
The Heart Protection Effect of Alcalase Potato Protein Hydrolysate Is through IGF1R-PI3K-Akt Compensatory Reactivation in Aging Rats on High Fat Diets

 ,
,
Abstract
:1. Introduction
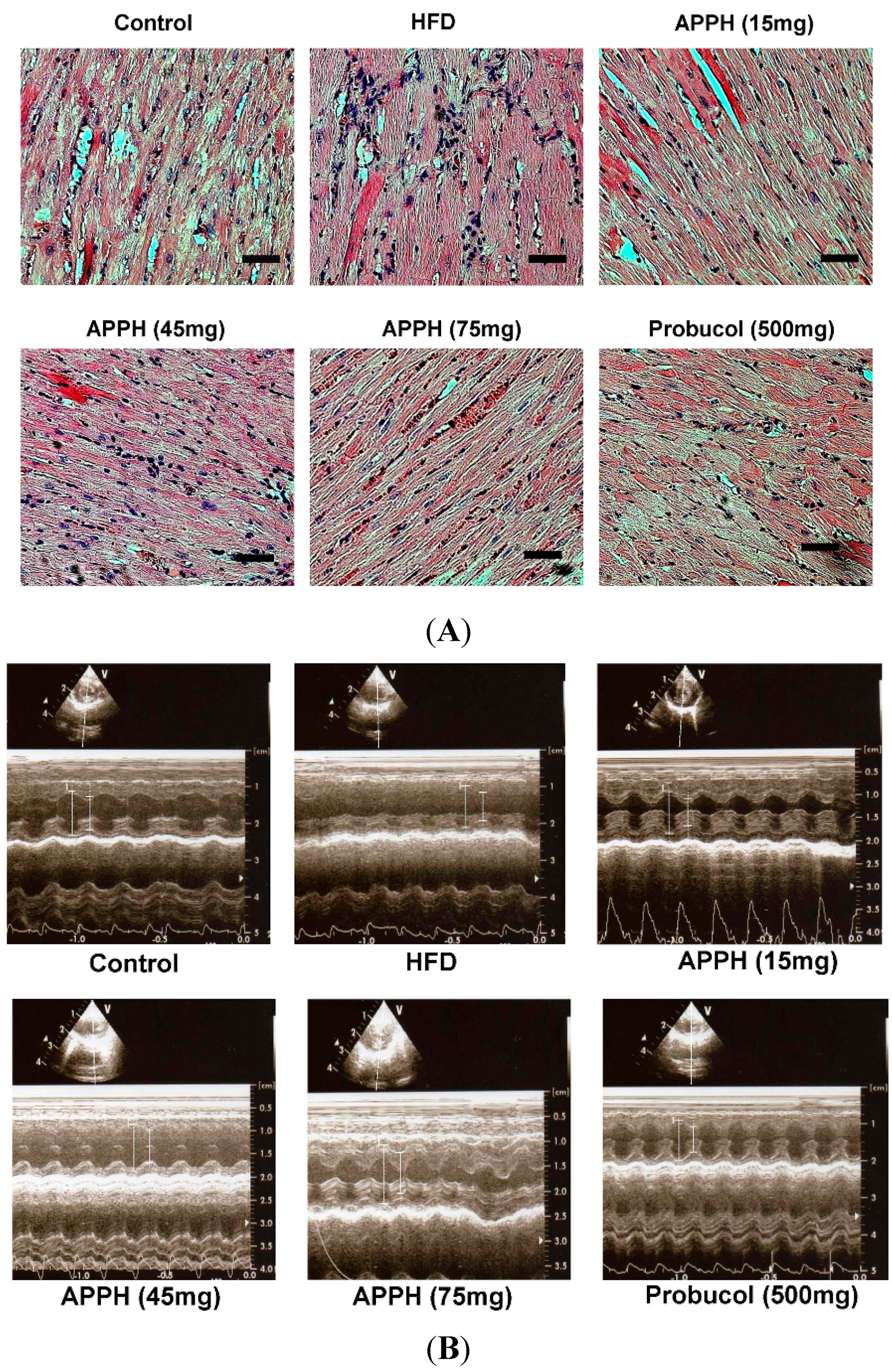
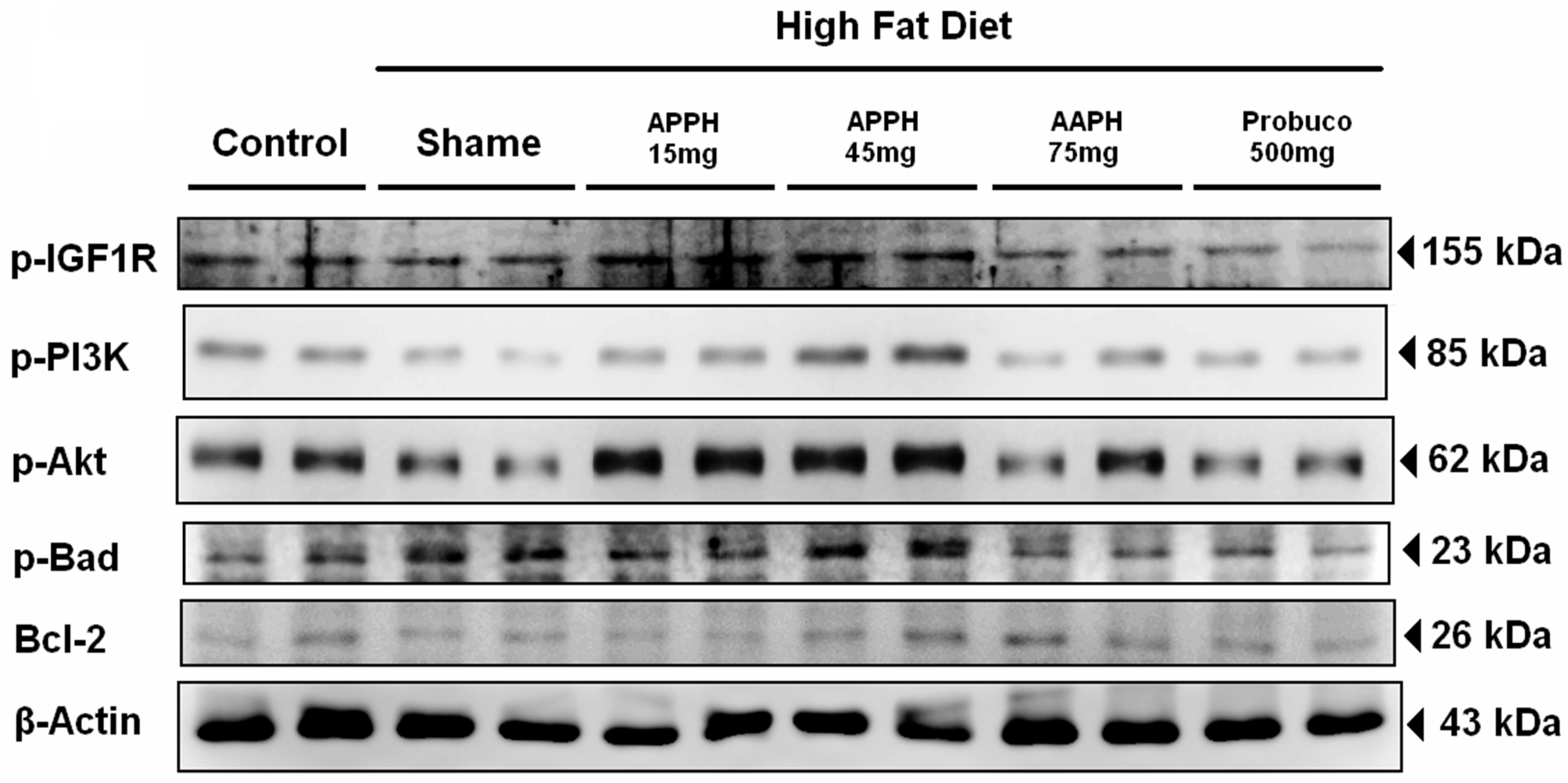
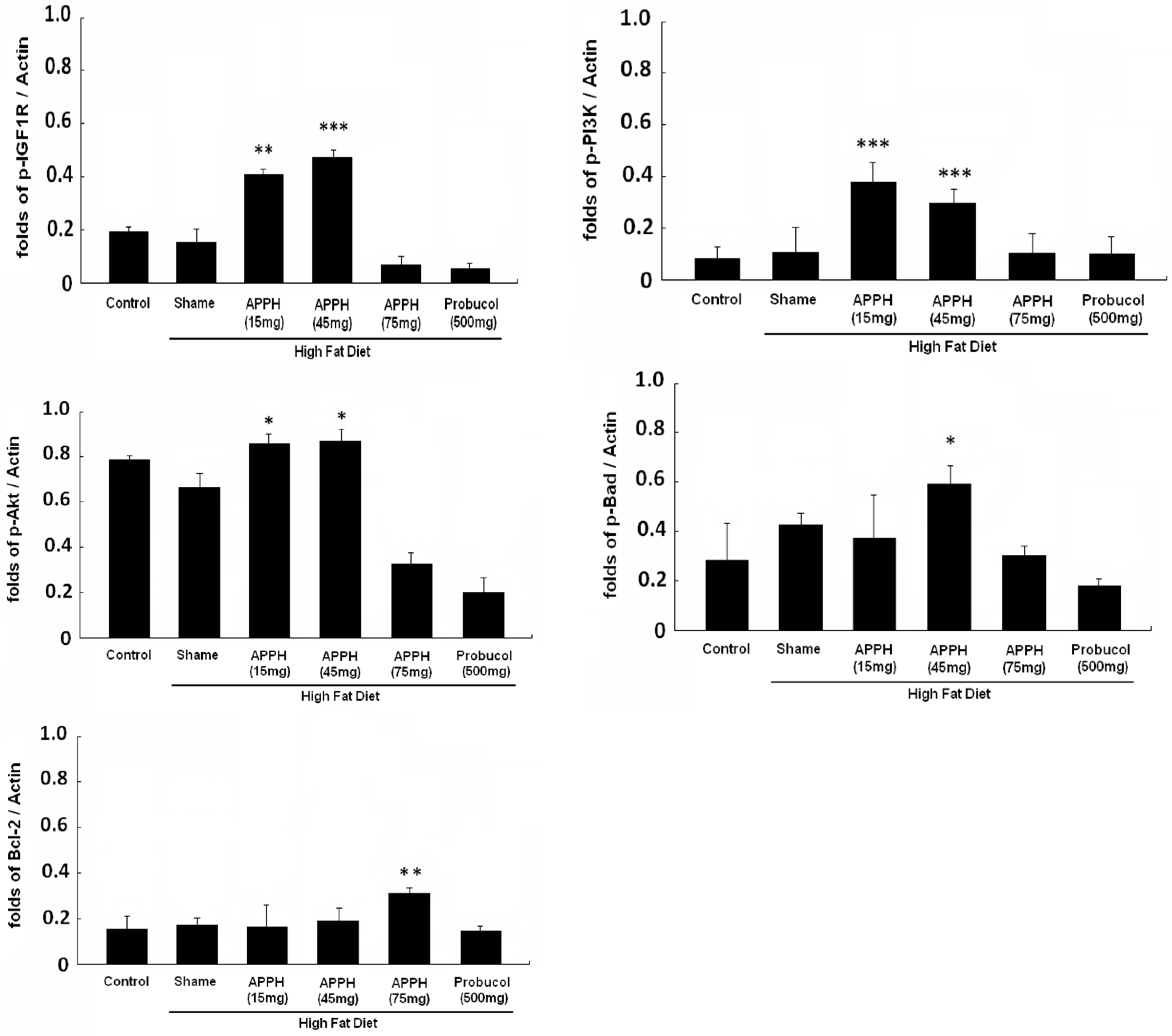
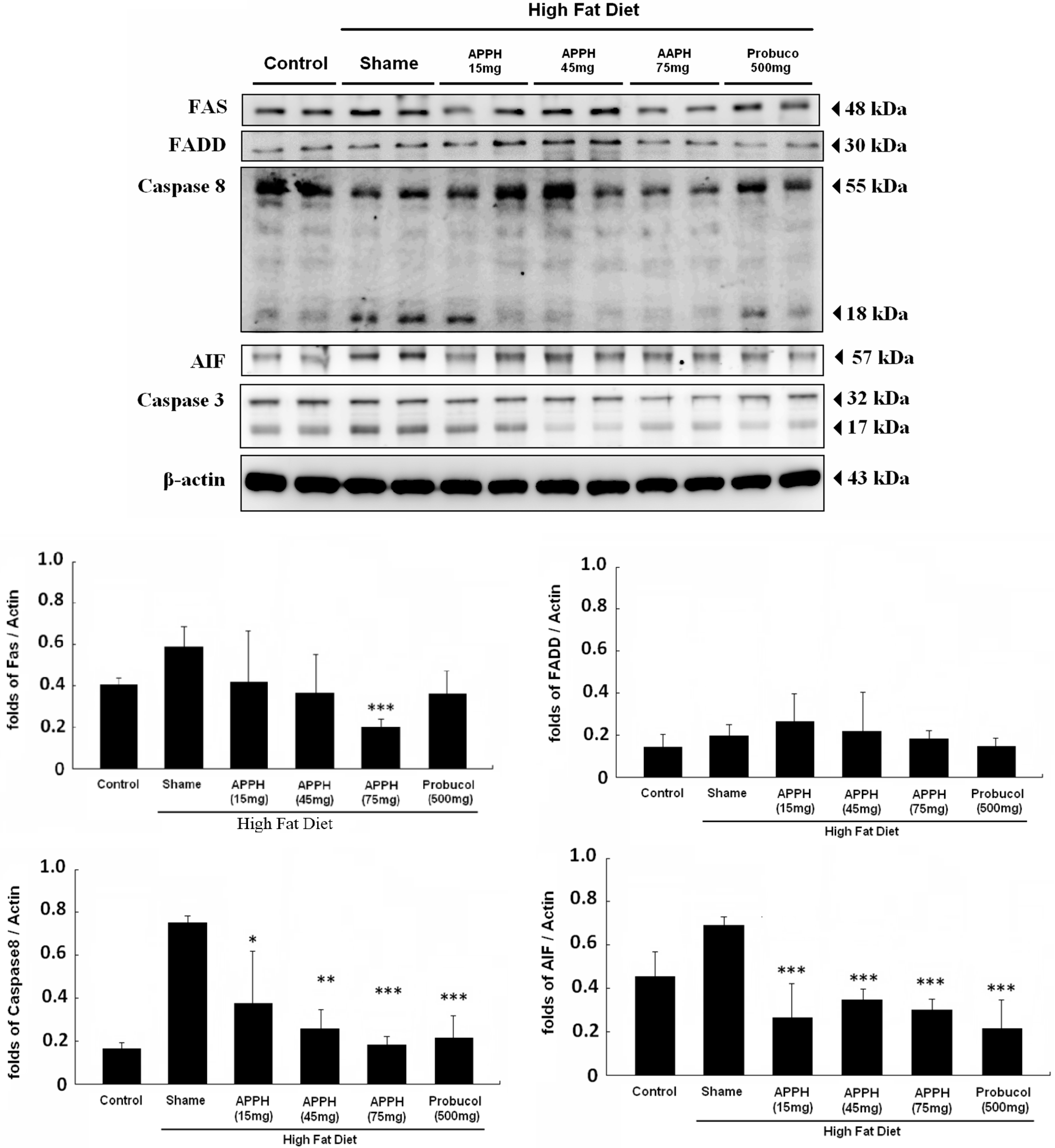
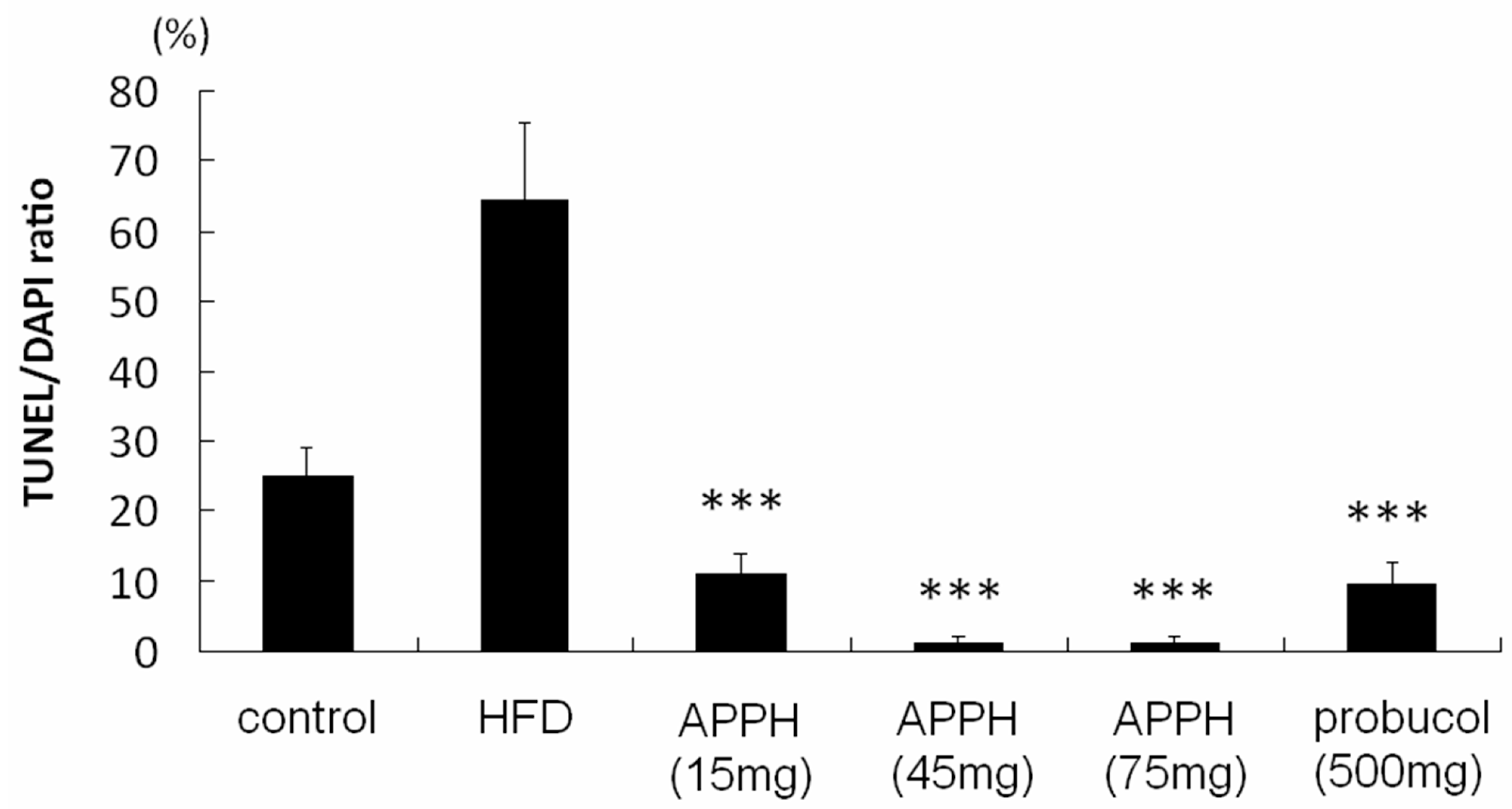
2. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Control | HFD | APPH (15 mg) | APPH (45 mg) | APPH (75 mg) | Probucol (500 mg) |
|---|---|---|---|---|---|---|
| HFD | HFD | HFD | HFD | |||
| BUN (mg/dL) | 14.4 ± 3.1 | 15.5 ± 1.7 | 13.6 ± 2.1 | 12.5 ± 2.6 | 12.6 ± 2.6 | 11.9 ± 1.2 |
| CRE (mg/dL) | 0.4 ± 0.1 | 0.5 ± 0.2 | 0.6 ± 0.1 | 0.5 ± 0.1 | 0.4 ± 0.1 | 0.5 ± 0.1 |
| TC (mg/dL) | 74.0 ± 20.7 a | 182.7 ± 43.4 | 121.3 ± 23.9 a | 96 ± 21.5 a | 89.7 ± 12.9 a | 84.3 ± 19.5 a |
| TG (mg/dL) | 122.3 ± 13.4 a | 152.7 ± 17.6 | 92.3 ± 12.1 a | 83.0 ± 19.7 a | 67.0 ± 16.6 a | 63.3 ± 11.0 a |
| HDL-C (mg/dL) | 27.0 ± 10.4 | 35.2 ± 8.8 | 34.7 ± 9.1 | 31.7 ± 7.8 | 27.6 ± 7.9 a | 23.3 ± 4.2 a |
| LDL-C (mg/dL) | 107.3 ± 12.9 a | 227.0 ± 14.6 | 185.5 ± 12.6 a | 92.3 ± 3.1 a | 81.0 ± 3.0 a | 74.3 ± 12.7 a |
| GLU (mg/dL) | 107.0 ± 35.2 a | 183.3 ± 39.6 | 173.7 ± 34.0 | 135.0 ± 20.1 a | 99.3 ± 13.3 a | 127.0 ± 32.0 a |

| Groups | Control | HFD | APPH (15 mg) | APPH (45 mg) | APPH (75 mg) | Probucol (500 mg) |
|---|---|---|---|---|---|---|
| HFD | HFD | HFD | HFD | |||
| BW (g) | 465.7 ± 63.9 a | 700.3 ± 88.6 | 690.3 ± 67.5 | 638.7 ± 71.6 | 607.2 ± 70.2 a | 688.2 ± 52.1 |
