
Insulin-Like Growth Factor-2 Is Induced Following 5-Aminolevulinic Acid-Mediated Photodynamic Therapy in SW620 Human Colon Cancer Cell Line

Abstract
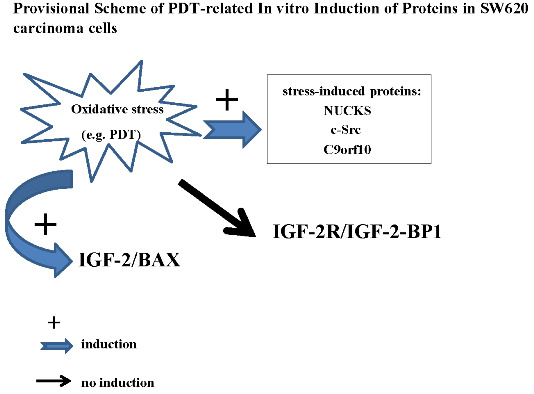
:
1. Introduction
2. Results and Discussion
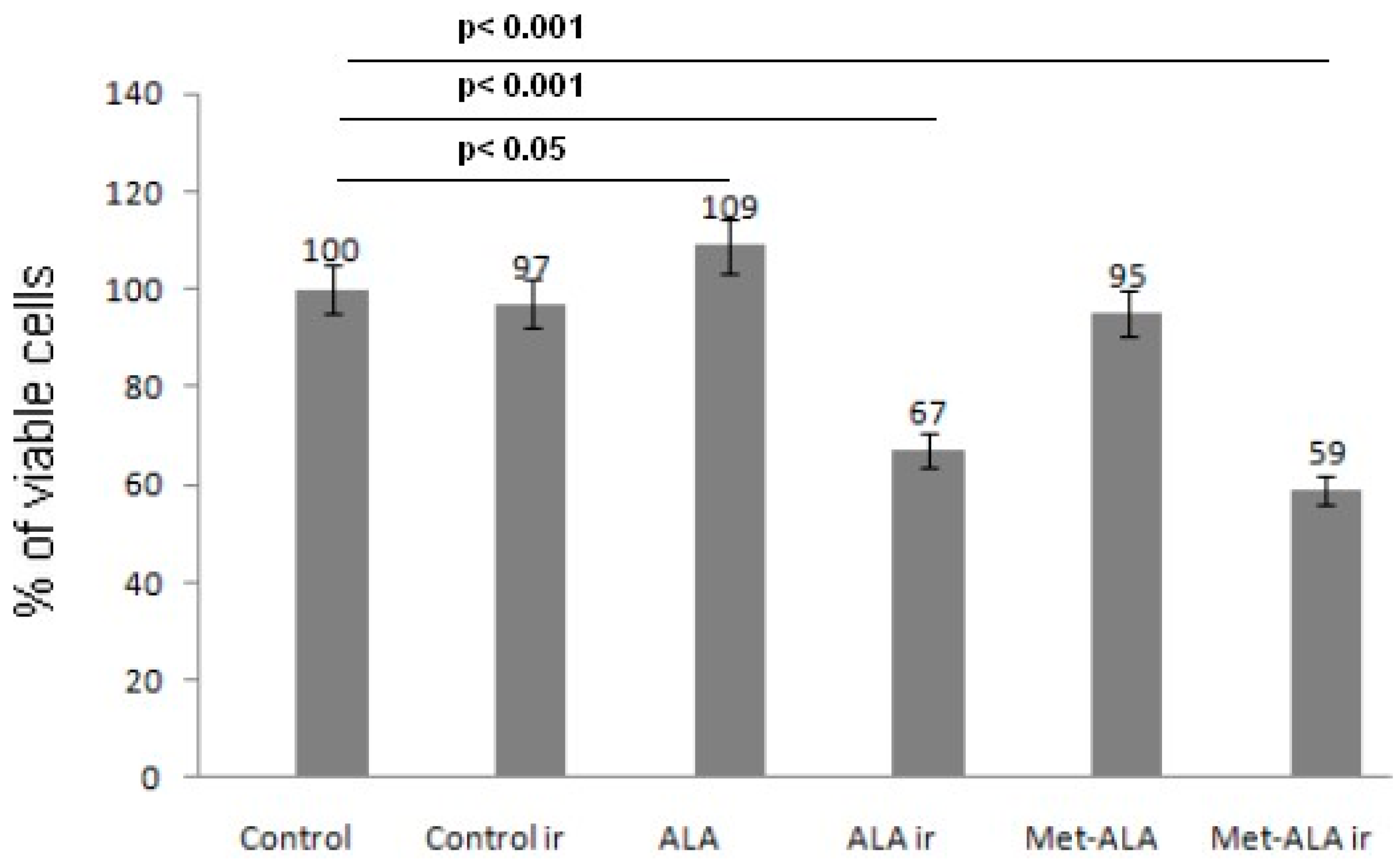
2.1. Cell Proliferation MTT Assay

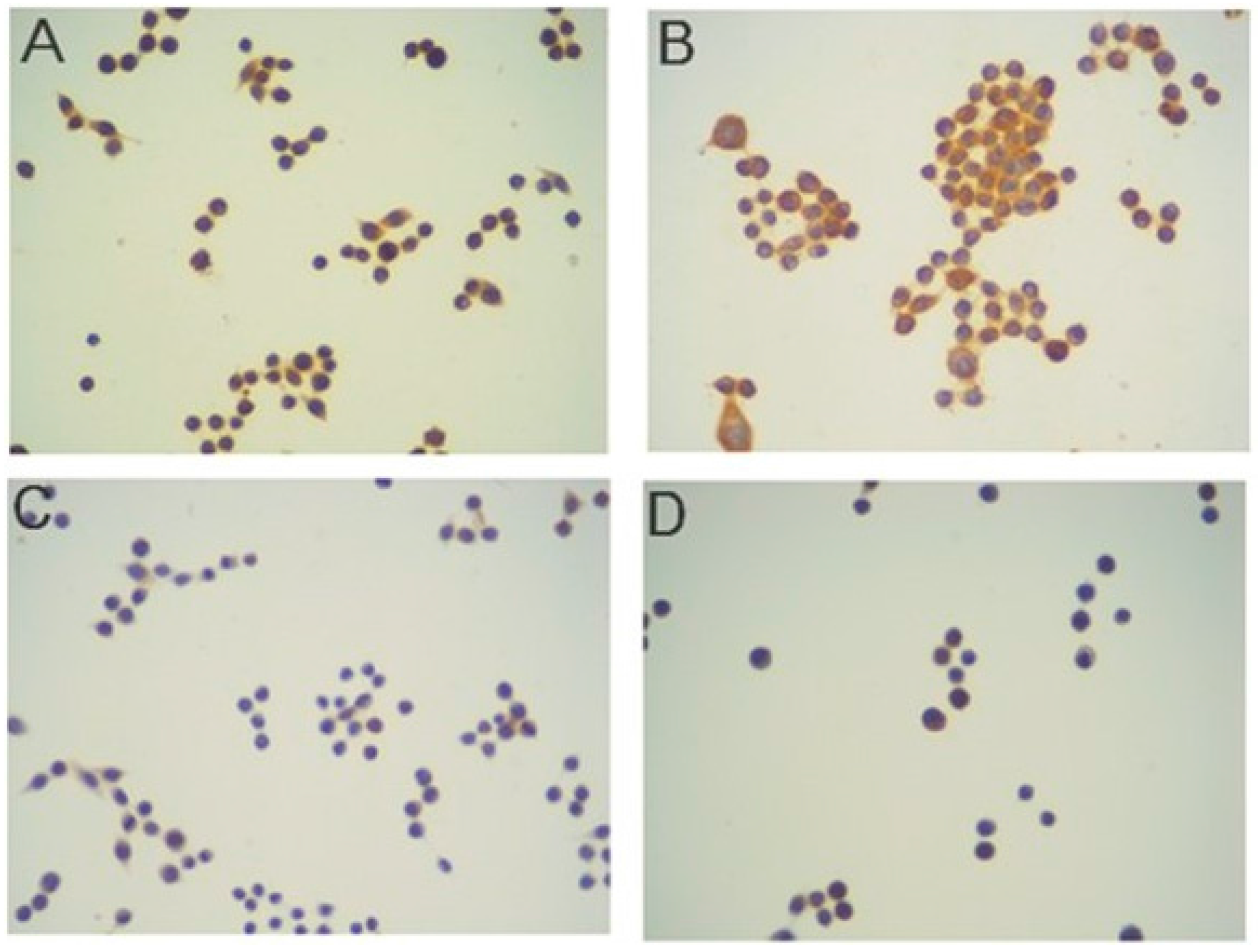
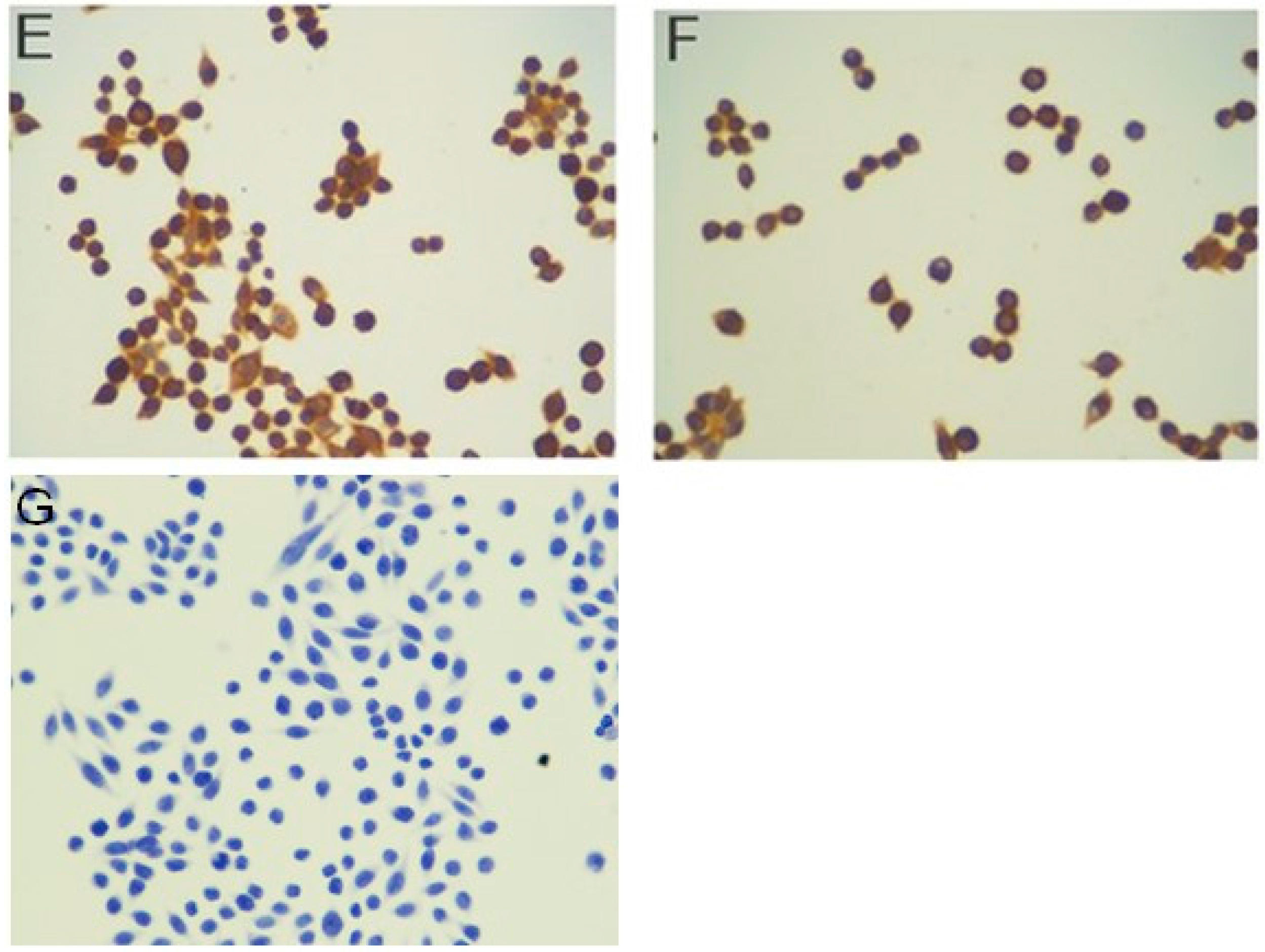
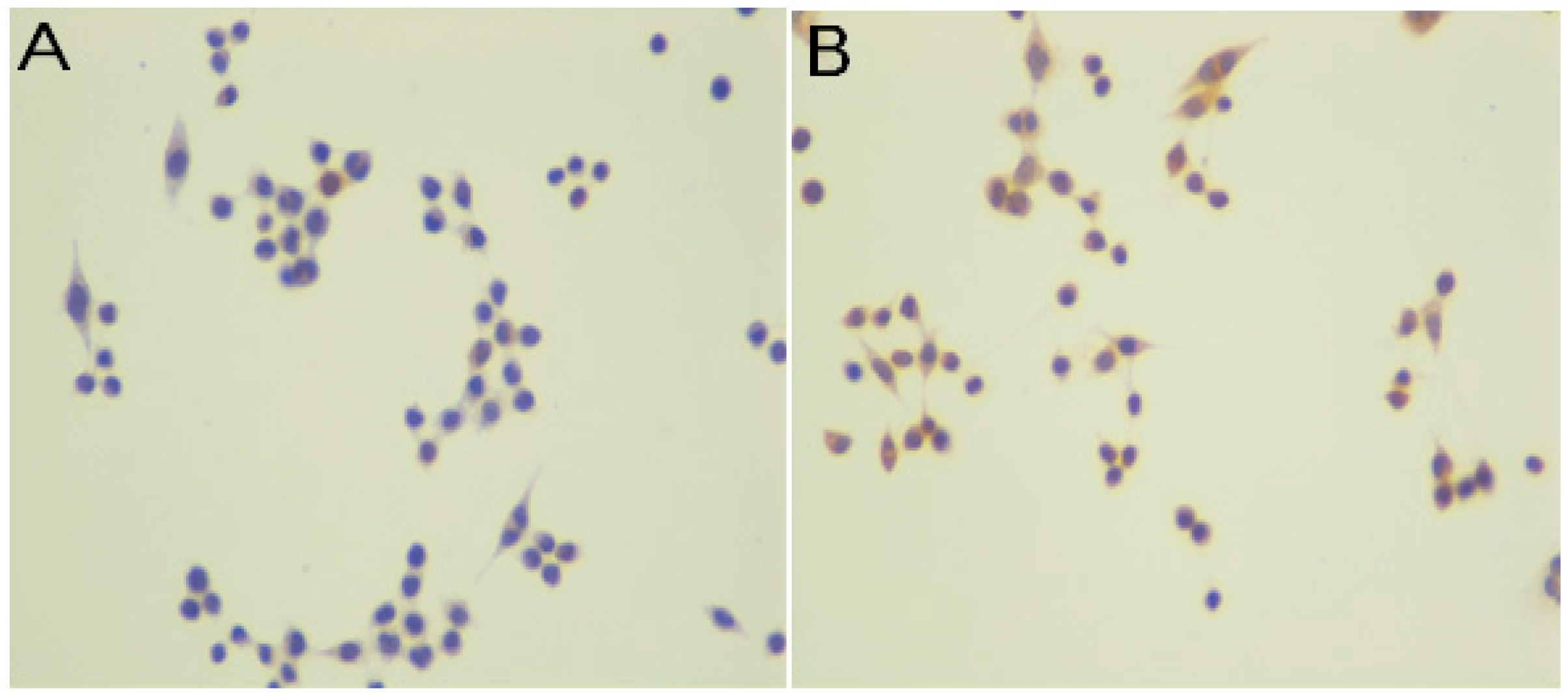
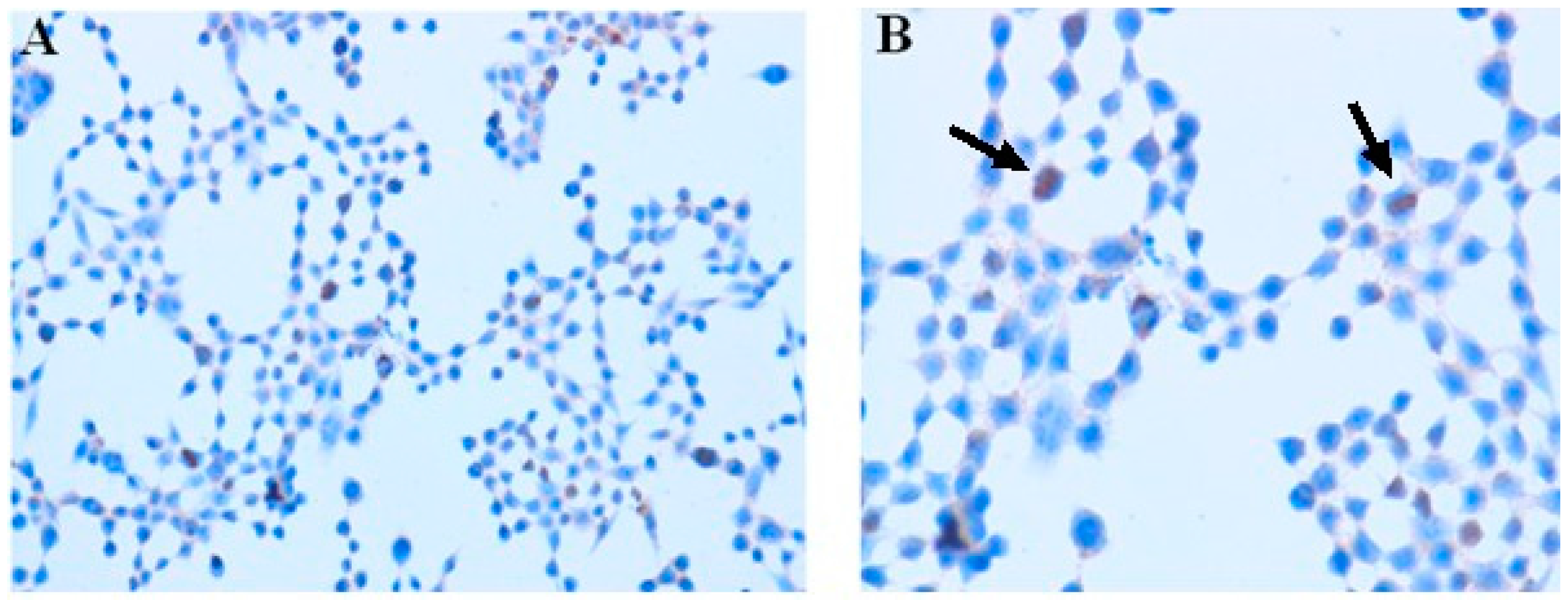
2.2. Immunocytochemistry



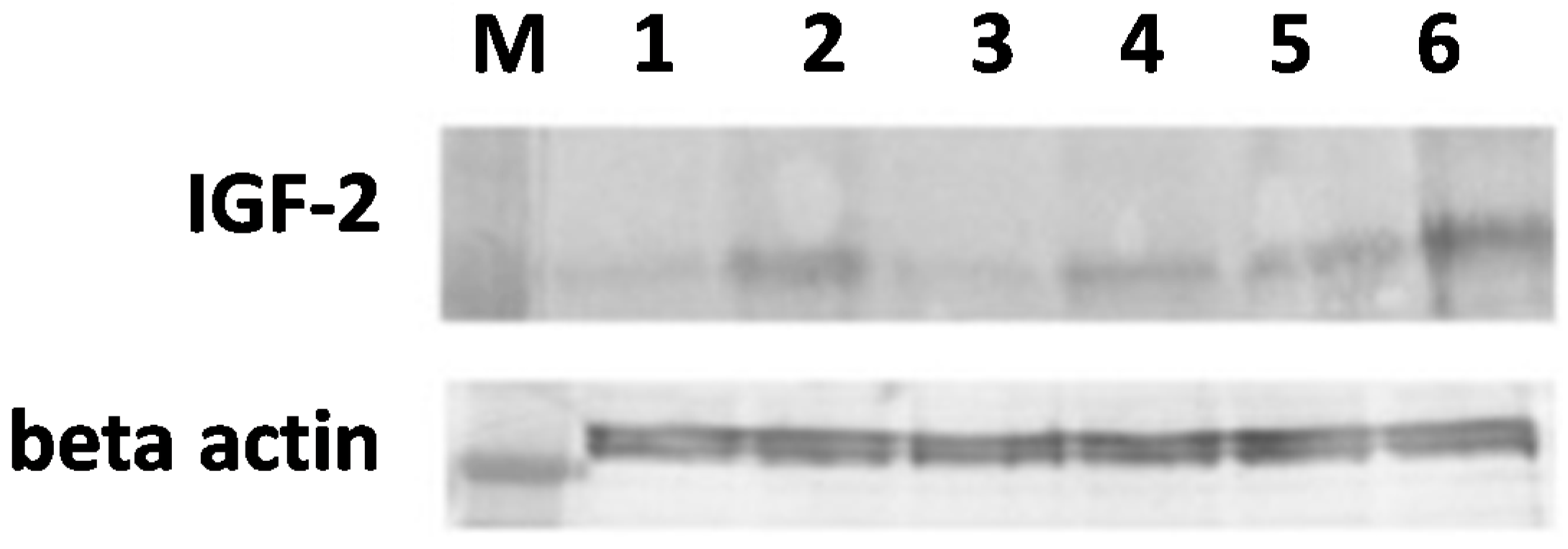
2.3. Western Blot Analysis

2.4. Apoptosis Detection

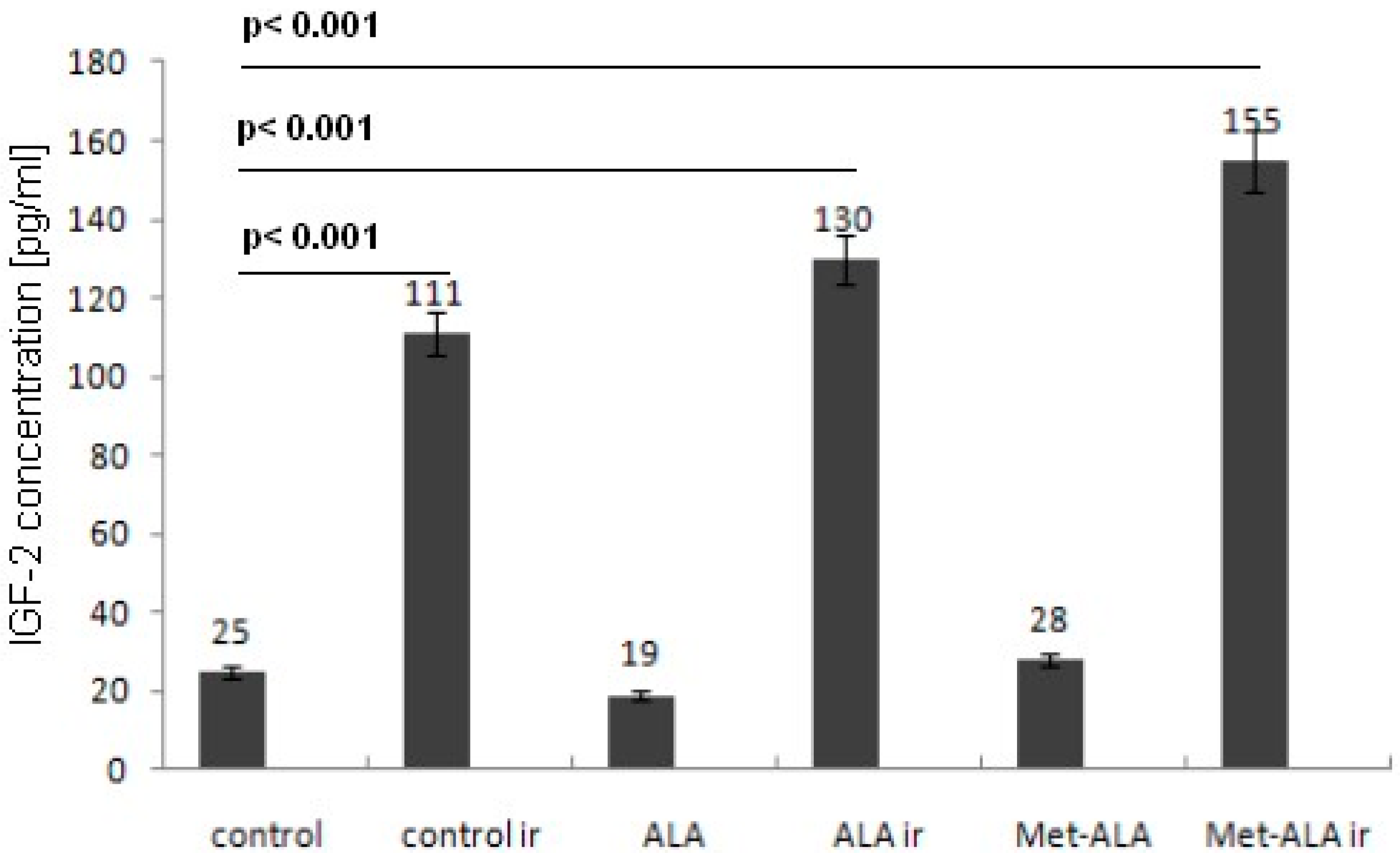
2.5. ELISA Test

2.6. Discussion
3. Experimental Section
3.1. Precursors of Photosensitizer
3.2. Cell Line
3.3. Light Source
3.4. Cell Proliferation MTT Assay
3.5. Cell Culture Conditions
3.6. Cells Culture and Photosensitization Conditions in Experiments for Immunocytochemical Studies
3.7. Immunocytochemistry

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Intensity of Staining | Number of Stained Cells (Quantity) |
