
Differences between Mice and Humans in Regulation and the Molecular Network of Collagen, Type III, Alpha-1 at the Gene Expression Level: Obstacles that Translational Research Must Overcome


Abstract
:1. Introduction
2. Results
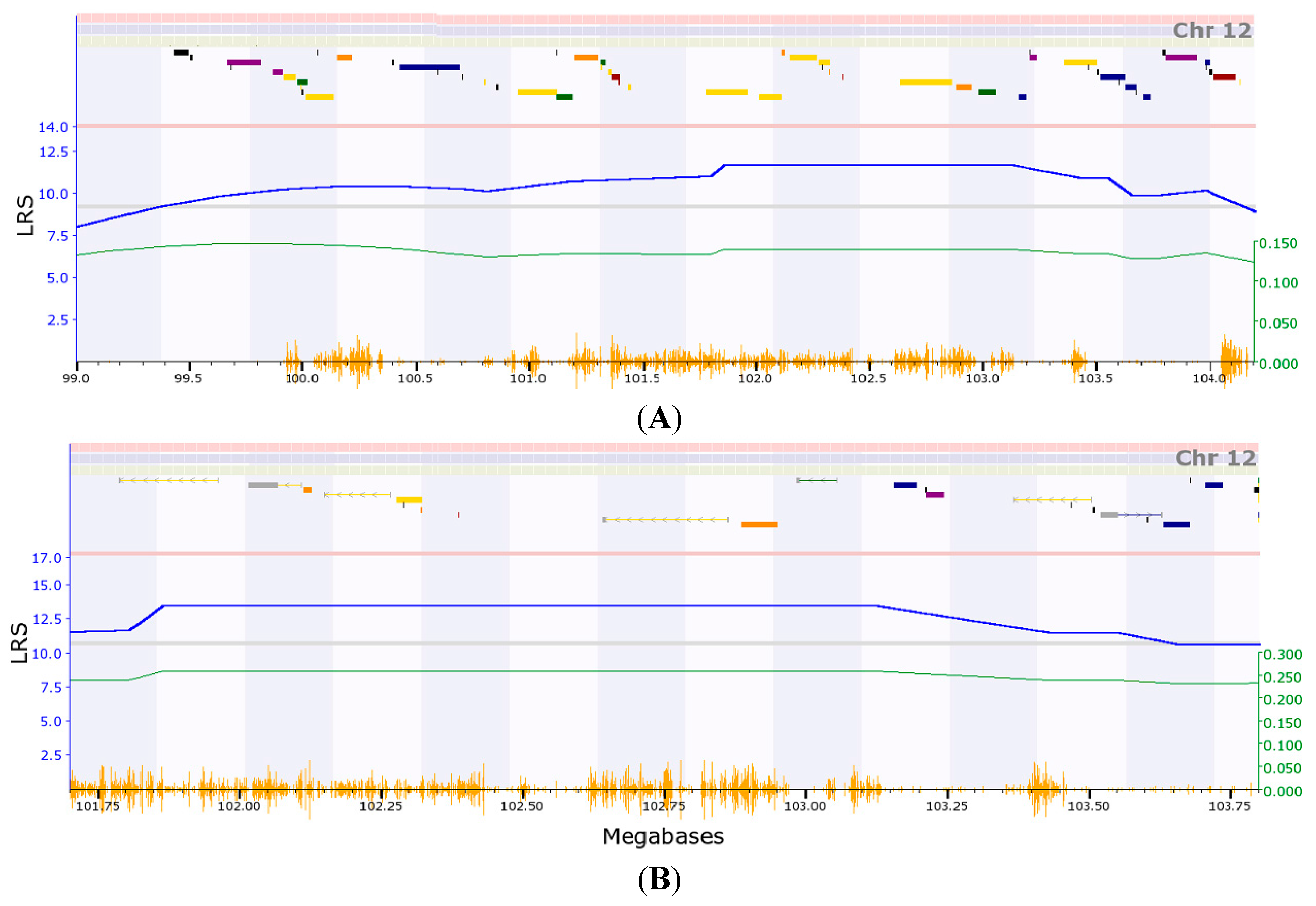
2.1. Transcriptome Mapping of Expression Quantitative Trait Loci (eQTL) that Regulates Expression Level of Collagen, Type III, alpha-1 (COL3A1)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | SNPS Name | Chr | MB | −log(p) | Gene Name | Function Consequence |
|---|---|---|---|---|---|---|
| 1 | exm-rs2585897 | 13 | 21.398979 | 4.82131 | XPO4 | intron variant |
| 2 | exm1564625 | 21 | 30.925928 | 4.69919 | – | – |
| 3 | exm2272985 | 21 | 42.047446 | 4.29611 | – | – |
| 4 | exm2270333 | 6 | 58.344955 | 4.04571 | CRIM1 | intron variant |
| 5 | exm609871 | 7 | 23.811795 | 3.93442 | – | – |
| 6 | exm-rs7326068 | 13 | 21.209512 | 3.69486 | IFT88 | intron variant |
| 7 | exm-rs10506525 | 12 | 65.783378 | 3.52754 | MSRB3 | intron variant |
| 8 | exm1082069 | 13 | 114.175038 | 3.51385 | – | – |
| 9 | exm1157141 | 15 | 44.093927 | 3.45730 | ZFPM2 | intron variant |
| 10 | exm2266405 | 7 | 46.919134 | 3.44515 | CCDC144A | intron variant |
| 11 | exm2266494 | 7 | 153.144983 | 3.27992 | – | – |
| 12 | exm-rs4380451 | 3 | 32.329497 | 3.27737 | CMTM8 | intron variant |
| 13 | exm2269464 | 3 | 69.422051 | 3.25987 | PHEX | intron variant |
| 14 | exm2271745 | 12 | 65.810174 | 3.25088 | – | – |
| 15 | exm506442 | 5 | 176.637576 | 3.21638 | TLDC1 | intron variant |
| 16 | exm869284 | 11 | 0.193146 | 3.17341 | – | – |
| 17 | exm2267394 | 12 | 117.082846 | 3.14527 | GTPBP1 | upstream variant 2 KB |
| 18 | exm2273373 | 3 | 52.874288 | 3.13365 | GPCPD1 | missense |
| 19 | exm882858 | 11 | 5.461834 | 3.11867 | – | – |
