
Coordinated Actions of Glyoxalase and Antioxidant Defense Systems in Conferring Abiotic Stress Tolerance in Plants

 ,
, 


Abstract
:1. Introduction
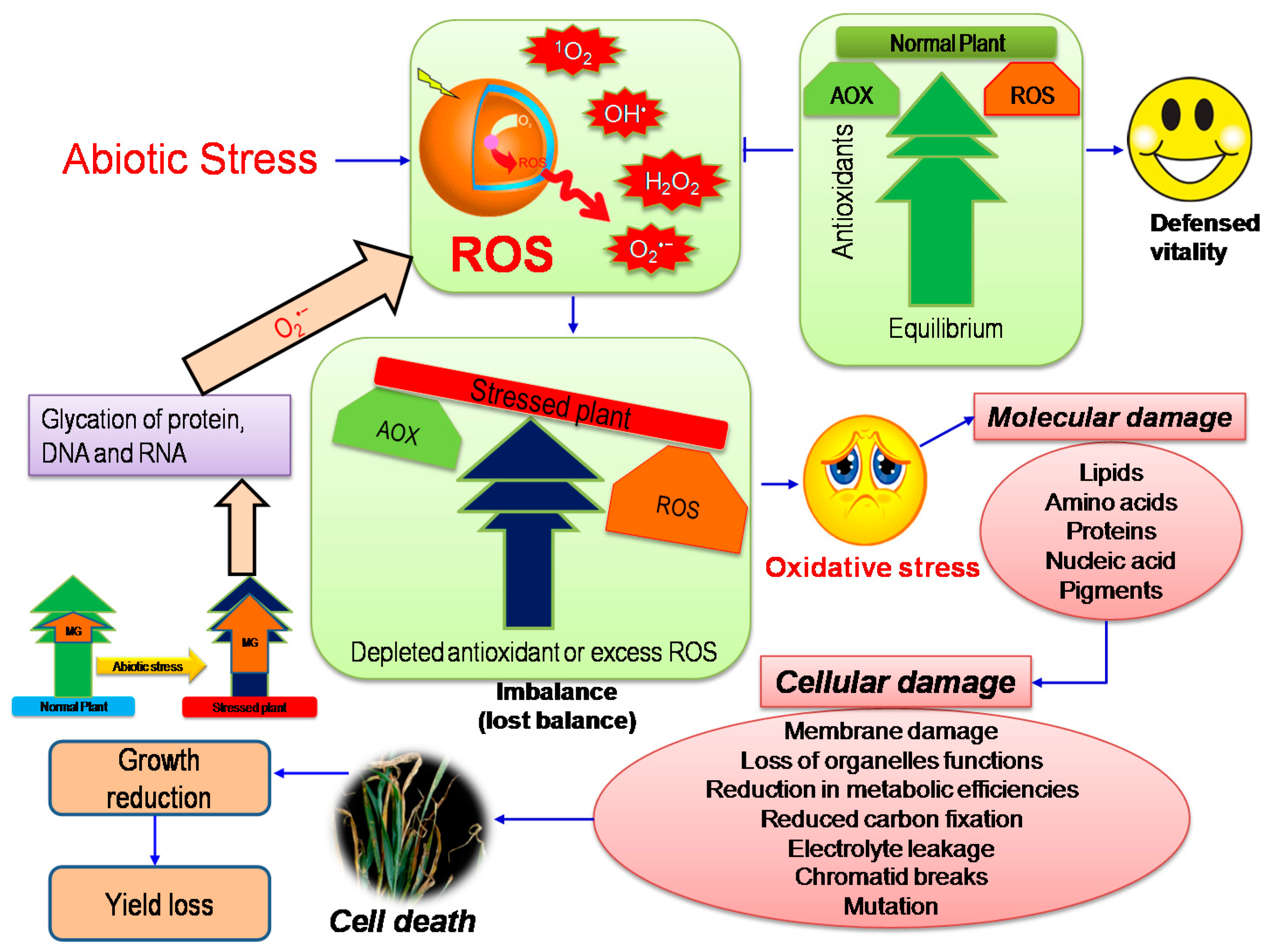
2. Oxidative Stress: A Major Consequence in Plants under Abiotic Stress
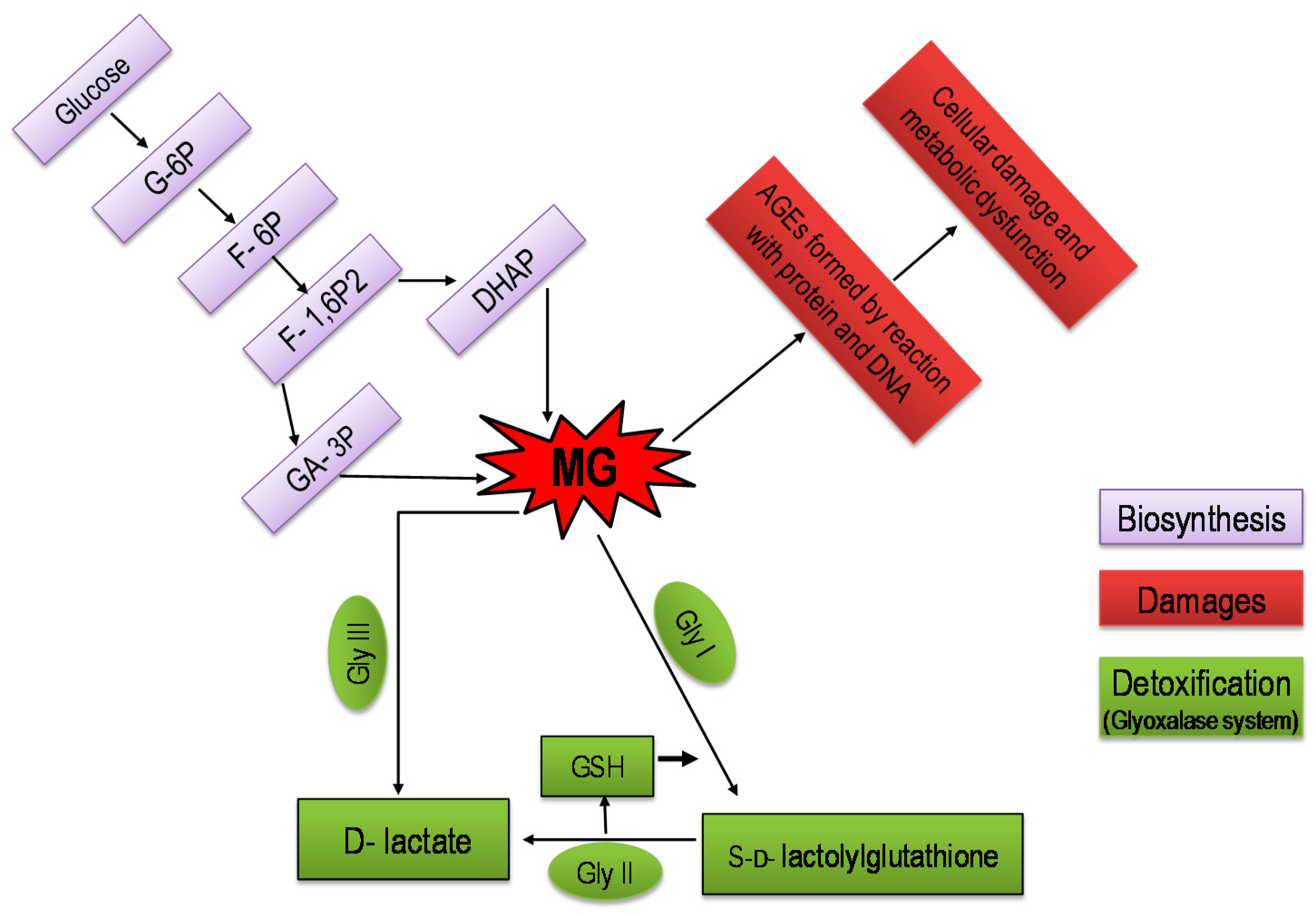
3. Methylglyoxal: An Unavoidable Foe for Plants
4. Methylglyoxal Biosynthesis and Metabolism in Plants
5. Glyoxalase System: The Eliminator of Methylglyoxal in Plants
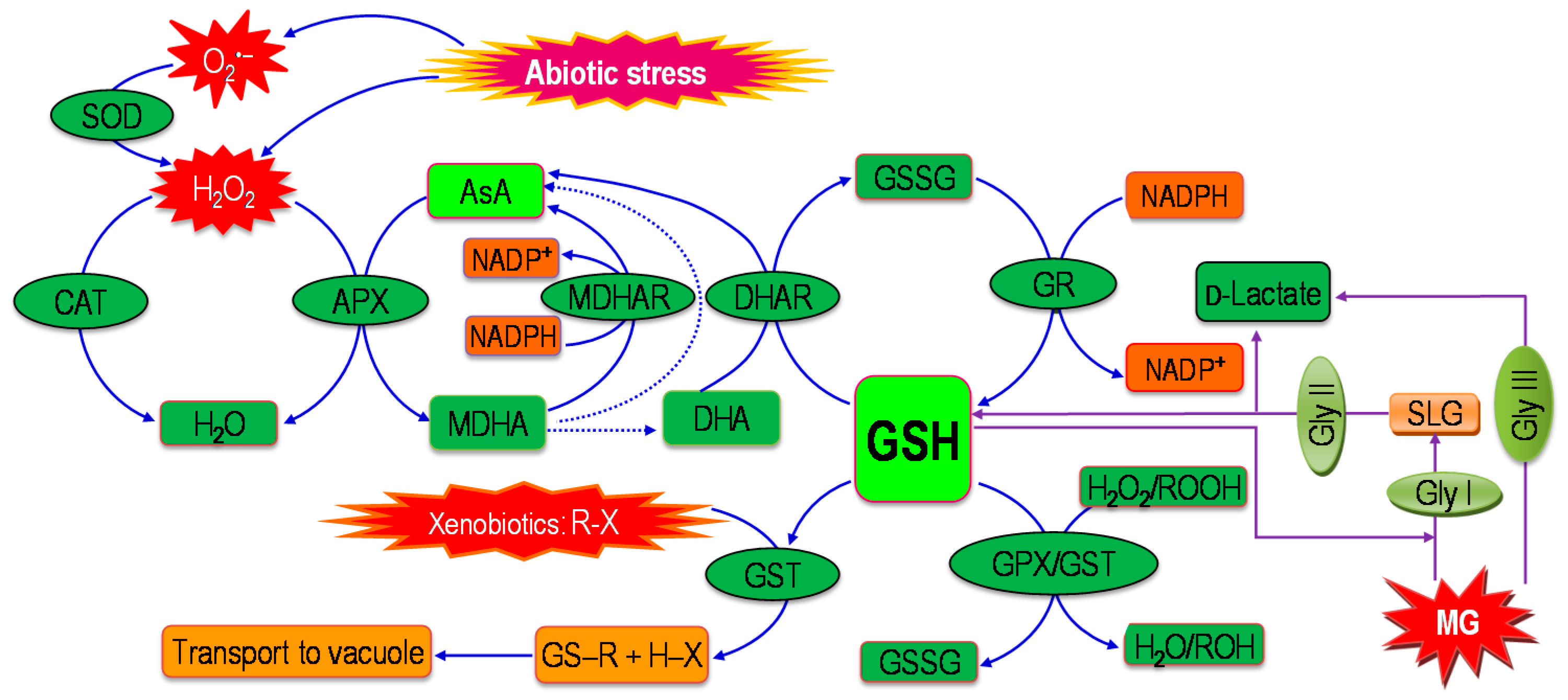
6. Coordinated Actions of Glyoxalase and Antioxidant Defense System in Mitigating Oxidative Stress in Plants
7. Regulation of the Glyoxalase System and the Antioxidant Defense System in Plants under Abiotic Stress
7.1. Salinity
7.2. Drought
7.3. Toxic Metals/Metalloids
7.4. Extreme Temperatures
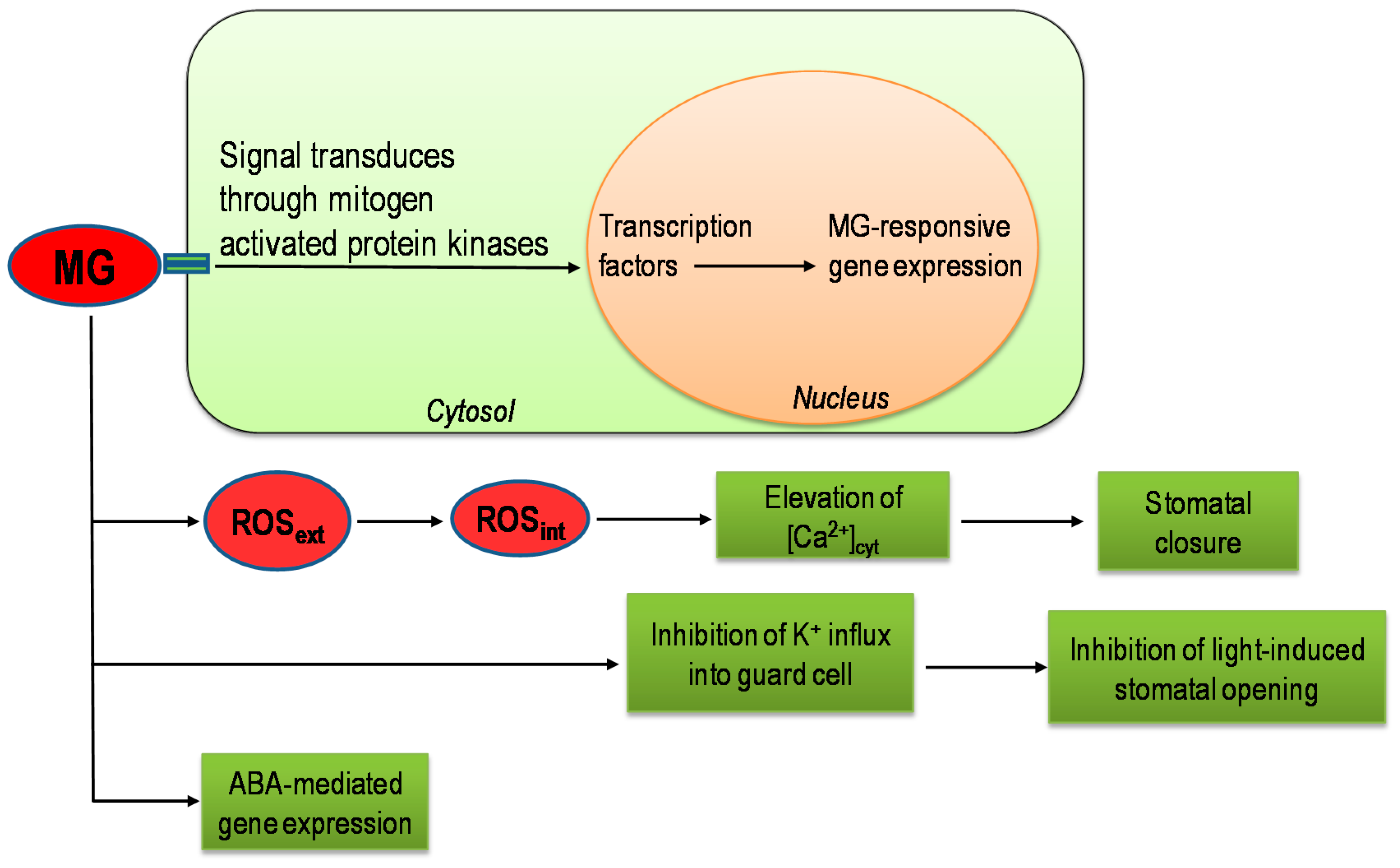
8. Role of Methylglyoxal as a Signaling Molecule
9. Genetic Manipulation in Enhancing Glyoxalase Pathway in Plants
10. Conclusions and Outlook
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AKR | aldo-keto reductase |
| ALR | aldose/aldehyde reductase |
| APX | ascorbate peroxidase |
| AsA | ascorbate |
| chl | chlorophyll |
| CAT | catalase |
| DHA | dehydroascorbic acid |
| DHAP | dihydroxyacetone phosphate |
| DHAR | dehydroascorbate reductase |
| ETC | electron transport chain |
| GAP | glyceraldehyde-3-phosphate |
| Gly | glyoxalase |
| GR | glutathione reductase |
| GSH | reduced glutathione |
| GSSG | oxidized glutathione |
| GPX | glutathione peroxidase |
| GST | glutathione s-transferase |
| HT | high temperature |
| LOX | lipoxygenase |
| LT | low temperature |
| MDHAR | monodehydroascorbate reductase |
| MG | methylglyoxal |
