
Addition of Interleukin-21 for Expansion of T-Cells for Adoptive Immunotherapy of Murine Melanoma

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
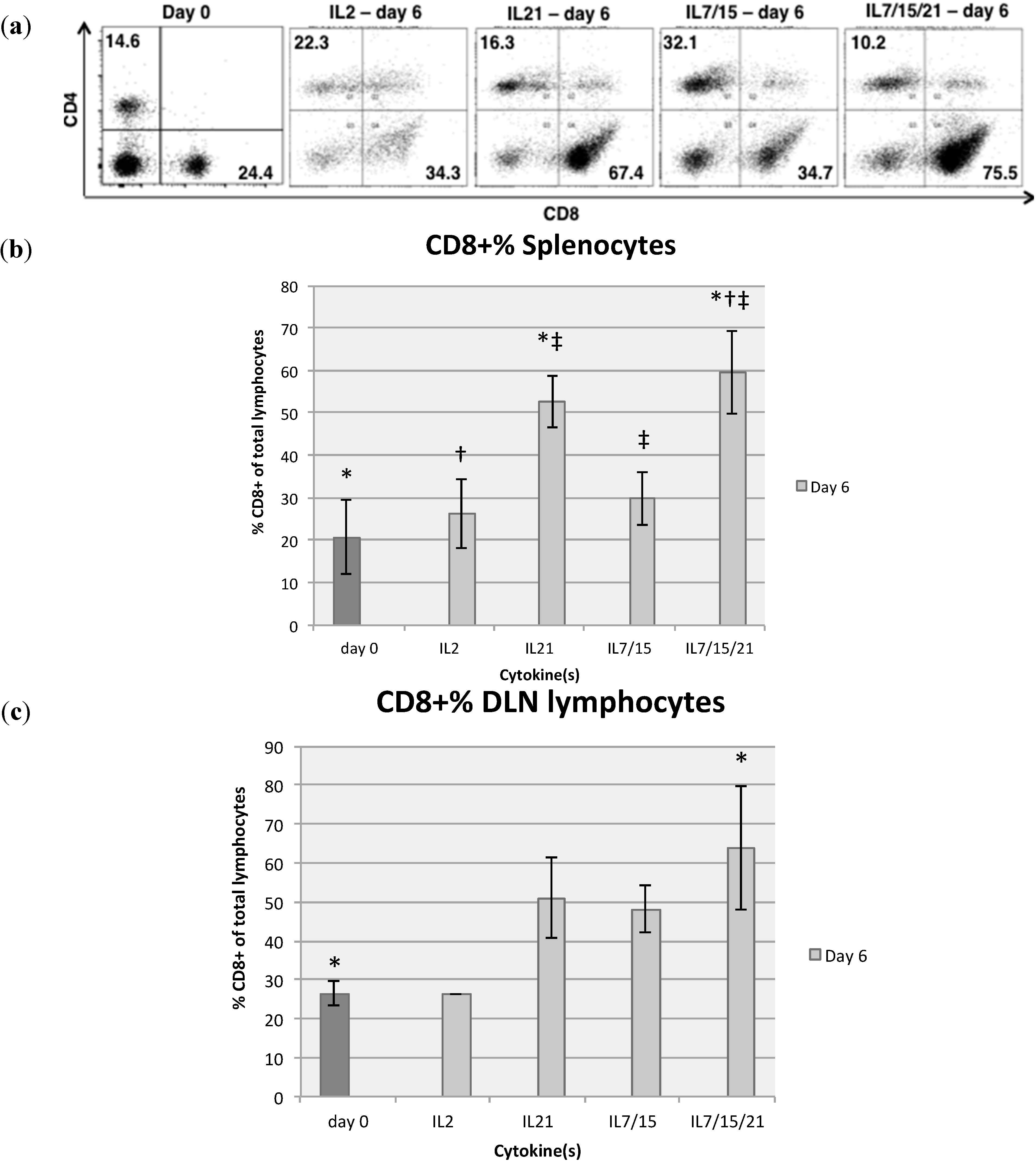
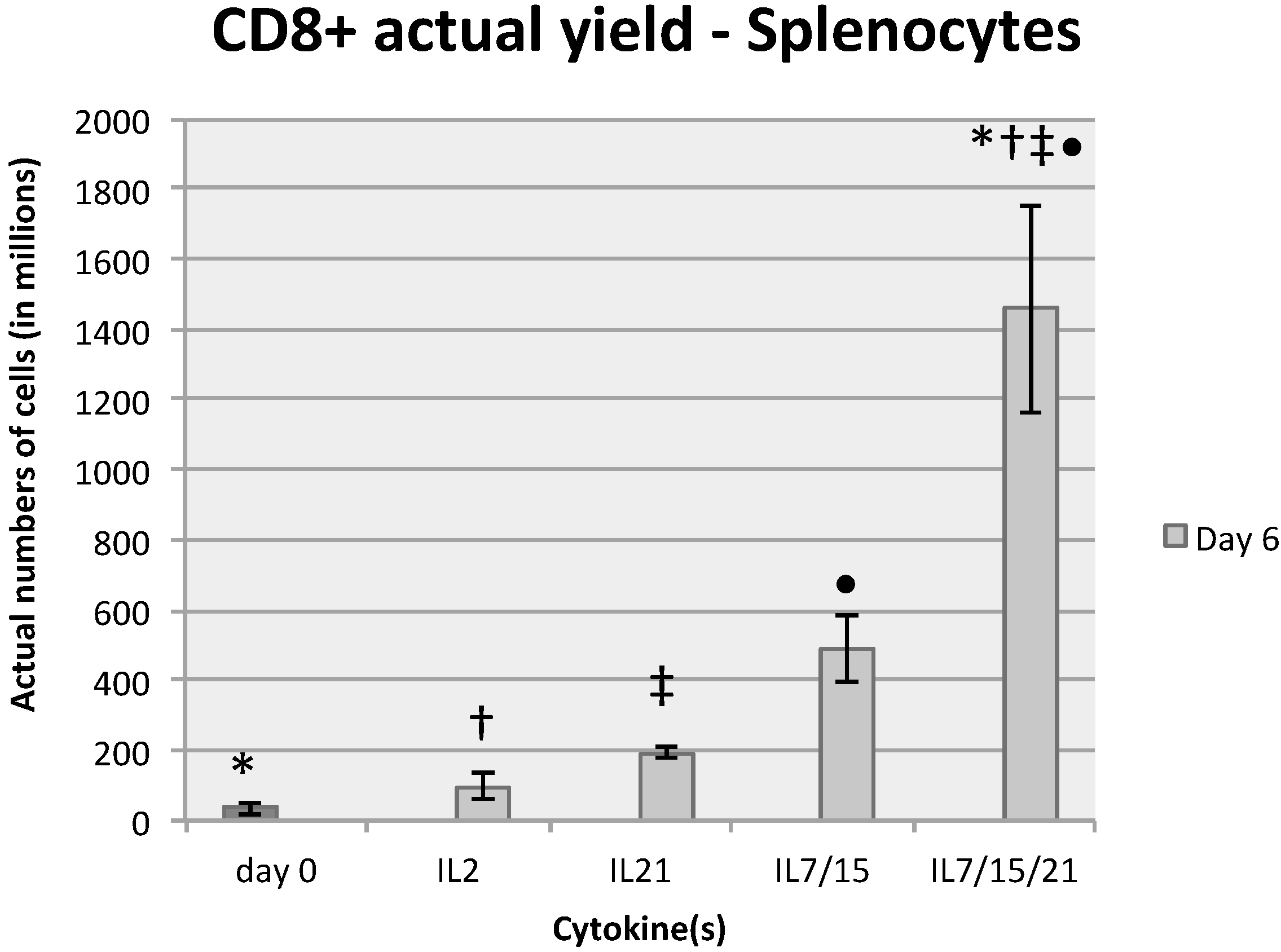
2.1. Comparative Analysis of T Cell Expansion
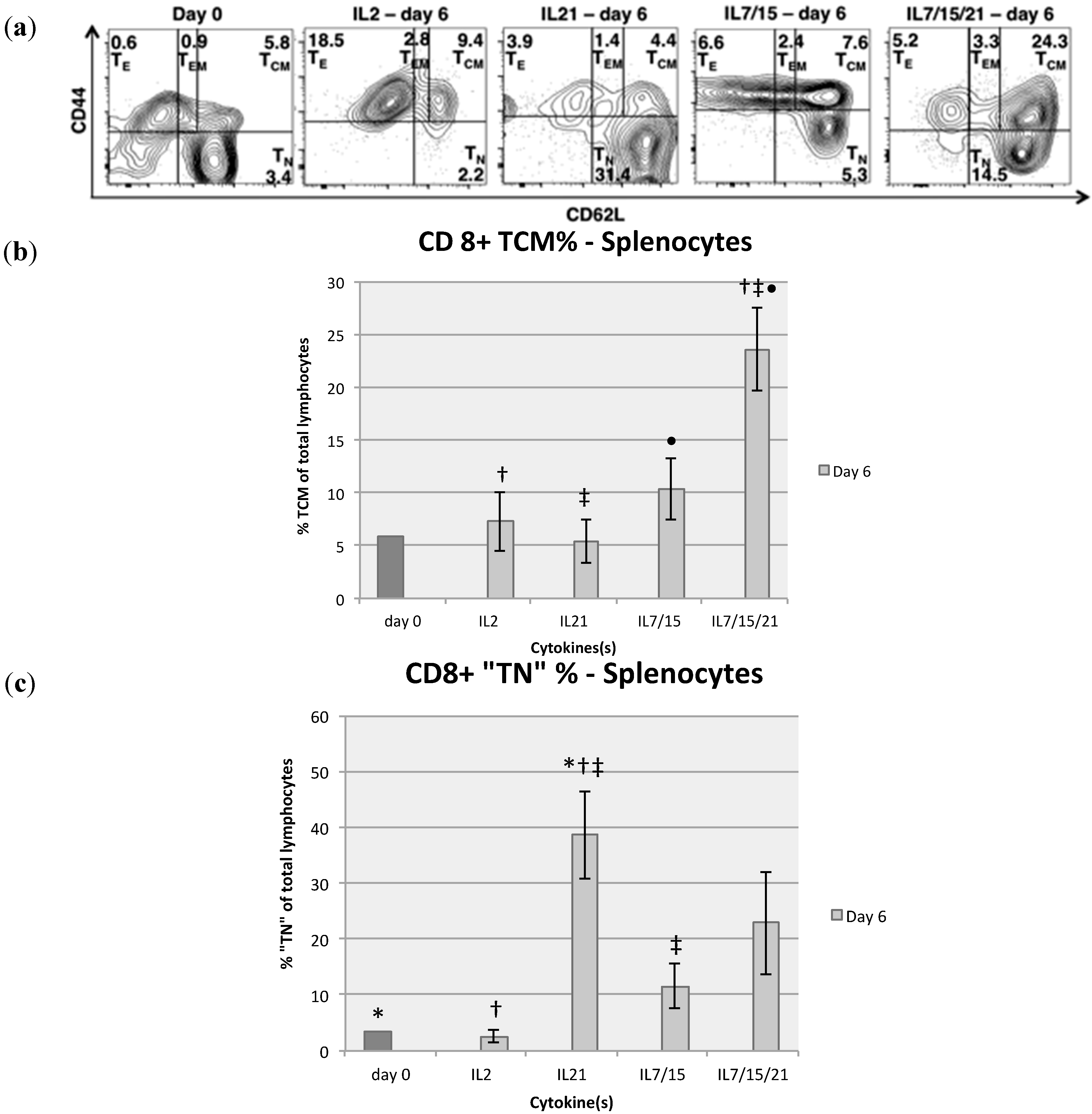
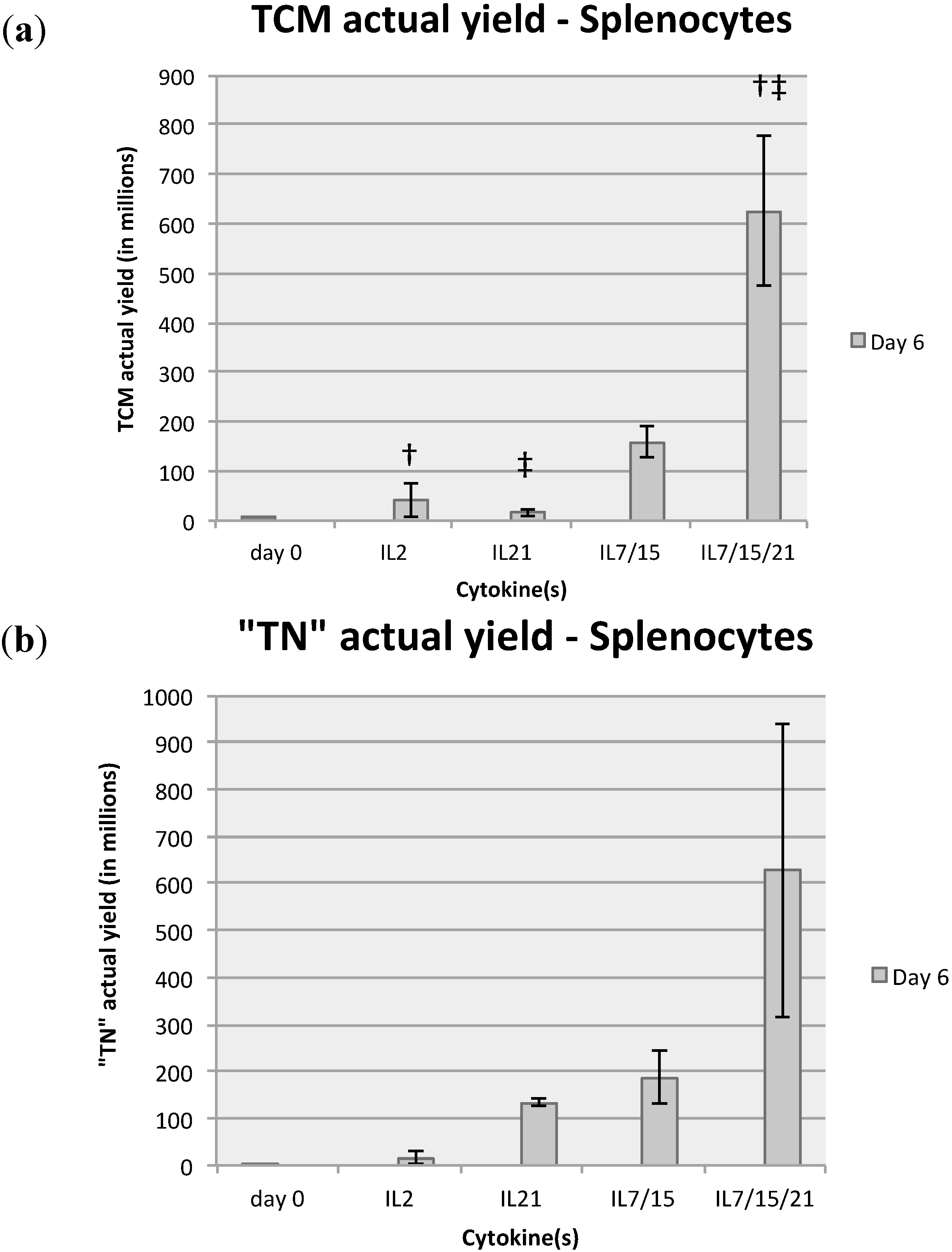
2.2. Comparison of T Cell Phenotype with Various Cytokine Exposure




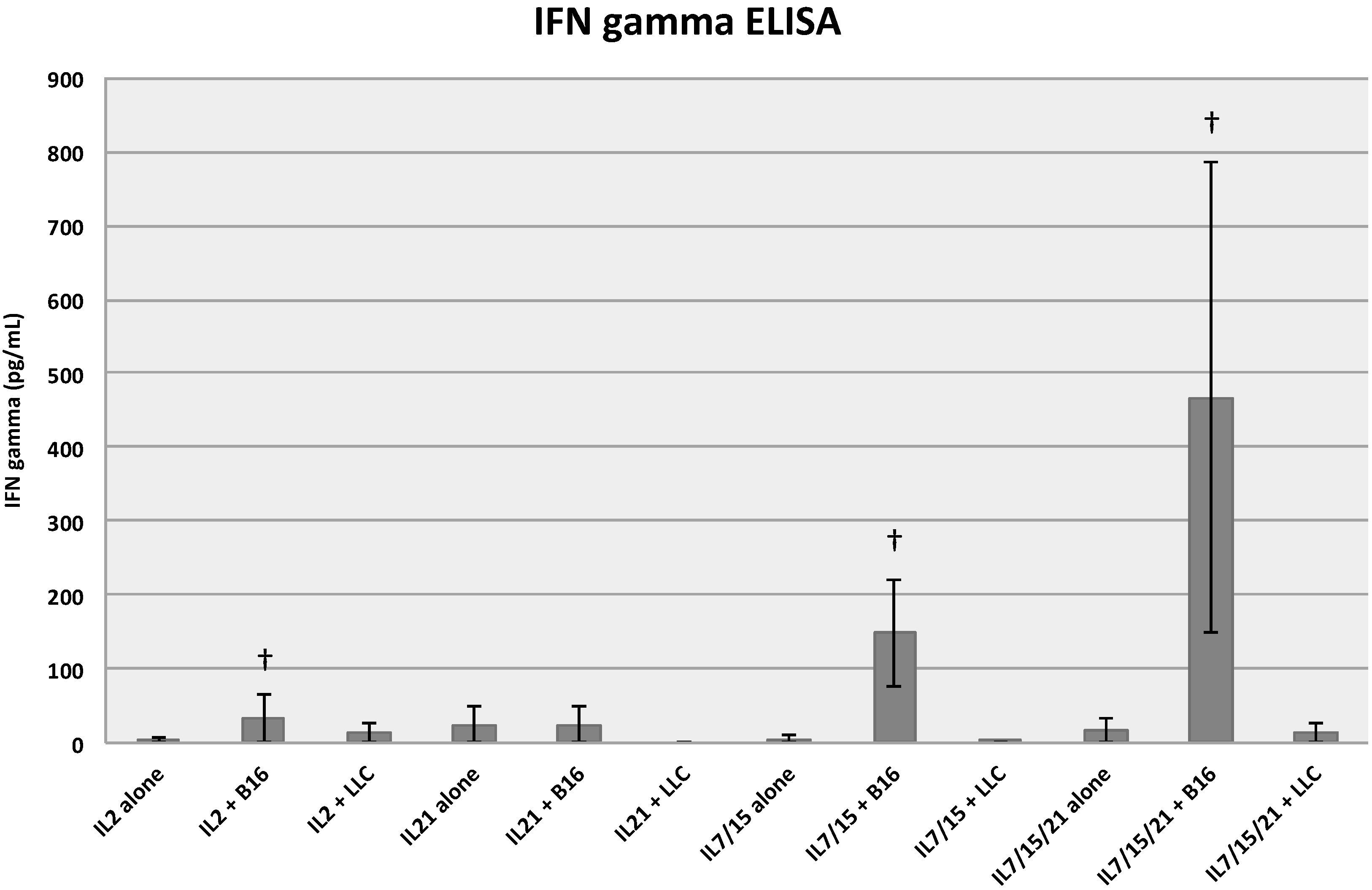
2.3. IFN-Gamma Release ELISA

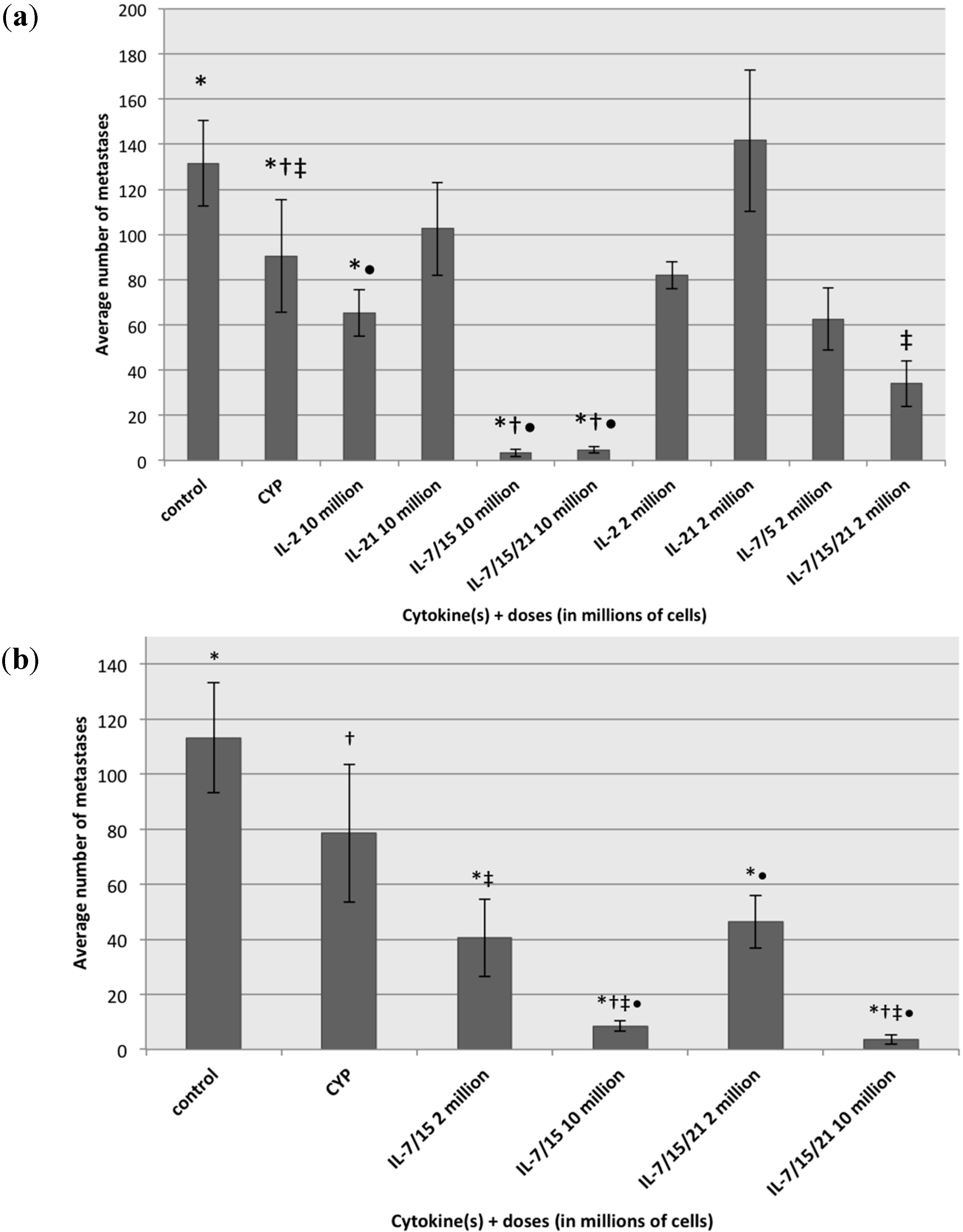
2.4. Treatment of B16 Melanoma Metastases with Lymphocytes Exposed to Different Cytokines

3. Discussion
4. Experimental Section
4.1. Mice
4.2. Tumor Cell Lines
4.3. Draining Lymph Node Sensitization
4.4. Lymphocyte Activation and in Vitro Expansion
4.5. Adoptive Immunotherapy
4.6. Flow Cytometry
4.7. IFN-γ ELISA
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Klebanoff, C.A.; Gattinoni, L.; Palmer, D.C.; Muranski, P.; Ji, Y.; Hinrichs, C.S.; Borman, Z.A.; Kerkar, S.P.; Scott, C.D.; Finkelstein, S.E.; et al. Determinants of successful CD8+ T-cell adoptive immunotherapy for large established tumors in mice. Clin. Cancer Res. 2011, 17, 5343–5352. [Google Scholar] [CrossRef] [PubMed]
- Pouw, N.; Treffers-Westerlaken, E.; Kraan, J.; Wittink, F.; ten Hagen, T.; Verweij, J.; Debets, R. Combination of IL-21 and IL-15 enhances tumour-specific cytotoxicity and cytokine production of TCR-transduced primary T Cells. Cancer Immunol. Immunother. 