| 20 | exm-rs2286720 | 3 | 42.448471 | 3.10924 | LYZL4 | synonymous codon |
| 21 | exm-rs2763196 | 9 | 117.150951 | 3.10885 | AKNA | intron variant, upstream variant 2 KB |
| 22 | exm-rs11073328 | 15 | 38.764843 | 3.10746 | FAM98B | intron variant |
| 23 | exm397086 | 4 | 42.895308 | 3.09675 | MEGF10 | intron variant |
| 24 | exm1156850 | 15 | 43.939642 | 3.03829 | RP11-23B15.1 | intron variant |
2.2. Initial Analysis of Candidate Genes for Regulation of Expression of Col3a1 on Mouse Chromosome (Chr) 12

2.3. Narrowing down the Candidate Genes for Regulation of Expression of Col3a1 on Mouse Chr 12
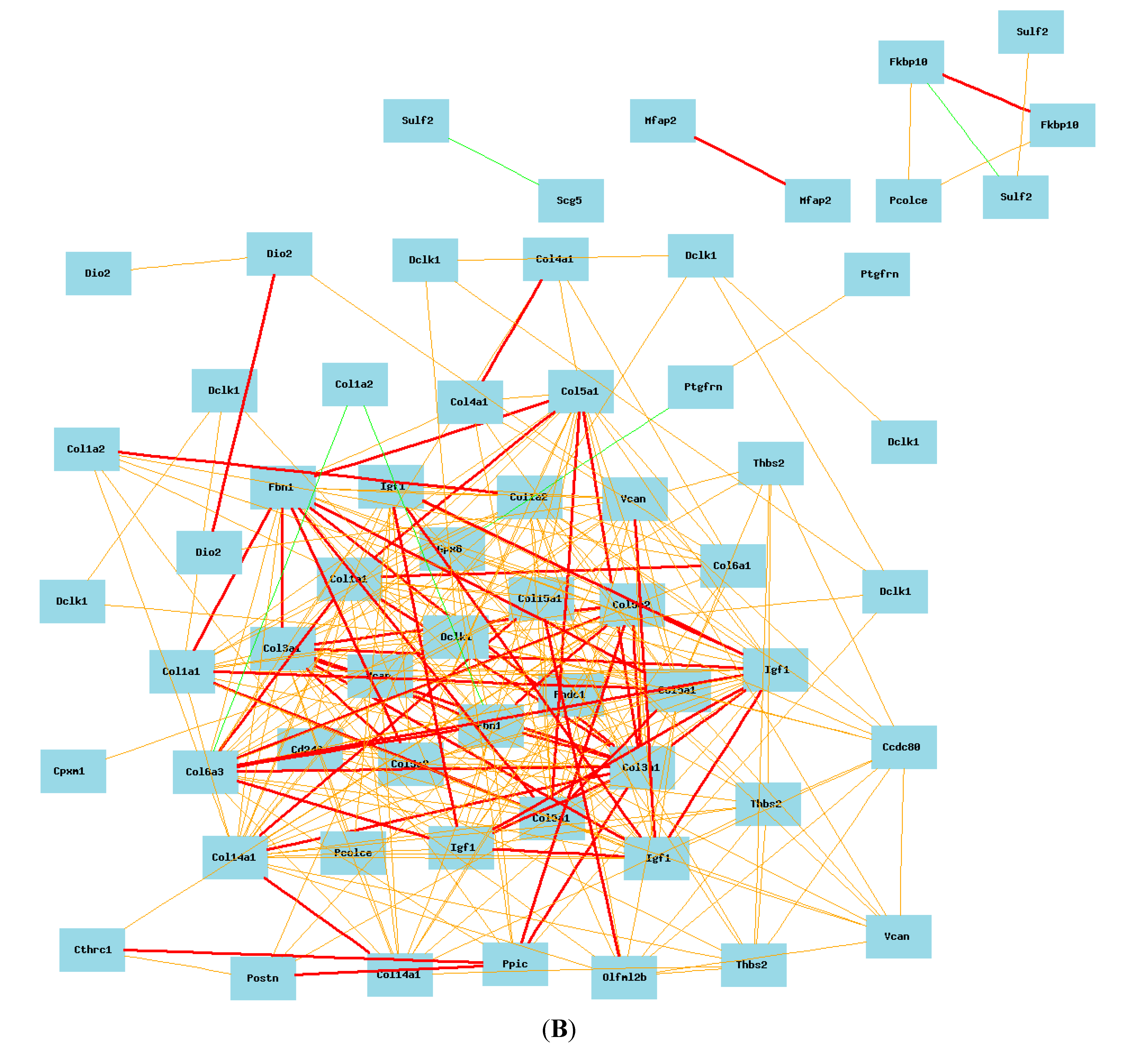
2.4. Gene Network of Col3a1-Related Genes in Lung in Mouse Model


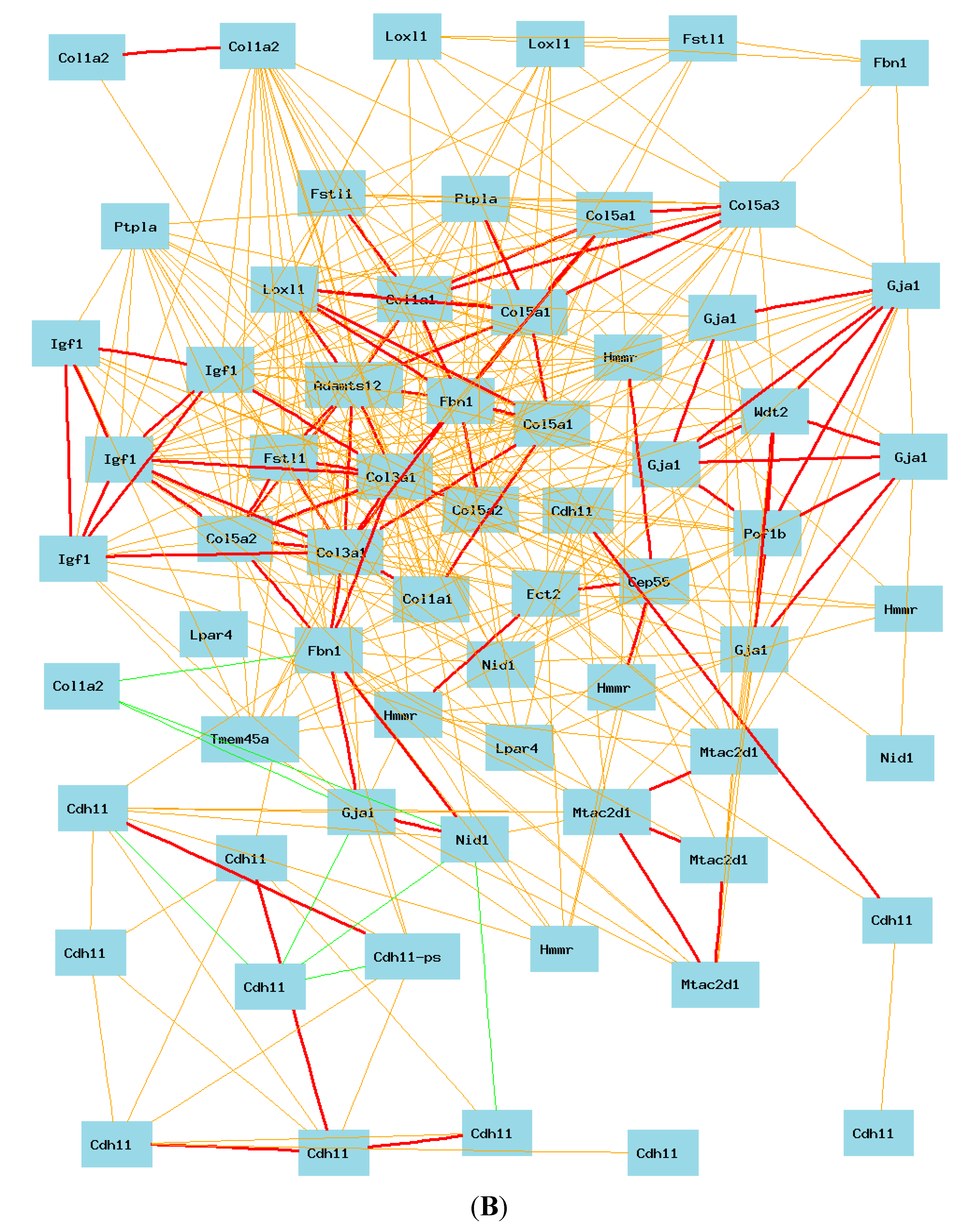

2.5. Gene Network of COL3A1 in Lung in Humans
2.6. Potential Difference in Regulation of Expression of COL3A1 between Human and Mice
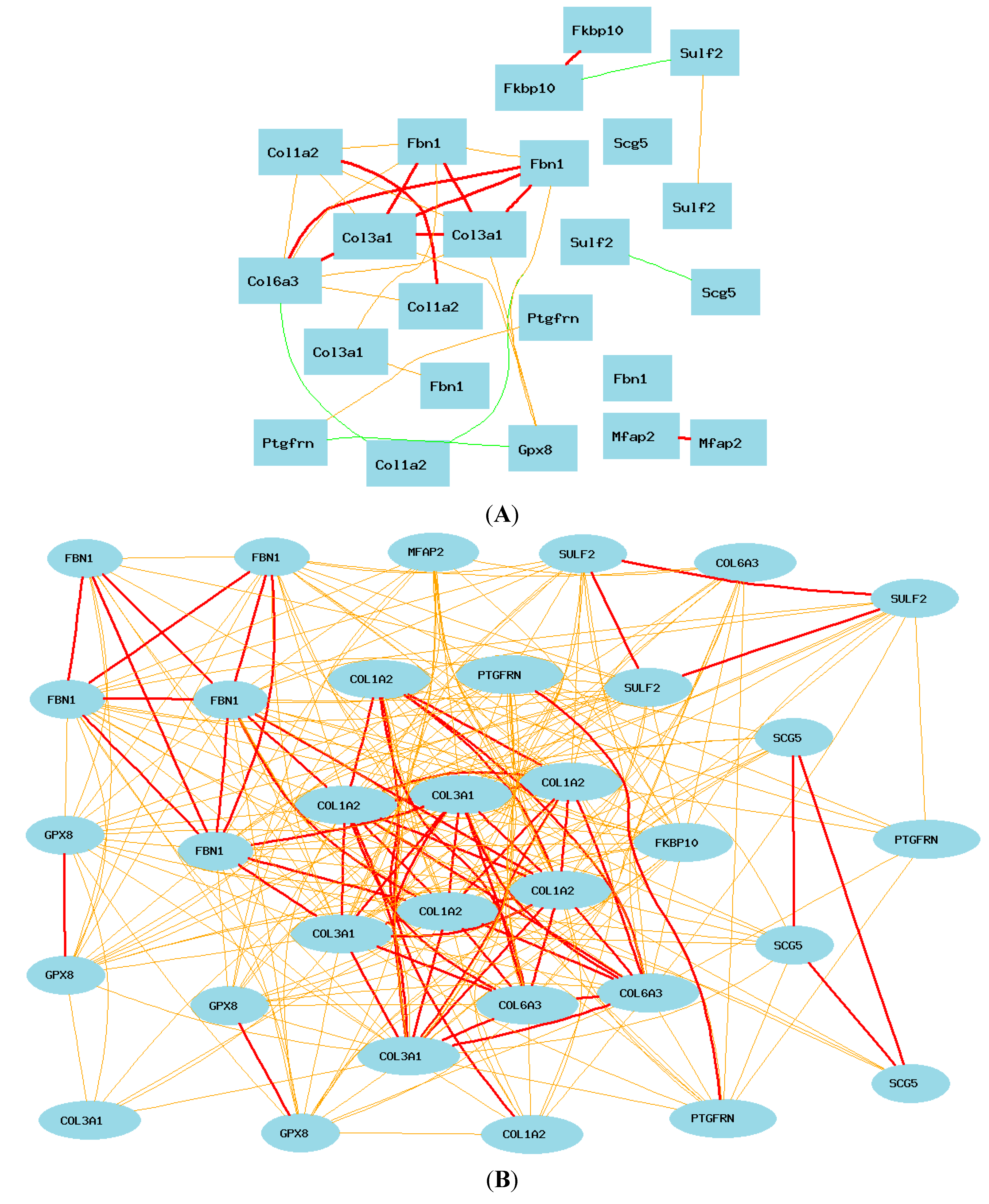
2.7. Gene Network between Mice and Humans


2.8. Potential Differences in Gene Network of COL3A1 between Humans and Mice




3. Discussion
4. Experimental Section
4.1. Mouse Gene Expression Data Sets for Lung
4.2. Human Gene Expression Data for Lung
4.3. eQTL Mapping
4.4. Gene Network Construction
4.5. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cole, W.G.; Chiodo, A.A.; Lamande, S.R.; Janeczko, R.; Ramirez, F.; Dahl, H.-H.M.; Chan, D.; Bateman, J.F. A base substitution at a splice site in the COL3A1 gene causes exon skipping and generates abnormal type III procollagen in a patient with Ehlers-Danlos syndrome type IV. J. Biol. Chem. 1990, 265, 17070–17077. [Google Scholar] [PubMed]