| EF (%) | 54.0 ± 5.2 | 54.0 ± 4.7 | 62.2 ± 4.6 | 67.5 ± 5.6 a | 66.4 ± 3.2 a | 69.4 ± 2.5 a |
| %FS (%) | 24.9 ± 3.0 | 24.9 ± 2.7 | 31.8 ± 2.9 a | 33.7 ± 4.1 a | 33.0 ± 2.2 a | 35.2 ± 1.8 a |






3. Experimental Section
3.1. Chemicals
3.2. Animals
3.3. Echocardiography
3.4. Hematoxylin and Eosin Staining
3.5. DAPI and TUNEL Staining
3.6. Blood Biochemical Analysis
3.7. Tissue Protein Extraction
3.8. Western Blot Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cynthia, L.O.; Margaret, D.C.; Brian, K.K.; Katherine, M.F. Prevalence of obesity among adults: United States, 2011–2012. NCHS Data Brief. 2013, 131, 1–8. [Google Scholar] [PubMed]
- Kranendonk, M.E.; de Kleijn, D.P.; Kalkhoven, E.; Kanhai, D.A.; Uiterwaal, C.S.; van der Graaf, Y.; Pasterkamp, G.; Visseren, F.L.; SMART Study Group. Extracellular vesicle markers in relation to obesity and metabolic complications in patients with manifest cardiovascular disease. Cardiovasc. Diabetol. 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Ruiz Ruiz, J.C.; Betancur Ancona, D.A.; Segura Campos, M.R. Bioactive vegetable proteins and peptides in lipid-lowering; nutraceutical potential. Nutr. Hosp. 2014, 29, 776–784. [Google Scholar]
- Singh, B.P.; Vij, S.; Hati, S. Functional significance of bioactive peptides derived from soybean. Peptides 2014, 54, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, K.; Cheung, B.W.; Schröder, H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; de Mejia, E.G. A new frontier in soy bioactive peptides that may prevent age-related chronic diseases. Compr. Rev. Food Sci. Food Saf. 2005, 4, 63–68. [Google Scholar] [CrossRef]