2010, 59, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Huarte, E.; Fisher, J.; Turk, M.J.; Mellinger, D.; Foster, C.; Wolf, B.; Meehan, K.R.; Fadul, C.E.; Ernstoff, M.S. Ex vivo expansion of tumor specific lymphocytes with IL-15 and IL-21 for adoptive immunotherapy in melanoma. Cancer Lett. 2009, 285, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Bear, H.D.; Roberts, J.; Cornell, D.; Tombes, M.B.; Kyle, B. Adoptive immunotherapy of cancer with pharmacologically activated lymph node lymphocytes: A pilot clinical trial. Cancer Immunol. Immunother. 2001, 50, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Riley, J.; Rosenberg, S.; Parkhurst, M. Comparison of common γ-chain cytokines, interleukin-2, interleukin-7, and interleukin-15 for the in vitro generation of human tumor-reactive T lymphocytes for adoptive cell transfer therapy. J. Immunother. 2006, 29, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Le, H.K.; Graham, L.; Miller, C.H.; Kmieciak, M.; Manjili, M.H.; Bear, H.D. Incubation of antigen-sensitized T lymphocytes activated with bryostatin 1 + ionomycin in IL-7 + IL-15 increases yield of cells capable of inducing regression of melanoma metastases compared to culture in IL-2. Cancer Immunol. Immunother. 2009, 58, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Chatila, T.; Silverman, L.; Miller, R.; Geha, R. Mechanisms of T cell activation by the calcium ionophore ionomycin. J. Immunol. 1989, 143, 1283–1289. [Google Scholar] [PubMed]
- Kazanietz, M.G.; Lewin, N.E.; Gao, F.; Pettit, G.R.; Blumberg, P.M. Binding of [26–3H]bryostatin 1 and analogs to calcium-dependent and calcium-independent protein kinase C isozymes. Mol. Pharmacol. 1994, 46, 374–379. [Google Scholar] [PubMed]
- Pettit, G.R.; Herald, S.L.; Doubek, D.L.; Arnold, E.; Clardy, J. Isolation and structure of bryostatin 1. J. Am. Chem. Soc. 1982, 104, 6846–6848. [Google Scholar] [CrossRef]