- Schwarze, U.; Schievink, W.I.; Petty, E.; Jaff, M.R.; Babovic-Vuksanovic, D.; Cherry, K.J.; Pepin, M.; Byers, P.H. Haploinsufficiency for one COL3A1 allele of type III procollagen results in a phenotype similar to the vascular form of Ehlers-Danlos syndrome, Ehlers-Danlos syndrome type IV. Am. J. Hum. Genet. 2001, 69, 989–1001. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, H.; Byrne, M.; Krane, S.; Aenisch, R. Type III collagen is crucial for collagen I fibrillogenesis and for normal cardiovascular development. Proc. Natl. Acad. Sci. USA 1997, 94, 1852–1856. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.K.; Zhong, Q.; Krawczyk, M.; Tae, H.J.; Müller, G.A.; Schubert, R.; Myers, L.A.; Dietz, H.C.; Talan, M.I.; Briest, W. The haploinsufficient Col3a1 mouse as a model for vascular Ehlers-Danlos syndrome. Vet. Pathol. 2010, 47, 1028–1039. [Google Scholar] [CrossRef] [PubMed]
- Christner, P.J.; Yufit, T.; Peters, J.; McGrath, R.; Jiménez, S.A. Transcriptional activation of alpha 1(III) procollagen gene in Tsk2/+ dermal fibroblasts. Biochem. Biophys. Res. Commun. 2003, 303, 406–412. [Google Scholar] [CrossRef]
- Long, K.B.; Li, Z.; Burgwin, C.M.; Choe, S.G.; Martyanov, V.; Sassi-Gaha, S.; Earl, J.P.; Eutsey, R.A.; Ahmed, A.; Ehrlich, G.D.; et al. The Tsk2/+ mouse fibrotic phenotype is due to a gain-of-function mutation in the PIIINP segment of the Col3a1 gene. J. Investig. Dermatol. 2015, 135, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, H.; Hock, T.; Thannickal, V.J.; Sanders, Y.Y. Histone deacetylase inhibition downregulates collagen 3A1 in fibrotic lung fibroblasts. Int. J. Mol. Sci. 2013, 14, 19605–19617. [Google Scholar] [CrossRef] [PubMed]
- Sa, Y.J.; Kim, Y.D.; Moon, S.W.; Kim, C.K.; Ki, C.S. Occlusive vascular Ehlers-Danlos syndrome accompanying a congenital cystic adenomatoid malformation of the lung: Report of a case. Surg. Today 2013, 43, 1467–1469. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhu, L.; Li, M. Antifibrotic effects of crocetin in scleroderma fibroblasts and in bleomycin-induced sclerotic mice. Clinics (Sao Paulo) 2013, 68, 1350–1357. [Google Scholar] [CrossRef]
