]
- Dercks, W.; Creasy, L.L. Influence of fosetyl-Al on phytoalexin accumulation in the Plasmopara viticola grapevine interaction. Physiol. Mol. Plant Pathol 1989, 34, 203–213. [Google Scholar]
- Iriti, M.; Rossoni, M.; Borgo, M.; Faoro, F. Benzothiadiazole enhances resveratrol and anthocyanin biosynethesis in grapevine, meanwhile improving resistance to Botrytis cinerea. J. Agric. Food Chem 2004, 52, 4406–4413. [Google Scholar]
- Vezzulli, S.; Civardi, S.; Ferrari, F.; Bavaresco, L. Methyl jasmonate treatment as a trigger of resveratrol synthesis in cultivated grapevine. Am. J. Enol. Viticult 2007, 58, 530–533. [Google Scholar]
- Bavaresco, L.; Fregoni, C.; van Zeller de Macedo Basto Gonçalves, M.I.; Vezzulli, S. Physiology & molecular biology of grapevine stilbenes: An update. In Grapevine Molecular Physiology and Biotechnology; Roubelakis Angelakis, K.A., Ed.; Springer Science + Business Media B.V.: New York, NY, USA, 2009; Volume 1, p. 341. [Google Scholar]
- Iriti, M.; Rossoni, M.; Borgo, M.; Ferrara, L.; Faoro, F. Induction of resistance to gray mold with benzothiadiazole modifies amino acid profile and increases proanthocyanidins in grape: Primary versus secondary metabolism. J. Agric. Food Chem 2005, 53, 9133–9139. [Google Scholar]
- Busam, G.; Junghanns, K.T.; Kneusel, R.E.; Kassemeyer, H.; Matern, U. Characterization and expression of caffeoyl-coenzyme A 3-O-methyltransferase proposed for the induced resistance responce of Vitis vinifera L. Plant Physiol 1997, 115, 1039–1048. [Google Scholar]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr 2005, 81, 230S–242S. [Google Scholar]
- Han, X.; Shen, T.; Lou, H. Dietary polyphenols and their biological significance. Int. J. Mol. Sci 2007, 8, 950–988. [Google Scholar]
- Mattivi, F.; Guzzon, R.; Vrhovsek, U.; Stefanini, M.; Velasco, R. Metabolite profiling of grape: Flavonols and anthocyanins. J. Agric. Food Chem 2006, 54, 7692–7702. [Google Scholar]
- Vidal, S.; Francis, L.; Noble, A.; Kwiatkowski, M.; Cheynier, V.; Waters, E. Taste and mouthfeel properties of different types of tannin-like polyphenolic compounds and anthocyanins in wine. Anal. Chim. Acta 2004, 513, 57–65. [Google Scholar]
- Dufour, C.; Sauvaitre, I. Interactions between anthocyanins and aroma substances in a model system. Effect on the flavor of grape-derived beverages. J. Agric. Food Chem 2000, 48, 1784–1788. [Google Scholar]
- Castillo-Muñoz, N.; Fernández-González, M.; Gómez-Alonso, S.; García-Romero, E.; Hermosín-Gutiérrez, I. Red-color related phenolic composition of Garnacha Tintorera (Vitis vinifera L.) grapes and red wines. J. Agric. Food Chem 2009, 57, 7883–7891. [Google Scholar]
- González-SanJosé, M.L.; Diez, C. Relationship between anthocyanins and sugars during the ripening of grape berries. Food Chem 1992, 43, 193–197. [Google Scholar]
- Di Stefano, R. Chemical methods for varietal characterization. Riv. Vitic. Enol 1996, 49, 51–56. [Google Scholar]
- Mochioka, R.; Yamaguchi, M.; Horiuchi, S.; Matsui, H.; Kurooka, H. Chemotaxonomic classification of native wild Japanese grapes by anthocyanins in berry skin. J. Jpn. Soc. Hortic. Sci 1995, 64, 463–470. [Google Scholar]