|---|---|
| 0, negative staining | 0, <5% |
| 1, weak | 1, 5% to 25% |
| 2, moderate | 2, 25% to 75% |
| 3, strong | 3, >75% |
3.8. Cell Culture and Photosensitization Conditions in Experiments for Western Blot Analysis
3.9. Western Blotting
3.10. IGF-2 ELISA
3.11. Apoptosis Detection
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.C.; Braghiroli, M.I.; Sabbaga, J.; Hoff, P.M. Primary prevention of colorectal cancer: Myth or reality? World J. Gastroenterol. 2014, 20, 15060–15069. [Google Scholar] [CrossRef] [PubMed]
- Mishra, J.; Dromund, J.; Quazi, S.H.; Karanki, S.S.; Shaw, J.J.; Chen, Ben.; Kumar, N. Prospective of Colon Cancer Treatments and Scope for Combinatorial Approach to Enhanced Cancer Cell Apoptosis. Crit. Rev. Oncol. Hematol. 2013, 86, 232–250. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Wilson, C.; Hasan, T.; Maytin, E.V. Vitamin D3 enhances the apoptotic response of epithelial tumors to aminolevulinate-based photodynamictherapy. Cancer Res. 2011, 71, 6040–6050. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, C. IGF-2 and cancer. Endocr. Relat. Cancer 2013, 20, 321–339. [Google Scholar] [CrossRef] [PubMed]
- Bach, L.; Fu, P.; Yang, Z. Insulin-like growth factor-binding protein-6 and cancer. Clin. Sci. 2013, 124, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Lynda, K.; Harris, I.P.; Crocker, P.N.; Baker, J.D.; Westwood, M. IGF2 actions on trophoblast in human placenta are regulated by the insulin-like growth factor 2 receptor, which can function as both a signaling and clearance receptor. Biol. Reprod. 2011, 84, 440–446. [Google Scholar]
- Cianfarani, S. Insulin-likegrowthfactor-II: New roles for an old actor. Front. Endocrinol. 2012, 3, 118. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.K.; Westwood, M. Biology and significance of signalling pathways activated by IGF-II. Growth Factors 2012, 30, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Firth, S.M.; Baxter, R.C. Cellular Actions of the Insulin-Like Growth Factor Binding Proteins. Endocr. Rev. 2002, 23, 824–854. [Google Scholar] [CrossRef] [PubMed]