- Artlett, C.M.; Sassi-Gaha, S.; Rieger, J.L.; Boesteanu, A.C.; Feghali-Bostwick, C.A.; Katsikis, P.D. The inflammasome activating caspase 1 mediates fibrosis and myofibroblast differentiation in systemic sclerosis. Arthritis Rheum. 2011, 63, 3563–3574. [Google Scholar] [CrossRef] [PubMed]
- Sabo-Attwood, T.; Ramos-Nino, M.E.; Eugenia-Ariza, M.; Macpherson, M.B.; Butnor, K.J.; Vacek, P.C.; McGee, S.P.; Clark, J.C.; Steele, C.; Mossman, B.T. Osteopontin modulates inflammation, mucin production, and gene expression signatures after inhalation of asbestos in a murine model of fibrosis. Am. J. Pathol. 2011, 178, 1975–1985. [Google Scholar] [CrossRef] [PubMed]
- Pommerenke, C.; Wilk, E.; Srivastava, B.; Schulze, A.; Novoselova, N.; Geffers, R.; Schughart, K. Global transcriptome analysis in influenza-infected mouse lungs reveals the kinetics of innate and adaptive host immune responses. PLoS ONE 2012, 7, e41169. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Qiu, Y.; Gu, W. PGMapper: A web-based tool linking phenotype to genes. Bioinformatics 2008, 24, 1011–1013. [Google Scholar] [CrossRef] [PubMed]
- Bossé, Y.; Postma, D.S.; Sin, D.D.; Lamontagne, M.; Couture, C.; Gaudreault, N.; Joubert, P.; Wong, V.; Elliott, M.; van den Berge, M.; et al. Molecular signature of smoking in human lung tissues. Cancer Res. 2012, 72, 3753–3763. [Google Scholar] [CrossRef] [PubMed]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, M.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haley, C.S.; Knott, S.A. A simple regression method for mapping quantitative trait loci in line crosses using flanking markers. Heredity (Edinb.) 1992, 69, 315–324. [Google Scholar]