| MGdG | 3-(2′-deoxyribosyl)-6,7-dihydro-6,7-dihydroxy-6/7-methylimidazo-[2,3-b]purine-9(8)one |
| MV | methyl viologen |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| PEG | polyethylene glycol |
| PS II | photosystem II |
| Pro | proline |
| ROS | reactive oxygen species |
| RWC | relative water content |
| SLG | s-d-lactoyl-glutathione |
| SOD | superoxide dismutase |
| SNP | sodium nitroprusside |
| Spd | spermidine |
| Spm | spermine |
| Tre | trehalose |
References
- IPCC. Climate change and water. In Technical Paper of the Intergovernmental Panel for Climate Change; Bates, B.C., Kundzewicz, Z.W., Palutikof, J., Wu, S., Eds.; Secretariat: Geneva, Switzerland, 2008; p. 210. [Google Scholar]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Annu. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Acquaah, G. Principles of Plant Genetics and Breeding; Blackwell: Oxford, UK, 2007; p. 385. [Google Scholar]
- Hasanuzzaman, M.; Hossain, M.A.; Teixeira da Silva, J.A.; Fujita, M. Plant responses and tolerance to abiotic oxidative stress: Antioxidant defense is a key factor. In Crop Stress and its Management: Perspectives and Strategies; Bandi, V., Shanker, A.K., Shanker, C., Mandapaka, M., Eds.; Springer: Berlin, Germany, 2012; pp. 261–316. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Extreme temperatures, oxidative stress and antioxidant defense in plants. In Abiotic Stress—Plant Responses and Applications in Agriculture; Vahdati, K., Leslie, C., Eds.; InTech: Rijeka, Yugoslavia, 2013; pp. 169–205. [Google Scholar]