- Flamini, R.; Tomasi, D. The Anthocyanin content in berries of the hybrid grape cultivars Clinton and Isabella. Vitis 2000, 39, 79–81. [Google Scholar]
- Favretto, D.; Flamini, R. Application of electrospray ionization mass spectrometry to the study of grape anthocyanins. Am. J. Enol. Viticult 2000, 51, 55–64. [Google Scholar]
- Figueiredo-González, M.; Martínez-Carballo, E.; Cancho-Grande, B.; Santiago, J.L.; Martínez, M.C.; Simal-Gándara, J. Pattern recognition of three Vitis vinifera L. red grapes varieties based on anthocyanin and flavonol profiles, with correlations between their biosynthesis pathways. Food Chem 2012, 130, 9–19. [Google Scholar]
- Garcia-Alonso, M.; Rimbach, G.; Sasai, M.; Nakahara, M.; Matsugo, S.; Uchida, Y.; Rivas-Gonzalo, J.C.; de Pascual-Teresa, S. Electron spin resonance spectroscopy studies on the free radical scavenging activity of wine anthocyanins and pyranoanthocyanins. Mol. Nutr. Food Res 2005, 49, 1112–1119. [Google Scholar]
- Tedesco, I.; Luigi Russo, G.; Nazzaro, F.; Russo, M.; Palumbo, R. Antioxidant effect of red wine anthocyanins in normal and catalase-inactive human erythrocytes. J. Nutr. Biochem 2001, 12, 505–511. [Google Scholar]
- De Pascual-Teresa, S.; Sanchez-Ballesta, M.T. Anthocyanins: From plant to health. Phytochem. Rev 2008, 7, 281–299. [Google Scholar]
- Mazza, G.; Brouillard, R. Recent developments in the stabilization of anthocyanins in food products. Food Chem 1987, 25, 207–225. [Google Scholar]
- Bassa, I.A.; Francis, F.J. Stability of anthocyanins from sweet potatoes in a model beverage. J. Food Sci 1987, 52, 1753–1754. [Google Scholar]
- Janvary, L.; Hoffmann, T.; Pfeiffer, J.; Hausmann, L.; Topfer, R.; Fischer, T.C.; Schwab, W. A double mutation in the anthocyanin 5-O-glucosyltransferase gene disrupts enzymatic activity in Vitis vinifera L. J. Agric. Food Chem 2009, 57, 3512–3518. [Google Scholar]
- Wang, H.; Race, E.J.; Shrikhande, A.J. Characterization of anthocyanins in grape juices by ion trap liquid chromatography-mass spectrometry. J. Agric. Food Chem 2003, 51, 1839–1844. [Google Scholar]
- He, F.; He, J.J.; Pan, Q.H.; Duan, C.Q. Mass-spectrometry evidence confirming the presence of pelargonidin-3-O-glucoside in the berry skins of Cabernet Sauvignon and Pinot Noir (Vitis vinifera L.). Aust. J. Grape Wine Res 2010, 16, 464–468. [Google Scholar]
- Vidal, S.; Meudec, E.; Cheynier, V.; Skouroumounis, G.; Hayasaka, Y. Mass spectrometric evidence for the existence of oligomeric anthocyanins in grape skins. J. Agric. Food Chem 2004, 52, 7144–7151. [Google Scholar]
- Sarni, P.; Fulcrand, H.; Souillol, V.; Souquet, J.M.; Cheynier, V. Mechanisms of anthocyanin degradation in grape mustlike model solutions. J. Sci. Food Agric 1995, 69, 385–391. [Google Scholar]
- Mazza, G. Anthocyanins in grapes and grape products. Crit. Rev. Food Sci 1995, 35, 341–371. [Google Scholar]
- European Community (CE) Regulation NO. 479/2008 of 29 April 2008 on the common organisation of the market in wine; Official Journal of the European Union: Luxembourg, Luxembourg, 2008.