- Wang, L.; Lu, W.; Zhang, L.; Huang, Y.; Scheib, R.; Liu, X.; Myers, L.; Lu, L.; Farber, C.R.; Liu, G.; et al. Trps1 differentially modulates the bone mineral density between male and female mice and its polymorphism associates with BMD differently between women and men. PLoS ONE 2014, 9, e84485. [Google Scholar]
- Huang, G.J.; Shifman, S.; Valdar, W.; Johannesson, M.; Yalcin, B.; Taylor, M.S.; Taylor, J.M.; Mott, R.; Flint, J. High resolution mapping of expression QTLs in heterogeneous stock mice in multiple tissues. Genome Res. 2009, 19, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Lu, J.; Auduong, L.; Sides, M.D.; Lasky, J.A. Overexpression of Sulf2 in idiopathic pulmonary fibrosis. Glycobiology 2013, 23, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Paun, A.; Lemay, A.M.; Tomko, T.G.; Haston, C.K. Association analysis reveals genetic variation altering bleomycin-induced pulmonary fibrosis in mice. Am. J. Respir. Cell Mol. Biol. 2013, 48, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Patterson, C.E.; Abrams, W.R.; Wolter, N.E.; Rosenbloom, J.; Davis, E.C. Developmental regulation and coordinate reexpression of FKBP65 with extracellular matrix proteins after lung injury suggest a specialized function for this endoplasmic reticulum immunophilin. Cell Stress Chaperones 2005, 10, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Artigas, M.S.; Loth, D.W.; Wain, L.V.; Gharib, S.A.; Obeidat, M.; Tang, W.; Zhai, G.; Zhao, J.H.; Smith, A.V.; Huffman, J.E.; et al. Genome-wide association and large-scale follow up identifies 16 new loci influencing lung function. Nat. Genet. 