- Payne, K.K.; Zoon, C.K.; Wan, W.; Marlar, K.; Keim, R.C.; Kenari, M.N.; Kazim, A.L.; Bear, H.D.; Manjili, M.H. Peripheral blood mononuclear cells of patients with breast cancer can be reprogrammed to enhance anti-HER-2/Neu reactivity and overcome myeloid-derived suppressor cells. Breast Cancer Res. Treat. 2013, 142, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Kmieciak, M.; Basu, D.; Payne, K.K.; Toor, A.; Yacoub, A.; Wang, X.Y.; Smith, L.; Bear, H.D.; Manjili, M.H. Activated NKT cells and NK cells render T cells resistant to myeloid-derived suppressor cells and result in an effective adoptive cellular therapy against breast cancer in the FVBN202 transgenic mouse. J. Immunol. 2011, 187, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Kmieciak, M.; Toor, A.; Graham, L.; Bear, H.D.; Manjili, M.H. Ex vivo expansion of tumor-reactive t cells by means of bryostatin 1/ionomycin and the common γ chain cytokines formulation. J. Vis. Exp. 2011, 47. [Google Scholar] [PubMed]
- Miller, C.H.; Graham, L.; Bear, H.D. Phenotype, functions and fate of adoptively transferred tumor draining lymphocytes activated ex vivo in mice with an aggressive weakly immunogenic mammary carcinoma. BMC Immunol. 2010, 11. [Google Scholar] [CrossRef]
- Cha, E.; Graham, L.; Manjili, M.H.; Bear, H.D. IL-7 + IL-15 are superior to IL-2 for the ex vivo expansion of 4T1 mammary carcinoma-specific T cells with greater efficacy against tumors in vivo. Breast Cancer Res. Treat. 2010, 122, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.S.; Miller, C.H.; Graham, L.; Parviz, M.; Zacur, S.; Patel, B.; Duong, A.; Bear, H.D. Bryostatin 1/Ionomycin (B/I) ex vivo stimulation preferentially activates l-selectinlow tumor-sensitized lymphocytes. Int. Immunol. 