- Kudoh, K.; Matsumoto, M.; Onodera, S.; Takeda, Y.; Ando, K.; Shiomi, N. Antioxidative activity and protective effect against ethanol-induced gastric mucosal damage of a potato protein hydrolysate. J. Nutr. Sci. Vitaminol. 2003, 49, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xiong, Y.L.; McNear, D.H. Changes in structural characteristics of antioxidative soy protein hydrolysates resulting from scavenging of hydroxyl radicals. J. Food Sci. 2013, 78, 152–159. [Google Scholar] [CrossRef]
- Dinnella, C.; Gargaro, M.T.; Rossano, R.; Monteleone, E. Spectrophotometric assay using O-phtaldialdehyde for the determination of transglutaminase activity on casein. Food Chem. 2002, 78, 363–368. [Google Scholar] [CrossRef]
- Cherng, S.H.; Huang, C.Y.; Kuo, W.W.; Lai, S.E.; Tseng, C.Y.; Lin, Y.M.; Tsai, F.J.; Wang, H.F. GABA tea prevents cardiac fibrosis by attenuating TNF-α and Fas/FasL-mediated apoptosis in streptozotocin-induced diabetic rats. Food Chem. Toxicol. 2014, 65, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Yang, A.L.; Lin, Y.M.; Wu, F.N.; Lin, J.A.; Chan, Y.S.; Tsai, F.J.; Tsai, C.H.; Kuo, C.H.; Lee, S.D. Anti-apoptotic and pro-survival effects of exercise training on hypertensive hearts. J. Appl. Physiol. 