2011, 43, 1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Winter, E.E.; Wang, H.; Weinstock, K.G.; Xing, H.; Goodstadt, L.; Stenson, P.D.; Cooper, D.N.; Smith, D.; Albà, M.M.; et al. Evolutionary conservation and selection of human disease gene orthologs in the rat and mouse genomes. Genome Biol. 2004, 5, R47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondrashov, A.S.; Shabalina, S.A. Classification of common conserved sequences in mammalian intergenic regions. Hum. Mol. Genet. 2002, 11, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Swergold, G.D.; Seldin, M.F. Examination of sequence homology between human chromosome 20 and the mouse genome: Intense conservation of many genomic elements. Hum. Genet. 2003, 113, 60–70. [Google Scholar] [PubMed]
- Pevzner, P.; Tesler, G. Human and mouse genomic sequences reveal extensive breakpoint reuse in mammalian evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 7672–7677. [Google Scholar] [CrossRef] [PubMed]
- Christmann, R.B.; Sampaio-Barros, P.; Stifano, G.; Borges, C.L.; de Carvalho, C.R.; Kairalla, R.; Parra, E.R.; Spira, A.; Simms, R.; Capellozzi, V.L.; et al. Association of Interferon- and transforming growth factor β regulated genes and macrophage activation with systemic sclerosis-related progressive lung fibrosis. Arthritis Rheumatol. 2014, 66, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Chen, J.; Schlueter, C.F.; Hoyle, G.W. Differential susceptibility of inbred mouse strains to chlorine-induced airway fibrosis. Am. J. Physiol. Lung Cell Mol. Physiol. 2013, 304, L92–L102. [Google Scholar] [CrossRef] [PubMed]