- Breuhah, K.; Longerich, T.; Schirmacher, P. Dysregulation of growth factor signaling in human hepatocellular carcinoma. Oncogene 2006, 25, 3787–3800. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Han, W.; Tergaonkar, V. NUCKS: A potential biomarker in cancer and metabolic disease. Clin. Sci. 2015, 128, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Ziółkowski, P.; Gamian, E.; Osiecka, B.; Zougman, A.; Wiśniewski, J.R. Immunohistochemical and proteomic evaluation of nuclear ubiquitous casein and cyclin-dependent kinases substrate in invasive ductal carcinoma of the breast. J. Biomed. Biotechnol. 2009, 2009, 919645. [Google Scholar] [CrossRef] [PubMed]
- Hotowy, K.; Woźniak, M.; Duś, K.; Czapińska, E.; Osiecka, B.; Krzystek-Korpacka, M.; Bronowicz, A.; Wiśniewski, J.; Gamian, A.; Terlecki, G.; et al. Immunocytochemical studies on the nuclear ubiquitous casein and cyclin-dependent kinases substrate following 5-aminolevulinic acid-mediated photodynamic therapy on MCF-7 cells. Photodiagn. Photodyn. Ther. 2013, 10, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, M.; Hotowy, K.; Czapińska, E.; Duś-Szachniewicz, K.; Szczuka, I.; Gamian, E.; Gamian, A.; Terlecki, G.; Ziółkowski, P. Early induction of stress-associated Src activator/Homo sapiens chromosome 9 open reading frame 10 protein following photodynamic therapy. Photodiagn. Photodyn. Ther. 2014, 11, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.F.; Wang, S.; Thakar, M.; Smolinski, K.N.; Yin, J.; Zou, T.T.; Kong, D.; Abraham, J.M.; Toretsky, J.A.; Meltzer, S.J. Expression of the wild-type insulin-like growth factor II receptor gene suppresses growth and causes death in colorectal carcinoma cells. Oncogene 1999, 18, 4063–4068. [Google Scholar] [CrossRef] [PubMed]
- Werner, H.; LeRoith, D. The insulin-like growth factor-I receptor signaling pathways are important for tumorigenesis and inhibition of apoptosis. Crit. Rev. Oncogene 1997, 8, 71–92. [Google Scholar] [CrossRef]