2012, 112, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.S.; Lee, S.D.; Kuo, W.W.; Fan, M.J.; Lin, Y.M.; Hu, W.S.; Huang, Y.C.; Velmurugan, B.K.; Tsai, F.J.; Tsai, C.H.; et al. Anti-apoptotic and pro-survival effect of protocatechuic acid on hypertensive hearts. Chem. Biol. Interact. 2014, 209, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.M.; Ho, T.J.; Yang, A.L.; Chen, I.J.; Kao, C.L.; Wu, F.N.; Lin, J.A.; Kuo, C.H.; Ou, H.C.; Huang, C.Y.; et al. Exercise training enhances cardiac IGFI-R/PI3K/Akt and Bcl-2 family associated pro-survival pathways in streptozotocin-induced diabetic rats. Int. J. Cardiol. 2013, 167, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.P.; Hsieh, Y.M.; Kuo, W.W.; Lin, Y.M.; Yeh, Y.L.; Lin, C.C.; Tsai, F.J.; Tsai, C.H.; Huang, C.Y.; Tsai, C.C. Probiotic-fermented purple sweet potato yogurt activates compensatory IGF-IR/PI3K/Akt survival pathways and attenuates cardiac apoptosis in the hearts of spontaneously hypertensive rats. Int. J. Mol. Med. 2013, 32, 1319–1328. [Google Scholar] [PubMed]
- Dutta, D.; Xu, J.; Dirain, M.L.; Leeuwenburgh, C. Calorie restriction combined with resveratrol induces autophagy and protects 26-month-old rat hearts from doxorubicin-induced toxicity. Free Radic. Biol. Med. 2014, 74, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Favari, E.; Zanotti, I.; Zimetti, F.; Ronda, N.; Bernini, F.; Rothblat, G.H. Probucol inhibits ABCA1-mediated cellular lipid efflux. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2345–2350. [Google Scholar] [CrossRef] [PubMed]