- Tsukahara, K.; Sawada, H.; Kohno, Y.; Matsuura, T.; Mori, I.C.; Terao, T. Ozone-induced rice grain yield loss is triggered via a change in panicle morphology that is controlled by aberrant panicle organization 1 Gene. PLoS ONE 2015, 10, e0123308. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Singla-Pareek, S.L.; Ray, M.; Reddy, M.K.; Sopory, S.K. Methylglyoxal levels in plants under salinity stress are dependent on glyoxalase I and glutathione. Biochem. Biophys. Res. Commun. 2005, 337, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Chaplen, F.W.R. Incidence and potential implications of the toxic metabolite methylglyoxal in cell culture: A review. Cytotechnology 1998, 26, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium-induced upregulation of the antioxidant defense and methylglyoxal detoxification system reduces salinity-induced damage in rapeseed seedlings. Biol. Trace Elem Res. 2011, 143, 1704–1721. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnol. Rep. 2011, 5, 353–365. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Exogenous nitric oxide alleviates high temperature induced oxidative stress in wheat (Triticum aestivum L.) seedlings by modulating the antioxidant defense and glyoxalase system. Aust. J. Crop Sci. 2012, 6, 1314–1323. [Google Scholar]
- Hasanuzzaman, M.; Fujita, M. Selenium pretreatment upregulates the antioxidant defense and methylglyoxal detoxification system and confers enhanced tolerance to drought stress in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1758–1776. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Fujita, M. Exogenous silicon treatment alleviates salinity-induced damage in Brassica napus L. seedlings by upregulating the antioxidant defense and methylglyoxal detoxification system. In Proceedings of the Annual Meeting of the American Society of Plant Biologists, Minniapolis, MN, USA, 6–10 August 2011.