2004, 16, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- Parviz, M.; Chin, C.S.; Graham, L.J.; Miller, C.; Lee, C.; George, K.; Bear, H.D. Successful adoptive immunotherapy with vaccine-sensitized T cells, despite no effect with vaccination alone in a weakly immunogenic tumor model. Cancer Immunol. Immunother. 2003, 52, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Klebanoff, C.A.; Gattinoni, L.; Torabi-Parizi, P.; Kerstann, K.; Cardones, A.R.; Finkelstein, S.E.; Palmer, D.C.; Antony, P.A.; Hwang, S.T.; Rosenberg, S.A.; et al. Central memory self/tumor-reactive CD8+ T cells confer superior antitumor immunity compared with effector memory T cells. Proc. Natl. Acad. Sci. USA 2005, 102, 9571–9576. [Google Scholar] [CrossRef] [PubMed]
- Gattinoni, L.; Klebanoff, C.A.; Palmer, D.C.; Wrzesinski, C.; Kerstann, K.; Yu, Z.; Finkelstein, S.E.; Theoret, M.R.; Rosenberg, S.A.; Restifo, N.P. Acquisition of full effector function in vitro paradoxically impairs the in vivo antitumor efficacy of adoptively transferred CD8+ T Cells. J. Clin. Investig. 2005, 115, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Parrish-Novak, J.; Dillon, S.R.; Nelson, A.; Hammond, A.; Sprecher, C.; Gross, J.A.; Johnston, J.; Madden, K.; Xu, W.; West, J.; et al. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature 2000, 408, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Spolski, R.; Leonard, W.J. Interleukin-21: basic biology and implications for cancer and autoimmunity. Annu. Rev. Immunol. 2008, 26, 57–79. [Google Scholar] [CrossRef] [PubMed]