- Niimi, M.; Keyamura, Y.; Nozako, M.; Koyama, T.; Kohashi, M.; Yasufuku, R.; Yoshikawa, T.; Fan, J. Probucol inhibits the initiation of atherosclerosis in cholesterol-fed rabbits. Lipids Health Dis. 2013, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, M.; Nakagawa-Toyama, Y.; Nishida, M.; Nakatani, K.; Nakaoka, H.; Kawase, M.; Kawase, R.; Tsubakio-Yamamoto, K.; Masuda, D.; Ohama, T.; et al. Effect of probucol on antioxidant properties of HDL in patients with heterozygous familial hypercholesterolemia. J. Atheroscler. Thromb. 2012, 19, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Miida, T.; Seino, U.; Miyazaki, O.; Hanyu, O.; Hirayama, S.; Saito, T.; Ishikawa, Y.; Akamatsu, S.; Nakano, T.; Nakajima, K.; et al. Probucol markedly reduces HDL phospholipids and elevated preβ1-HDL without delayed conversion into α-migrating HDL: Putative role of angiopoietin-like protein 3 in probucol-induced HDL remodeling. Atherosclerosis 2008, 200, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Spielmann, J.; Kluge, H.; Stangl, G.I.; Eder, K. Hypolipidaemic effects of potato protein and fish protein in pigs. J. Anim. Physiol. Anim. Nutr. 2009, 93, 400–409. [Google Scholar] [CrossRef]
- Knorr, D.; Kohler, G.O.; Betschart, A.A. Potato protein concentrates: The influence of various methods of recovery upon yield, compositional and functional characteristics. J. Food Process. Preserv. 1977, 1, 235–247. [Google Scholar] [CrossRef]