- Hasanuzzaman, M.; Fujita, M. Exogenous sodium nitroprusside alleviates arsenic-induced oxidative stress in wheat (Triticum aestivum L.) seedlings by enhancing antioxidant defence and glyoxalase system. Ecotoxicology 2013, 22, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Exogenous spermidine alleviates low temperature injury in mung bean (Vigna radiata L.) seedlings by modulating ascorbate-glutathione and glyoxalase pathway. Int. J. Mol. Sci. 2015, 16, 30117–30132. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.A.; Fujita, M. Exogenous glutathione confers high temperature stress tolerance in mung bean (Vigna radiata L.) by modulating antioxidant defense and methylglyoxal detoxification system. Environ. Exp. Bot. 2015, 112, 44–54. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Glutathione-induced drought stress tolerance in mung bean: Coordinated roles of the antioxidant defence and methylglyoxal detoxification systems. AoB Plants 2015, 7, plv069. [Google Scholar] [CrossRef] [PubMed]
- Rajwanshi, R.; Kumar, D.; Yusuf, M.; DebRoy, S.; Sarin, N. Stress-inducible overexpression of glyoxalase I is preferable to its constitutive overexpression for abiotic stress tolerance in transgenic Brassica juncea. Mol. Breed. 2016, 36, 1–15. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Polyamine and nitric oxide crosstalk: Antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense and methylglyoxal detoxification systems. Ecotoxicol. Environ. Saf. 2016, 126, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Mahmud, J.A.; Suzuki, T.; Fujita, M. Insights into spermine-induced combined high temperature and drought tolerance in mung bean: Osmoregulation and roles of antioxidant and glyoxalase system. Protoplasma 2016. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Rahman, M.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Physiological and biochemical mechanisms of spermine-induced cadmium stress tolerance in mung bean (Vigna radiata L.) seedlings. Environ. Sci. Pollut. Res. 2016, 23, 21206–21218. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Rahman, A.; Alam, M.M.; Mahmud, J.A.; Suzuki, T.; Fujita, M. Polyamines confer salt tolerance in mung bean (Vigna radiata L.) by reducing sodium uptake, improving nutrient homeostasis, antioxidant defense, and methylglyoxal detoxification systems. Front. Plant Sci. 2016, 7, 1104. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Kushwaha, H.R.; Hasan, M.R.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Presence of unique glyoxalase III proteins in plants indicates the existence of shorter route for methylglyoxal detoxification. Sci. Rep. 2016, 6, 18358. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Yamamoto, H.; Makino, A.; Sugimoto, T.; Miyake, C. Methylglyoxal functions as hill oxidant and stimulates the photoreductin of O2 at photosystem I: A symptom of plant diabetes. Plant Cell Environ. 2011, 34, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.A.; Uraji, M.; Banu, M.N.A.; Mori, I.C.; Nakamura, Y.; Murata, Y. Methylglyoxal inhibition of cytosolic ascorbate peroxidase from Nicotiana tabacum. J. Biochem. Mol. Toxicol. 2012, 26, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Hoque, T.S.; Uraji, M.; Ye, W.; Hossain, M.A.; Nakamura, Y.; Murata, Y. Methylglyoxal-induced stomatal closure accompanied by peroxidase-mediated ROS production in Arabidopsis. J. Plant Physiol. 2012, 169, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS Function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Bartosz, G. Oxidative stress in plants. Acta Physiol. Plant. 1997, 19, 47–64. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Mol. Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Logan, B. Reactive oxygen species and photosynthesis. In Antioxidants and Reactive Oxygen Species in Plants; Smrinoff, N., Ed.; Blackwell: Oxford, UK, 2005; pp. 250–267. [Google Scholar]
- Kim, K.; Portis, J. Oxygen-dependent H2O2 production by Rubisco. FEBS Lett. 2004, 571, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Ou, W.; Lu, S.; Zhong, Q. Differential responses of antioxidant system to chilling and drought in four rice cultivars differing in sensitivity. Plant Physiol. Biochem. 2006, 44, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Nawata, E.; Hosokawa, M.; Domae, Y.; Sakuratani, T. Alterations in photosynthesis and some antioxidant enzymatic activities of mungbean subjected to waterlogging. Plant Sci. 2002, 163, 117–123. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2008, 14, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Tripathi, R.D.; Dwivedi, U.N. Synthesis of phytochelatins and modulation of antioxidants in response to cadmium stress in Cuscuta reflexa—An angiospermic parasite. J. Plant Physiol. 2004, 161, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Qadir, S.; Qureshi, M.I.; Javed, S.; Abdin, M.Z. Genotypic variation in phytoremediation potential of Brassica juncea cultivars exposed to Cd stress. Plant Sci. 2004, 167, 1171–1181. [Google Scholar] [CrossRef]