- Leonard, W.J.; Spolski, R. Interleukin-21: A modulator of lymphoid proliferation, apoptosis and differentiation. Nat. Rev. Immunol. 2005, 5, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Wolfl, M.; Merker, K.; Morbach, H.; van Gool, S.W.; Eyrich, M.; Greenberg, P.D.; Schlegel, P.G. Primed tumor-reactive multifunctional CD62L+ human CD8+ T cells for immunotherapy. Cancer Immunol. Immunother. 2011, 60, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Figliola, M.J.; Dawson, M.J.; Huls, H.; Olivares, S.; Switzer, K.; Mi, T.; Maiti, S.; Kebriaei, P.; Lee, D.A.; et al. Reprogramming CD19-Specific T Cells with IL-21 Signaling can improve adoptive immunotherapy of B-lineage malignancies. Cancer Res. 2011, 71, 3516–3527. [Google Scholar] [CrossRef] [PubMed]
- Chapuis, A.G.; Ragnarsson, G.B.; Nguyen, H.N.; Chaney, C.N.; Pufnock, J.S.; Schmitt, T.M.; Duerkopp, N.; Roberts, I.M.; Pogosov, G.L.; Ho, W.Y.; et al. Transferred WT1-Reactive CD8+ T cells can mediate antileukemic activity and persist in post-transplant patients. Sci. Transl. Med. 2013, 5. [Google Scholar] [CrossRef]
- Hinrichs, C.S.; Spolski, R.; Paulos, C.M.; Gattinoni, L.; Kerstann, K.W.; Palmer, D.C.; Klebanoff, C.A.; Rosenberg, S.A.; Leonard, W.J.; Restifo, N.P. IL-2 and IL-21 confer opposing differentiation programs to CD8+ T cells for adoptive immunotherapy. Blood 2008, 111, 5326–5333. [Google Scholar] [CrossRef] [PubMed]
- Brentjens, R.J.; Latouche, J.B.; Santos, E.; Marti, F.; Gong, M.C.; Lyddane, C.; King, P.D.; Larson, S.; Weiss, M.; Riviere, I.; et al. Eradication of systemic B-cell tumors by genetically targeted human T lymphocytes co-stimulated by CD80 and Interleukin-15. Nat. Med. 2003, 9, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, E.; Comes, A.; Orengo, A.M.; Rosso, O.; Meazza, R.; Musiani, P.; Colombo, M.P.; Ferrini, S. IL-21 Induces tumor rejection by specific CTL and IFN-γ-dependent CXC Chemokines in Syngeneic Mice. J. Immunol. 2004, 172, 1540–1547. [Google Scholar] [CrossRef]