- Wang, L.L.; Xiong, Y.L. Inhibition of lipid oxidation in cooked beef patties by hydrolyzed potato protein is related to its reducing and radical scavenging ability. J. Agric. Food Chem. 2005, 53, 9186–9192. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Chiang, W.D.; Pai, P.; Lin, W.T. Potato protein hydrolysate attenuates high fat diet-induced cardiac apoptosis through SIRT1/PGC-1α/Akt signaling. J. Funct. Food 2015, 12, 389–398. [Google Scholar] [CrossRef]
- Becker, A.E.; Caruso, G. Myocardial disarray. A critical review. Br. Heart J. 1982, 47, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Sobkiw, C.L.; Hirshman, M.F.; Logsdon, M.N.; Li, T.Q.; Goodyear, L.J.; Cantley, L.C. Loss of class IA PI3K signaling in muscle leads to impaired muscle growth, insulin response, and hyperlipidemia. Cell Metable 2006, 3, 355–366. [Google Scholar] [CrossRef]
- Huang, C.Y.; Lee, S.D. Possible pathophysiology of heart failure in obesity: Cardiac apoptosis. BioMedicine 2012, 2, 36–40. [Google Scholar] [CrossRef]
- Yin, H.; Chao, L.; Chao, J. Kallikrein/kinin protects against myocardial apoptosis after ischemia/reperfusion via Akt-glycogen synthase kinase-3 and Akt-Bad.14-3-3 signaling pathways. J. Biol. Chem. 2005, 280, 8022–8030. [Google Scholar] [CrossRef] [PubMed]
- Chakir, K.; Daya, S.K.; Tunin, R.S.; Helm, R.H.; Byrne, M.J.; Dimaano, V.L.; Lardo, A.C.; Abraham, T.P.; Tomaselli, G.F.; Kass, D.A. Reversal of global apoptosis and regional stress kinase activation by cardiac resynchronization. Circulation 2008, 117, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.W.; Chu, C.Y.; Wu, C.H.; Lin, J.A.; Liu, J.Y.; Hsieh, Y.H.; Ueng, K.C.; Lee, S.D.; Hsieh, D.J.; Hsu, H.H.; et al. Impaired IGF-I signalling of hypertrophic hearts in the developmental phase of hypertension in genetically hypertensive rats. Cell Biochem. Funct. 