- Dong, J.; Wu, F.B.; Zhang, G.P. Influence of cadmium on antioxidant capacity and four microelement concentrations in tomato (Lycopersicon esculentum). Chemosphere 2006, 64, 1659–1666. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Kushwaha, H.R.; Mustafiz, A.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Analysis of global gene expression profile of rice in response to methylglyoxal indicates its possible role as a stress signal molecule. Front. Plant Sci. 2015, 6, 682. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Sharma, S.; Singla-Pareek, S.L.; Sopory, S.K. Methylglyoxal, triose phosphateisomerase and glyoxalase pathway: Implications in abiotic stress and signaling in plants. In Elucidation of Abiotic Stress Signaling in Plants; Pandey, G.K., Ed.; Springer: New York, NY, USA, 2015; pp. 347–366. [Google Scholar]
- Yadav, S.K.; Singla-Pareek, S.L.; Ray, M.; Reddy, M.K.; Sopory, S.K. Transgenic tobacco plants overexpressing glyoxalase enzymes resist an increase in methylglyoxal and maintain higher reduced glutathione levels under salinity stress. FEBS Lett. 2005, 579, 6265–6271. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Singla-Pareek, S.L.; Sopory, S.K. An overview on the role of methylglyoxal and glyoxalases in plants. Drug Metabol. Drug Interact. 2008, 23, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J. Glutathione-dependent detoxification of α-oxoaldehydesby the glyoxalase system: Involvement in disease mechanisms and antiproliferative activity of glyoxalase I inhibitors. Chem. Biol. Interact. 1998, 111–112, 137–151. [Google Scholar] [CrossRef]
- Thornalley, P.J. Protein and nucleotide damage by glyoxal and methylglyoxal in physiological systems—Role in ageing and disease. Drug Metabol. Drug Interact. 2008, 23, 125–150. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Thornalley, P.J. Methylglyoxal, glyoxalase 1 and the dicarbonyl proteome. Amino Acids 2012, 42, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Thornalley, P.J. Dicarbonyl proteome and genome damage in metabolic and vascular disease. Biochem. Soc. Trans. 2014, 42, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Singla-Pareek, S.L.; Sopory, S.K. Glyoxalase and methylglyoxal as biomarkers for plant stress tolerance. Crit. Rev. Plant Sci. 2014, 33, 429–456. [Google Scholar] [CrossRef]
- Hoque, T.S.; Hossain, M.A.; Mostofa, M.G.; Burritt, D.J.; Fujita, M.; Tran, L.S.P. Methylglyoxal: An emerging signaling molecule in plant abiotic stress responses and tolerance. Front. Plant Sci. 2016, 7, 1341. [Google Scholar] [CrossRef] [PubMed]
- Mankikar, S.; Rangekar, P. Effects of methylglyoxal on germination of barley. Fyton 1974, 32, 9–16. [Google Scholar]
- Engqvist, M.K.M.; Drincovich, M.F.; Flügge, U.I.; Maurino, V.G. Two d-2-hydroxy-acid dehydrogenases in Arabidopsis thaliana with catalytic capacities to participate in the last reactions of themethylglyoxal and β-oxidation pathways. J. Biol. Chem. 2009, 284, 25026–25037. [Google Scholar] [CrossRef] [PubMed]
- Wienstroer, J.; Engqvist, M.K.M.; Kunz, H.H.; Flügge, U.I.; Maurino, V.G. d-Lactate dehydrogenase as a marker gene allows positive selection oftransgenic plants. FEBS Lett. 2012, 586, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Mano, J.; Miyatake, F.; Hiraoka, E.; Tamoi, M. Evaluation of the toxicity of stress-related aldehydes to photosynthesis in chloroplasts. Planta 2009, 230, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Fukuda, Y.; Watanabe, K.; Saikusa, T.; Shimosaka, M.; Kimura, A. Characterization of methylglyoxal synthase in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 1985, 131, 190–198. [Google Scholar] [CrossRef]
- Richard, J.P. Mechanism for the formation of methylglyoxal from triosephosphates. Biochem. Soc. Trans. 1993, 21, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Kalapos, M.P. Methylglyoxal in living organisms: Chemistry, biochemistry, toxicology and biological implications. Toxicol. Lett. 1999, 110, 145–175. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hossain, M.Z.; Fujita, M. Stress-induced changes of methylglyoxal level and glyoxalase I activity in pumpkin seedlings and cDNA cloning of glyoxalase I gene. Aust. J. Crop Sci. 2009, 3, 53–64. [Google Scholar]
- Hegedüs, A.; Erdei, S.; Janda, T.; Tóth, E.; Horváth, G.; Dudits, D. Transgenictobacco plants overproducing alfalfa aldose/aldehydes reductase show highertolerance to low temperature and cadmium stress. Plant Sci. 2004, 166, 1329–1333. [Google Scholar] [CrossRef]
- Turόczy, Z.; Kis, P.; Török, K.; Cserháti, M.; Lendvai, A.; Dudits, D.; Horváth, G.V. Overproduction of a rice aldo-keto reductase increases oxidative and heat stress tolerance by malondialdehyde and methylglyoxal detoxification. Plant Mol. Biol. 2011, 75, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Teixeira da Silva, J.A.; Fujita, M. Glyoxalase system and reactive oxygen species detoxification system in plant abiotic stress response and tolerance: An intimate relationship. In Abiotic Stress in Plants—Mechanisms and Adaptations; Shanker, A.K., Ed.; Intech: Rejika, Yugoslavia, 2011; pp. 235–266. [Google Scholar]
- Neuberg, C. The destruction of lactic aldehyde and methylglyoxal by animal organs. Biochem. J. 1913, 49, 502–506. [Google Scholar]
- Dakin, H.D.; Dudley, H.W. An enzyme concerned with the formation of hydroxyl acids from ketonic aldehydes. J. Biol. Chem. 1913, 14, 155–157. [Google Scholar]
- Norton, S.J.; Talesa, V.; Yuan, W.J.; Principato, G.B. Glyoxalase I and Glyoxalase II from Aloevera: Purification, characterization and comparison with animal glyoxalases. Biochem. Int. 1990, 22, 411–418. [Google Scholar] [PubMed]
- Maiti, M.K.; Krishnasamy, S.; Owen, H.A.; Makaroff, C.A. Molecular characterization of glyoxalase II from Arabidopsis thaliana. Plant Mol. Biol. 1997, 35, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Racker, E. The mechanism of action of glyoxalase. J. Biol. Chem. 1951, 190, 685–696. [Google Scholar] [PubMed]