- Klebanoff, C.A.; Finkelstein, S.E.; Surman, D.R.; Lichtman, M.K.; Gattinoni, L.; Theoret, M.R.; Grewal, N.; Spiess, P.J.; Antony, P.A.; Palmer, D.C.; et al. IL-15 enhances the in vivo antitumor activity of tumor-reactive CD8+ T cells. Proc. Natl. Acad. Sci. USA 2004, 101, 1969–1974. [Google Scholar] [CrossRef] [PubMed]
- Sondergaard, H.; Frederiksen, K.S.; Thygesen, P.; Galsgaard, E.D.; Skak, K.; Kristjansen, P.E.; Odum, N.; Kragh, M. Interleukin 21 therapy increases the density of tumor infiltrating CD8+ T cells and inhibits the growth of syngeneic tumors. Cancer Immunol. Immunother. 2007, 56, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Teague, R.M.; Sather, B.D.; Sacks, J.A.; Huang, M.Z.; Dossett, M.L.; Morimoto, J.; Tan, X.; Sutton, S.E.; Cooke, M.P.; Ohlen, C.; et al. Interleukin-15 rescues tolerant CD8+ T cells for use in adoptive immunotherapy of established tumors. Nat. Med. 2006, 12, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Spolski, R.; Finkelstein, S.E.; Oh, S.; Kovanen, P.E.; Hinrichs, C.S.; Pise-Masison, C.A.; Radonovich, M.F.; Brady, J.N.; Restifo, N.P.; et al. Synergy of IL-21 and IL-15 in regulating CD8+ T cell expansion and function. J. Exp. Med. 2005, 201, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Kasaian, M.T.; Whitters, M.J.; Carter, L.L.; Lowe, L.D.; Jussif, J.M.; Deng, B.; Johnson, K.A.; Witek, J.S.; Senices, M.; Konz, R.F.; et al. IL-21 limits NK Cell responses and promotes