2005, 23, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.F.; Lin, P.P.; Chen, C.H.; Yeh, Y.L.; Huang, C.C.; Huang, C.Y.; Tsai, C.C. Effects of lactic acid bacteria on cardiac apoptosis are mediated by activation of the phosphatidylinositol-3 kinase/Akt survival-signalling pathway in rats fed a high-fat diet. Int. J. Mol. Med. 2015, 35, 460–470. [Google Scholar] [PubMed]
- Udenigwe, C.C.; Je, J.Y.; Cho, Y.S.; Yada, R.Y. Almond protein hydrolysate fraction modulates the expression of proinflammatory cytokines and enzymes in activated macrophages. Food Funct. 2013, 4, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Haffar, T.; Bérubé-Simard, F.A.; Bousette, N. Cardiomyocyte lipotoxicity is mediated by IL-6 and causes down-regulation of PPARs. Biochem. Biophys. Res. Commun. 2015, 459, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Turkieh, A.; Caubère, C.; Barutaut, M.; Desmoulin, F.; Harmancey, R.; Galinier, M.; Berry, M.; Dambrin, C.; Polidori, C.; Casteilla, L.; et al. Apolipoprotein O is mitochondrial and promotes lipotoxicity in heart. J. Clin. Investig. 2014, 124, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.F.; Luvizotto, R.A.; Leopoldo, A.S.; Lima-Leopoldo, A.P.; Seiva, F.R.; Justulin, L.A., Jr.; Silva, M.D.; Okoshi, K.; Wang, X.D.; Cicogna, A.C. Long-term high-fat diet-induced obesity