- Reddy, V.S.; Sopory, S.K. Glyoxalase I from Brassica juncea: Molecular cloning, regulation and its over-expression confer tolerance in transgenic tobacco under stress. Plant J. 1999, 17, 385–395. [Google Scholar] [CrossRef]
- Singla-Pareek, S.L.; Yadav, S.K.; Pareek, A.; Reddy, M.K.; Sopory, S.K. Enhancing salt tolerance in a crop plant by overexpression of glyoxalase II. Transgenic Res. 2008, 17, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Singla-Pareek, S.L.; Reddy, M.K.; Sopory, S.K. Genetic engineering of the glyoxalase pathway in tobacco leads to enhanced salinity tolerance. Proc. Natl. Acad. Sci. USA 2003, 100, 14672–14677. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Mostofa, M.G.; Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium mitigates arsenic toxicity in rice seedlings by reducing arsenic uptake and modulating the antioxidant defense and glyoxalase systems and stress markers. BioMed Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Simpson, P.J.; Tantitadapitak, C.; Reed, A.M.; Mather, O.C.; Bunce, C.M.; White, S.A.; Ride, J.P. Characterization of two novel aldo-keto reductases from Arabidopsis: Expression patterns, broad substratespecificity, and an open active-site structure suggest a role in toxicant metabolism following stress. J. Mol. Biol. 2009, 392, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Hasegawa, A.; Taninaka, A.; Mizutani, M.; Sugimoto, Y. NADPH-dependent reductases involved in the detoxification of reactive carbonyls in plants. J. Biol. Chem. 2011, 286, 6999–7009. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Singh, P.; Yusuf, M.A.; Upadhyaya, C.P.; Roy, S.D.; Hohn, T.; Sarin, N.B. The Xerophytaviscosa aldose reductase (ALDRXV4) confers enhanced drought and salinity tolerance to transgenic tobacco plants by scavenging methylglyoxal and reducing the membrane damage. Mol. Biotechnol. 2013, 54, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Narawongsanont, R.; Kabinpong, S.; Auiyawong, B.; Tantitadapitak, C. Cloning and characterization of AKR4C14, a rice aldo-keto reductase, from Thai Jasmine rice. Protein J. 2012, 31, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.A.; Uraji, M.; Banu, M.N.A.; Mori, I.C.; Nakamura, Y.; Murata, Y. The effects of methylglyoxal on glutathione S-transferase from Nicotiana tabacum. Biosci. Biotechnol. Biochem. 2010, 74, 2124–2126. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Ghosh, A.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Glyoxalases and stress tolerance in plants. Biochem. Soc. Trans. 2014, 42, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Alam, M.M.; Nahar, K.; Al-Mahmud, J.; Ahamed, K.U.; Fujita, M. Exogenous salicylic acid alleviates salt stress-induced oxidative damage in Brassica napus by enhancing the antioxidant defense and glyoxalase systems. Aust. J. Crop Sci. 2014, 8, 631–639. [Google Scholar]
- Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous proline and glycine betaine mediated upregulation of antioxidant defense and glyoxalase systems provides better protection against salt-induced oxidative stress in two rice (Oryza sativa L.) varieties. BioMed Res. Int. 2014. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Saegusa, D.; Fujita, M.; Tran, L.P. Hydrogen sulfide regulates salt tolerance in rice by maintaining Na+/K+ balance, mineral homeostasis and oxidative metabolism under excessive salt stress. Front. Plant Sci. 2015, 6, 1055. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Hossain, M.A.; Fujita, M.; Tran, L.S. Physiological and biochemical mechanisms associated with trehalose-induced copper-stress tolerance in rice. Sci. Rep. 2015, 5, 11433. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium supplementation improves Na+/K+ ratio, antioxidant defense and glyoxalase systems in salt-stressed rice seedlings. Front. Plant Sci. 2016, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Hossain, M.S.; Mahmud, J.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Manganese-induced salt stress tolerance in rice seedlings: Regulation of ion homeostasis, antioxidant defense and glyoxalase systems. Physiol. Mol. Biol. Plants 2016. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Singla-Pareek, S.L.; Reddy, M.K.; Sopory, S.K. Methylglyoxal detoxification by glyoxalase system: A survival strategy during environmental stresses. Physiol. Mol. Biol. Plants 2005, 11, 1–11. [Google Scholar]
- Hossain, M.A.; Fujita, M. Evidence for a role of exogenous glycinebetaine and proline in antioxidant defense and methylglyoxal detoxification systems in mung bean seedlings under salt stress. Physiol. Mol. Biol. Plants 2010, 16, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Mostofa, M.G.; Fujita, M. Heat-shock positively modulates oxidative protection of salt and drought-stressed mustard (Brassica campestris L.) seedlings. J. Plant Sci. Mol. Breed. 2013. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Hossain, M.A.; Fujita, M. Trehalose pretreatment induces salt tolerance in rice (Oryza sativa L.) seedlings: Oxidative damage and co-induction of antioxidantdefense and glyoxalase systems. Protoplasma 2015, 252, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Fujita, M. Drought stress responses in plants, oxidative stress, and antioxidant defense. In Climate Change and Plant Abiotic Stress Tolerance; Tuteja, N., Gill, S.S., Eds.; Wiley: Weinheim, Germany, 2014; pp. 209–250. [Google Scholar]