- De Rosso, M.; Tonidandel, L.; Larcher, R.; Nicolini, G.; Ruggeri, V.; dalla Vedova, A.; de Marchi, F.; Gardiman, M.; Flamini, R. Study of anthocyanic profiles of twenty-one hybrid grape varieties by liquid chromatography and precursor-ion mass spectrometry. Anal. Chim. Acta 2012, 732, 120–129. [Google Scholar]
- Mazzuca, P.; Ferranti, P.; Picariello, G.; Chianese, L.; Addeo, F. Mass spectrometry in the study of anthocyanins and their derivatives: Differentiation of Vitis vinifera and hybrid grapes by liquid chromatography/electrospray ionization mass spectrometry and tandem mass spectrometry. J. Mass Spectrom 2005, 40, 83–90. [Google Scholar]
- Gatto, P.; Vrhovsek, U.; Muth, J.; Segala, C.; Romualdi, C.; Fontana, P.; Pruefer, D.; Stefanini, M.; Moser, C.; Mattivi, F.; et al. Ripening and genotype control stilbene accumulation in healthy grapes. J. Agric. Food Chem 2008, 56, 11773–11785. [Google Scholar]
- Bavaresco, L.; Fregoni, M.; Trevisan, M.; Mattivi, F.; Vrhovsek, U.; Falchetti, R. The occurrence of the stilbene piceatannol in grapes. Vitis 2002, 41, 133–136. [Google Scholar]
- Vitrac, X.; Bornet, A.L.; Vanderlinde, R.; Valls, J.; Richard, T.; Delaunay, J.C.; Mérillon, J.M.; Teissédre, P.L. Determination of stilbenes (δ-viniferin, trans-astringin, trans-piceid, cis- and trans-resveratrol, ɛ-viniferin) in brazilian wines. J. Agric. Food Chem 2005, 53, 5664–5669. [Google Scholar]
- Castellarin, S.D.; Bavaresco, L.; Falginella, L.; Gonçalves, M.I.V.Z.; di Gaspero, G. Phenolics in Grape Berry and Key Antioxidants. In The Biochemistry of the Grape Berry; Gerós, H., Chaves, M.M., Delrot, S., Eds.; Bentham Science Publishers: Bussum, The Netherlands, 2012; pp. 90–91. [Google Scholar]
- Hung, L.M.; Chen, J.K.; Huang, S.S.; Lee, R.S.; Su, M.J. Cardioprotective effect of resveratrol, a natural antioxidant derived from grapes. Cardiovasc. Res 2000, 47, 549–555. [Google Scholar]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar]
- Frankel, E.N.; Waterhouse, A.L.; Kinsella, J.E. Inhibition of human LDL oxidation by resveratrol. Lancet 1993, 341, 1103–1104. [Google Scholar]
- Frémont, L.; Belguendouz, L.; Delpal, S. Antioxidant activity of resveratrol and alcohol-free wine polyphenols related to LDL oxidation and polyunsaturated fatty acids. Life Sci 1999, 64, 2511–2521. [Google Scholar]
- Bertelli, A.A.; Giovannini, L.; Giannessi, D.; Migliori, M.; Bernini, W.; Fregoni, M.; Bertelli, A. Antiplatelet activity of synthetic and natural resveratrol in red wine. Int. J. Tissue React 1995, 17, 1–3. [Google Scholar]
- Pace-Asciak, C.R.; Hahn, S.E.; Diamandis, E.P.; Soleas, G.; Goldberg, D.M. The red wine phenolics trans-resveratrol and quercetin block human platelet aggregation and eicosanoid synthesis: Implications for protection against coronary heart disease. Clin. Chim. Acta 1995, 235, 207–219. [Google Scholar]
- Bavaresco, L.; Mattivi, F.; de Rosso, M.; Flamini, R. Effects of elicitors, viticultural factors, and enological practices on resveratrol and stilbenes in grapevine and wine. Mini Rev. Med. Chem 2012, 12, 1366–1381. [Google Scholar]
- Geahlen, R.L.; McLaughlin, J.L. Piceatannol (3,4,3′,5′-tetrahydroxy-trans-stilbene) is a naturally occurring protein-tyrosine kinase inhibitor. Biochem. Bioph. Res. Commun 1989, 165, 241–245. [Google Scholar]