antigen-specific T cell activation: A mediator of the transition from innate to adaptive immunity. Immunity 2002, 16, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Moroz, A.; Eppolito, C.; Li, Q.; Tao, J.; Clegg, C.H.; Shrikant, P.A. IL-21 enhances and sustains CD8+ T cell responses to achieve durable tumor immunity: Comparative evaluation of IL-2, IL-15, and IL-21. J. Immunol. 2004, 173, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bleakley, M.; Yee, C. IL-21 Influences the frequency, phenotype, and affinity of the antigen-specific CD8 T cell response. J. Immunol. 2005, 175, 2261–2269. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, C.S.; Borman, Z.A.; Gattinoni, L.; Yu, Z.; Burns, W.R.; Huang, J.; Klebanoff, C.A.; Johnson, L.A.; Kerkar, S.P.; Yang, S.; et al. Human effector CD8+ T cells derived from naive rather than memory subsets possess superior traits for adoptive immunotherapy. Blood 2011, 117, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, J.; Frey, M.; Teschner, D.; Carbol, A.; Theobald, M.; Herr, W.; Distler, E. IL-21-treated naive CD45RA+CD8+ T cells represent a reliable source for producing leukemia-reactive cytotoxic T lymphocytes with high proliferative potential and early differentiation phenotype. Cancer Immunol. Immunother. 2011, 60, 235–248. [Google Scholar] [CrossRef] [PubMed]
- </b>Gattinoni, L.; Klebanoff, C.A.; Restifo, N.P. Paths to stemness: Building the ultimate antitumour T cell. Nat. Rev. Cancer 2012, 12, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Lipshy, K.A.; Kostuchenko, P.J.; Hamad, G.G.; Bland, C.E.; Barrett, S.K.; Bear, H.D. Sensitizing T-lymphocytes for adoptive immunotherapy by vaccination with wild-type or cytokine gene-transduced melanoma. Ann. Surg. Oncol. 