- Espartero, J.; Sanchez-Aguayo, I.; Pardo, J.M. Molecular characterization of glyoxalase-I from a higher plant; upregulation by stress. Plant Mol. Biol. 1995, 29, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Singla-Pareek, S.L.; Yadav, S.K.; Pareek, A.; Reddy, M.K.; Sopory, S.K. Transgenic tobacco overexpressing glyoxalase pathway enzymes grow and set viable seeds in zinc-spiked soils. Plant Physiol. 2006, 140, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Hideg, É.; Nagy, T.; Oberschall, A.; Dudits, D.; Vass, I. Detoxification function of aldose/aldehyde reductase during drought and ultraviolet-B (280–320 nm) stresses. Plant Cell Environ. 2003, 26, 513–522. [Google Scholar] [CrossRef]
- Oberschall, A.; Deák, M.; Török, K.; Sass, L.; Vass, I.; Kovács, I.; Fehér, A.; Dudits, D.; Horváth, G.V. A novel aldose/aldehyde reductase protects transgenic plants against lipid peroxidation under chemical and drought stress. Plant J. 2000, 24, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Mostofa, M.G.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Exogenous calcium alleviates cadmium-induced oxidative stress in rice (Oryza sativa L.) seedlings by regulating the antioxidant defense and glyoxalase systems. Braz. J. Bot. 2016, 39, 393–407. [Google Scholar]
- Rahman, A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Manganese-induced cadmium stress tolerance in rice seedlings: Coordinated action of antioxidant defense, glyoxalase system and nutrient homeostasis. C. R. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Hasanuzzaman, M.; Fujita, M. Upregulation of antioxidant and glyoxalase systems by exogenous glycinebetaine and proline in mung bean confer tolerance to cadmium stress. Physiol. Mol. Biol. Plants 2010, 16, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Exogenous selenium pretreatment protects rapeseed seedlings from cadmium-induced oxidative stress by upregulating the antioxidant defense and methylglyoxal detoxification systems. Biol. Trace Elem. Res. 2012, 149, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Fujita, M. Salicylic acid alleviates copper toxicity in rice (Oryza sativa L.) seedlings by upregulating antioxidative and glyoxalase systems. Ecotoxicology 2013, 22, 959–973. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Seraj, Z.I.; Fujita, M. Exogenous sodium nitroprusside and glutathione alleviate copper toxicity by reducing copper uptake and oxidative damage in rice (Oryza sativa L.) seedlings. Protoplasma 2014, 251, 1373–1386. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Wollenweber, B.; Jiang, D. Multiple heat priming enhances thermo-tolerance to a later high temperature stress via improving subcellular antioxidant activities in wheat seedlings. Plant Physiol.Biochem. 2014, 74, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Li, X.Q.; Wang, G.G.; Zhu, X.T. Brassinosteroids alleviate high-temperature injury in Ficusconcinna seedlings via maintaining higher antioxidant defence and glyoxalase systems. AoB Plants 2015, 7, plv009. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Singla-Pareek, S.L.; Kumar, M.; Pareek, A.; Saxena, M.; Sarin, N.B.; Sopory, S.K. Characterization and functional validation of glyoxalase II from rice. Protein Exp. Purif. 2007, 51, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Snedden, W.A.; Fromm, H. Calmodulin, calmodulin-related proteins and plant responses to the environment. Trends Plant Sci. 1998, 3, 299–304. [Google Scholar] [CrossRef]
- Hoque, T.S.; Uraji, M.; Tuya, A.; Nakamura, Y.; Murata, Y. Methylglyoxal inhibits seed germination and root elongation and upregulates transcription of stress-responsive genes in ABA-dependent pathway in Arabidopsis. Plant Biol. 2012, 14, 854–858. [Google Scholar] [CrossRef] [PubMed]
- Devanathan, S.; Erban, A.; Rodolfo, P.J.; Kopka, J.; Makaroff, C.A. Arabidopsis thaliana glyoxalase 2-1 is required during abiotic stress but is not essential under normal plant growth. PLoS ONE 2014, 9, e95971. [Google Scholar] [CrossRef] [PubMed]
- Bhomkar, P.; Upadhyay, C.P.; Saxena, M.; Muthusamy, A.; Prakash, N.S.; Pooggin, M.; Hohn, T.; Sarin, B. Salt stress alleviation in transgenic Vigna mungo L. Hepper (blackgram) by overexpression of the glyoxalase I gene using a novel Cestrum yellow leaf curling virus (CmYLCV) promoter. Mol. Breed. 2008, 22, 169–181. [Google Scholar] [CrossRef]
- Tuomainen, M.; Ahonen, V.; Kärenlampi, S.O.; Schat, H.; Paasela, T.; Svanys, A.; Tuohimetsä, S.; Peraniemi, S.; Tervahauta, A. Characterization of the glyoxalase 1 gene TcGLX1 in the metal hyperaccumulator plant Thlaspi caerulescens. Planta 2011, 233, 1173–1184. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Xu, J.; Shi, J.; Li, H.; Li, B. Molecular cloning and characterization of a novel glyoxalase I gene TaGLY I in wheat (Triticum aestivum L.). Mol. Biol. Rep. 2010, 37, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Gerding, X.; Cortés-Bullemore, R.; Medina, C.; Romero-Romero, J.L.; Inostroza-Blancheteau, C.; Aquea, F.; Arce-Johnson, P. Improved salinity tolerance in carrizo citrange rootstock through overexpression of glyoxalase system genes. BioMed Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Ma, C.; Pan, Y.; Gong, S.; Zhao, C.; Chen, S.; Li, H. Sugar beet M14 glyoxalase I gene can enhance plant tolerance to abiotic stresses. J. Plant Res. 2013, 126, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; Bisht, R.; Roy, S.D.; Sopory, S.K.; Bhalla-Sarin, N. Cloning and characterization of a mitochondrial glyoxalase II from Brassica juncea that is upregulated by NaCl, Zn, and ABA. Biochem. Biophys. Res. Commun. 2005, 336, 813–819. [Google Scholar]