- Swanson-Mungerson, M.; Ikeda, M.; Lev, L.; Longnecker, R.; Portis, T. Identification of latent membrane protein 2A (LMP2A) specific targets for treatment and eradication of Epstein-Barr virus (EBV)-associated diseases. J. Antimicrob. Chemother 2003, 52, 152–154. [Google Scholar]
- Larrosa, M.; Tomás-Barberán, F.A.; Espín, J.-C. The grape and wine polyphenol piceatannol is a potent inducer of apoptosis in human SK-Mel-28 melanoma cells. Eur. J. Nutr 2004, 43, 275–284. [Google Scholar]
- Bavaresco, L.; Petegolli, D.; Cantu, E.; Fregoni, M.; Chiusa, G.; Trevisan, M. Elicitation and accumulation of stilbene phytoalexins in grapevine berries infected by Botrytis cinerea. Vitis 1997, 36, 77–83. [Google Scholar]
- Pezet, R.; Gindro, K.; Viret, O.; Spring, J.L. Glycosylation and oxidative dimerization of resveratrol are respectively associated to sensitivity and resistance of grapevine cultivars to downy mildew. Physiol. Mol. Plant Pathol 2004, 65, 297–303. [Google Scholar]
- Mattivi, F.; Vrhovsek, U.; Malacarne, G.; Masuero, D.; Zulini, L.; Stefanini, M.; Moser, C.; Velasco, R.; Guella, G. Profiling of resveratrol oligomers, important stress metabolites accumulating in the leaves of hybrid V. vinifera (Merzling × Teroldego) genotypes infected with Plasmopara viticola. J. Agric. Food Chem 2011, 59, 5364–5375. [Google Scholar]
- Pawlus, A.D.; Sahli, R.; Bisson, J.; Rivière, C.; Delaunay, J.-C.; Richard, T.; Gomès, E.; Bordenave, L.; Waffo-Téguo, P.; Mérillon, J.-M. Stilbenoid profiles of canes from Vitis and Muscadinia Species. J. Agric. Food Chem 2013, 61, 501–511. [Google Scholar]
- Cichewicz, R.H.; Kouzi, S.A.; Hamann, M.T. Dimerization of resveratrol by the grapevine pathogen Botrytis cinerea. J. Nat. Prod 2000, 63, 29–33. [Google Scholar]
- Sbaghi, M.; Jeandet, P.; Bessis, R.; Leroux, P. Degradation of stilbene-type phytoalexins in relation to the pathogenicity of Botrytis cinerea to grapevines. Plant Pathol 1996, 45, 139–144. [Google Scholar]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar]
- Koyama, K.; Ikeda, H.; Poudel, P.R.; Goto-Yamamoto, N. Light quality affects flavonoid biosynthesis in young berries of Cabernet Sauvignon grape. Phytochemistry 2012, 78, 54–64. [Google Scholar]
- Makris, D.P.; Kallithraka, S.; Kefalas, P. Flavonols in grapes, grape products and wines: Burden, profile and influential parameters. J. Food Compos. Anal 2006, 19, 396–404. [Google Scholar]
- Castillo-Muñoz, N.; Gómez-Alonso, S.; García-Romero, E.; Gómez, M.V.; Velders, A.H.; Hermosín-Gutiérrez, I. Flavonol 3-O-glycosides series of Vitis vinifera Cv. Petit Verdot Red Wine Grapes. J. Agric. Food Chem 2009, 57, 209–219. [Google Scholar]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. Synthesis of flavonols and expression of flavonol synthase genes in the developing grape berries of Shiraz and Chardonnay (Vitis vinifera L.). Aust. J. Grape Wine Res 2003, 9, 110–121. [Google Scholar]
- Hermosín-Gutiérrez, I.; Castillo-Muñoz, N.; Gómez-Alonso, S.; García-Romero, E. Flavonol Profiles for Grape and Wine Authentication. In Progress in Authentication of Food and Wine; Ebeler, S.E., Takeoka, G.R., Winterhalter, P., Eds.; American Chemical Society: Washington, DC, USA, 2012; pp. 113–129. [Google Scholar]