decreases the cardiac leptin receptor without apparent lipotoxicity. Life Sci. 2011, 88, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zhang, Z.; Chen, Q.; Yin, X.; Fu, Y.; Zheng, Y.; Cai, L.; Kim, K.S.; Kim, K.H.; Tan, Y.; et al. Magnolia extract (BL153) protection of heart from lipid accumulation caused cardiac oxidative damage, inflammation, and cell death in high-fat diet fed mice. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, W.-S.; Ting, W.-J.; Chiang, W.-D.; Pai, P.; Yeh, Y.-L.; Chang, C.-H.; Lin, W.-T.; Huang, C.-Y. The Heart Protection Effect of Alcalase Potato Protein Hydrolysate Is through IGF1R-PI3K-Akt Compensatory Reactivation in Aging Rats on High Fat Diets. Int. J. Mol. Sci. 2015, 16, 10158-10172. https://doi.org/10.3390/ijms160510158
Hu W-S, Ting W-J, Chiang W-D, Pai P, Yeh Y-L, Chang C-H, Lin W-T, Huang C-Y. The Heart Protection Effect of Alcalase Potato Protein Hydrolysate Is through IGF1R-PI3K-Akt Compensatory Reactivation in Aging Rats on High Fat Diets. International Journal of Molecular Sciences. 2015; 16(5):10158-10172. https://doi.org/10.3390/ijms160510158
Chicago/Turabian StyleHu, Wei-Syun, Wei-Jen Ting, Wen-Dee Chiang, Peiying Pai, Yu-Lan Yeh, Chung-Ho Chang, Wan-Teng Lin, and Chih-Yang Huang. 2015. "The Heart Protection Effect of Alcalase Potato Protein Hydrolysate Is through IGF1R-PI3K-Akt Compensatory Reactivation in Aging Rats on High Fat Diets" International Journal of Molecular Sciences 16, no. 5: 10158-10172. https://doi.org/10.3390/ijms160510158