- Haston, C.K.; Cory, S.; Lafontaine, L.; Dorion, G.; Hallett, M.T. Strain-dependent pulmonary gene expression profiles of a cystic fibrosis mouse model. Physiol. Genomics 2006, 25, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.M.; Crowell, T.P.; McCrann, M.E.; Frew, E.M.; Gardner, H. Analysis of the tight skin (Tsk1/+) mouse as a model for testing antifibrotic agents. Lab. Investig. 2005, 85, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Liu, H.; Jiao, Y.; Wang, E.; Clark, S.H.; Postlethwaite, A.E.; Gu, W.; Chen, H. Differences between Mice and Humans in Regulation and the Molecular Network of Collagen, Type III, Alpha-1 at the Gene Expression Level: Obstacles that Translational Research Must Overcome. Int. J. Mol. Sci. 2015, 16, 15031-15056. https://doi.org/10.3390/ijms160715031
Wang L, Liu H, Jiao Y, Wang E, Clark SH, Postlethwaite AE, Gu W, Chen H. Differences between Mice and Humans in Regulation and the Molecular Network of Collagen, Type III, Alpha-1 at the Gene Expression Level: Obstacles that Translational Research Must Overcome. International Journal of Molecular Sciences. 2015; 16(7):15031-15056. https://doi.org/10.3390/ijms160715031
Chicago/Turabian StyleWang, Lishi, Hongchao Liu, Yan Jiao, Erjian Wang, Stephen H. Clark, Arnold E. Postlethwaite, Weikuan Gu, and Hong Chen. 2015. "Differences between Mice and Humans in Regulation and the Molecular Network of Collagen, Type III, Alpha-1 at the Gene Expression Level: Obstacles that Translational Research Must Overcome" International Journal of Molecular Sciences 16, no. 7: 15031-15056. https://doi.org/10.3390/ijms160715031
APA StyleWang, L., Liu, H., Jiao, Y., Wang, E., Clark, S. H., Postlethwaite, A. E., Gu, W., & Chen, H. (2015). Differences between Mice and Humans in Regulation and the Molecular Network of Collagen, Type III, Alpha-1 at the Gene Expression Level: Obstacles that Translational Research Must Overcome. International Journal of Molecular Sciences, 16(7), 15031-15056. https://doi.org/10.3390/ijms160715031