- Srivastava, M.; Ahmad, N.; Gupta, S.; Mukhtar, H. Involvement of Bcl-2 and Bax in photodynamic therapy-mediated apoptosis. Antisense Bcl-2 oligonucleotide sensitizes RIF 1 cells to photodynamic therapy apoptosis. J. Biol. Chem. 2001, 276, 15481–15488. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Luo, Y.; Li, G.; Kessel, D. Enhanced apoptotic response to photodynamic therapy after Bcl-2 transfection. Cancer Res. 1999, 59, 3429–3432. [Google Scholar] [PubMed]
- Chan, J.; Stampfer, M.; Giovannucci, E.; Gann, P.; Ma, J.; Wilkinson, P.; Hennekens, C.H.; Pollak, M. Plasma insulin-like growth factor 1 and prostate cancer risk: A prospective study. Science 1998, 279, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Zaina, S.; Squire, S. The soluble type 2 insulin-like growth (IGF-II) receptor reduces organ size by IGF-II-mediated and IGF-II-independent mechanisms. J. Biol. Chem. 1998, 273, 28610–28616. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, D.B.; Weiss, J.; Hettiaratchi, A.; Firth, S.M.; Scott, C.D. Insulin-like growth factor-II/mannose 6-phosphate receptor overexpression reduces growth of choriocarcinoma cells in vitro and in vivo. Endocrinology 2002, 143, 4287–4294. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, H.; Shiwaku, H.O.; Hagiwara, H.; Miura, K.; Abe, T.; Kato, Y.; Ohtani, H.; Shiiba, K.; Souza, R.F.; Meltzer, S.J.; et al. The insulin-like growth factor II receptor gene is mutated in genetically unstable cancers of the endometrium, stomach, and colorectum. Cancer Res. 1997, 57, 1851–1854. [Google Scholar] [PubMed]
- Hebert, E. Mannose-6-phosphate/insulin-like growth factor II receptor expression and tumor development. Biosci. Rep. 2006, 26, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ge, Y.; Landman, N.; Kang, J.X. Decreased expression of the mannose 6-phosphate/insulin-like growth factor-2 receptor promotes growth of human breast cancer cells. BMC Cancer 2002, 30, 2–18. [Google Scholar]