- Liang, N.-N.; He, F.; Bi, H.-Q.; Duan, C.-Q.; Reeves, M.J.; Wang, J. Evolution of flavonols in berry skins of different grape cultivars during ripening and a comparison of two vintages. Eur. Food Res. Technol 2012, 235, 1187–1197. [Google Scholar]
- Liang, Z.; Yang, Y.; Cheng, L.; Zhong, G.-Y. Polyphenolic composition and content in the ripe berries of wild Vitis species. Food Chem 2012, 132, 730–738. [Google Scholar]
- Castillo-Muñoz, N.; Gómez-Alonso, S.; García-Romero, E.; Hermosín-Gutiérrez, I. Flavonol profiles of Vitis vinifera white grape cultivars. J. Food Compos. Anal 2010, 23, 699–705. [Google Scholar]
- Perestrelo, R.; Lu, Y.; Santos, S.A.O.; Silvestre, A.J.D.; Neto, C.P.; Câmara, J.S.; Rocha, S.M. Phenolic profile of Sercial and Tinta Negra Vitis vinifera L. grape skins by HPLC–DAD–ESI-MSn: Novel phenolic compounds in Vitis vinifera L. grape. Food Chem 2012, 135, 94–104. [Google Scholar]
- Liang, Z.; Owens, C.L.; Zhong, G.-Y.; Cheng, L. Polyphenolic profiles detected in the ripe berries of Vitis vinifera germplasm. Food Chem 2011, 129, 940–950. [Google Scholar]
- McDonald, M.S.; Hughes, M.; Burns, J.; Lean, M.E.J.; Matthews, D.; Crozier, A. Survey of the free and conjugated myricetin and quercetin content of red wines of different geographical origins. J. Agric. Food Chem 1998, 46, 368–375. [Google Scholar]
- Mori, K.; Sugaya, S.; Gemma, H. Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci. Hortic. Amst 2005, 105, 319–330. [Google Scholar]
- Yamane, T.; Jeong, S.T.; Goto-Yamamoto, N.; Koshita, Y.; Kobayashi, S. Effects of temperature on anthocyanin biosynthesis in grape berry skins. Am. J. Enol. Vitic 2006, 57, 54–59. [Google Scholar]
- Kliewer, W.M.; Torres, R.E. Effect of controlled day and night temperatures on grape coloration. Am. J. Enol. Vitic 1972, 23, 71–77. [Google Scholar]
- Figueiredo-González, M.; Cancho-Grande, B.; Boso, S.; Santiago, J.L.; Martínez, M.C.; Simal-Gándara, J. Evolution of flavonoids in Mouratón berries taken from both bunch halves. Food Chem 2013, 138, 1868–1877. [Google Scholar]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. The effect of bunch shading on berry development and flavonoid accumulation in Shiraz grapes. Aust. J. Grape Wine Res 2004, 10, 55–73. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Flamini, R.; Mattivi, F.; Rosso, M.D.; Arapitsas, P.; Bavaresco, L. Advanced Knowledge of Three Important Classes of Grape Phenolics: Anthocyanins, Stilbenes and Flavonols. Int. J. Mol. Sci. 2013, 14, 19651-19669. https://doi.org/10.3390/ijms141019651
Flamini R, Mattivi F, Rosso MD, Arapitsas P, Bavaresco L. Advanced Knowledge of Three Important Classes of Grape Phenolics: Anthocyanins, Stilbenes and Flavonols. International Journal of Molecular Sciences. 2013; 14(10):19651-19669. https://doi.org/10.3390/ijms141019651
Chicago/Turabian StyleFlamini, Riccardo, Fulvio Mattivi, Mirko De Rosso, Panagiotis Arapitsas, and Luigi Bavaresco. 2013. "Advanced Knowledge of Three Important Classes of Grape Phenolics: Anthocyanins, Stilbenes and Flavonols" International Journal of Molecular Sciences 14, no. 10: 19651-19669. https://doi.org/10.3390/ijms141019651
APA StyleFlamini, R., Mattivi, F., Rosso, M. D., Arapitsas, P., & Bavaresco, L. (2013). Advanced Knowledge of Three Important Classes of Grape Phenolics: Anthocyanins, Stilbenes and Flavonols. International Journal of Molecular Sciences, 14(10), 19651-19669. https://doi.org/10.3390/ijms141019651