1997, 4, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J.; Teichgraber, V.; Becker, T.C.; Masopust, D.; Kaech, S.M.; Antia, R.; von Andrian, U.H.; Ahmed, R. Lineage relationship and protective immunity of memory CD8 T cell subsets. Nat. Immunol. 2003, 4, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Parish, I.A.; Kaech, S.M. Diversity in CD8+ T cell differentiation. Curr. Opin. Immunol. 2009, 21, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Obar, J.J.; Lefrancois, L. Memory CD8+ T cell differentiation. Ann. N. Y. Acad. Sci. 2010, 1183, 251–266. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoon, C.K.; Wan, W.; Graham, L.; Bear, H.D. Addition of Interleukin-21 for Expansion of T-Cells for Adoptive Immunotherapy of Murine Melanoma. Int. J. Mol. Sci. 2015, 16, 8744-8760. https://doi.org/10.3390/ijms16048744
Zoon CK, Wan W, Graham L, Bear HD. Addition of Interleukin-21 for Expansion of T-Cells for Adoptive Immunotherapy of Murine Melanoma. International Journal of Molecular Sciences. 2015; 16(4):8744-8760. https://doi.org/10.3390/ijms16048744
Chicago/Turabian StyleZoon, Christine Kathryn, Wen Wan, Laura Graham, and Harry D. Bear. 2015. "Addition of Interleukin-21 for Expansion of T-Cells for Adoptive Immunotherapy of Murine Melanoma" International Journal of Molecular Sciences 16, no. 4: 8744-8760. https://doi.org/10.3390/ijms16048744
APA StyleZoon, C. K., Wan, W., Graham, L., & Bear, H. D. (2015). Addition of Interleukin-21 for Expansion of T-Cells for Adoptive Immunotherapy of Murine Melanoma. International Journal of Molecular Sciences, 16(4), 8744-8760. https://doi.org/10.3390/ijms16048744