- Marshman, E.; Streuli, C.H. Insulin-like growth factors and insulin-like growth factor binding proteins in mammary gland function. Breast Cancer Res. 2002, 4, 231–239. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woźniak, M.; Duś-Szachniewicz, K.; Ziółkowski, P. Insulin-Like Growth Factor-2 Is Induced Following 5-Aminolevulinic Acid-Mediated Photodynamic Therapy in SW620 Human Colon Cancer Cell Line. Int. J. Mol. Sci. 2015, 16, 23615-23629. https://doi.org/10.3390/ijms161023615
Woźniak M, Duś-Szachniewicz K, Ziółkowski P. Insulin-Like Growth Factor-2 Is Induced Following 5-Aminolevulinic Acid-Mediated Photodynamic Therapy in SW620 Human Colon Cancer Cell Line. International Journal of Molecular Sciences. 2015; 16(10):23615-23629. https://doi.org/10.3390/ijms161023615
Chicago/Turabian StyleWoźniak, Marta, Kamila Duś-Szachniewicz, and Piotr Ziółkowski. 2015. "Insulin-Like Growth Factor-2 Is Induced Following 5-Aminolevulinic Acid-Mediated Photodynamic Therapy in SW620 Human Colon Cancer Cell Line" International Journal of Molecular Sciences 16, no. 10: 23615-23629. https://doi.org/10.3390/ijms161023615