- Mustafiz, A.; Singh, A.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Genome-wide analysis of rice and Arabidopsis identifies two glyoxalase genes that are highly expressed in abiotic stresses. Funct. Integr. Genom. 2011, 11, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Mustafiz, A.; Ghosh, A.; Tripathi, A.K.; Kaur, C.; Ganguly, A.K.; Bhavesh, N.S.; Tripathi, J.K.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. A unique Ni2+-dependent and methylglyoxal-inducible rice glyoxalase I possesses a single active site and functions in abiotic stress response. Plant J. 2014, 78, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, T.N.; Sopory, S.K. Light stimulated cell proliferation and glyoxalase-I activity in callus cultures of Amaranthus paniculatus. In Progress in Plant Cellular and Molecular Biology; Nijkamp, H.J.J., van der Plas, L.H.W., Aartrijk, V.J., Eds.; Kluwer Academic: Dordrecht, The Netherland, 1990; pp. 379–384. [Google Scholar]
- Chakravarty, T.N.; Sopory, S.K. Blue light stimulation of cell proliferation and glyoxalase I activity in callus cultures of Amaranthus paniculatus. Plant Sci. 1998, 132, 63–69. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Stress (Dose and Duration) | Changes in Glyoxalase and Antioxidant Defense Systems | Protectants | Changes in Glyoxalase and Antioxidant Defense Systems after Protection | Reference |
|---|---|---|---|---|---|
| V. radiata | 300 mM NaCl, 24 and 48 h |
| 15 mM Pro, 15 mM GB |
| [83] |
| B. campestris | 150 mM NaCl, 48 h |
| Heat shock (42 °C), 5 h |
| [84] |
| B. napus | 100 and 200 mM NaCl, 48 h |
| 100 μM SA |
| [76] |
| T. aestivum | 150 and 300 mM NaCl, 4 days |
| 1 mM sodium nitroprusside (SNP), 24 h pretreatment |
| [12] |
| O. sativa | 150 and 300 mM NaCl |
| 5 mM Pro and 5 mM GB |
| [77] |
| O. sativa | 150 and 250 mM NaCl, 72 h |
| 10 mM Trehalose (Tre) |
| [85] |
| O. sativa | 150 mM NaCl, 4 days |
| 50 μM H2S |
| [78] |
| O. sativa | 200 mM NaCl, 3 days |
| 2 mM CaCl2 |
| [80] |
| O. sativa | 150 mM NaCl, 3 and 6 days |
| 0.5 mM MnSO4 |
| [81] |
| Plant Species | Stress (Dose and Duration) | Changes in Glyoxalase and Antioxidant Defense Systems | Protectants | Changes in Glyoxalase and Antioxidant Defense Systems after Protection | Reference |
|---|---|---|---|---|---|
| V. radiata | 5% PEG-6000, 48 h |
| 0.2 mM Spermidine (Spd) |
| [22] |
| V. radiata | 25% PEG-6000, 24 and 48 h |
| 1 mM GSH |
| [19] |
| B. juncea | Water deficit stress |
| - | - | [66] |
| S. lycopersicum | Water deficit stress created by mannitol |
| - | - | [87] |
| B. napus | 10% and 20% PEG-6000, 48 h |
| 25 μM Na2SeO4 |
| [14] |
| O. sativa | Desiccation, 8 h |
| - | - | [25] |
| Plant Species | Stress (Dose and Duration) | Changes in Glyoxalase and Antioxidant Defense Systems | Protectants | Changes in Glyoxalase and Antioxidant Defense Systems after Protection | Reference |
|---|---|---|---|---|---|
| V. radiata | 1 mM CdCl2, 48 h |
| 5 mM Pro or GB, 48 h |
| [93] |
| B. napus | 0.5 and 1.0 mM CdCl2, 48 h |
| Seed pretreatment; 50 and 100 μM Na2SeO4, 24 h |
| [94] |
| T. aestivum | 0.25 and 0.5 mM Na2HAsO4·7H2O, 72 h |
| 0.25 mM SNP, 72 h |
| [16] |
| O. sativa | 150 µM CuSO4, 48 h |
| Pretratment, 100 µM SA, 24 h |
| [95] |
| O. sativa | 100 μM CuSO4, 48 h |
| 200 μM SNP or 200 μM GSH, 48 h |
| [96] |
| O. sativa | 100 µM CuSO4, 4 and 7 days |
| Pretreatment, 10 mM Tre, 48 h |
| [79] |
| O. sativa | 0.5 and 1 mM Na2HAsO4, 5 days |
| 10 mM CaCl2 (Ca), 5 days |
| [69] |
| O. sativa | 0.25 and 0.5 mM CdCl2, 72 h |
| 2.5 mM CaCl2, 72 h |
| [91] |
| O. sativa | 0.3 mM CdCl2, 72 h |
| 0.3 mM MnSO4, 72 h |
| [92] |
| V. radiata | 1.5 mM CdCl2, 48 h |
| Pretreatment, 0.2 mM Put and 1 mM SNP, 24 h |
| [21] |
| V. radiata | CdCl2, 1.0 and 1.5 mM |
| Pretreatment, 0.25 mM Spm, 24 h |
| [23] |
| Plant Species | Extent of Temperature Stress | Changes in Glyoxalase and Antioxidant Defense Systems | Protectants | Changes in Glyoxalase and Antioxidant Defense Systems after Protection | Reference |
|---|---|---|---|---|---|
| F. concinna | 35 and 40 °C, 48 h |
| 0.25 µM 24-epibrassinolide (EBR) |
| [98] |
| O. sativa | 45 °C, 15–120 min |
| - | - | [99] |
| O. sativa | 42 °C, 8 h |
| - | - | [25] |
| V. radiata | 40 °C, 2 days |
| Spermine (Spm, 0.2 mM) |
| [22] |
| T. aestivum | 38 °C, 24 and 48 h |
| 0.5 mM SNP |
| [13] |
| V. radiata | 6 °C, 2 and 3 days |
| 0.25 mM Spd |
| [17] |
| O. sativa | 4 °C, 8 h |
| - | - | [25] |
| O. sativa | 4 °C, 15 min–2 h |
| - | - | [99] |
| Transgenic Plant | Gene | Gene Sources | Tolerance Response in Transgenic Plant | References |
|---|---|---|---|---|
| Beta corolliflora × B. vulgaris | BvM14-glyoxalase I | B. corolliflora and B. vulgaris |
| [107] |
| N. tabacum | TaGly I | T. aestivum |
| [105] |
| N. tabacum | TcGLX1 | B. juncea |
| [104] |
| N. tabacum | Gly I cDNA | B. juncea |
| [66] |
| O. sativa | OsDJ-1C | Arabidopsis sp. |
| [25] |
| O. sativa | OsDJ-1C | A. thaliana |
| [25] |
| C. Citrange | BjGlyI and PgGlyII | BjGlyI from B. juncea and PgGlyII from Pennisetum glaucum |
| [106] |
| B. juncea | Bj glyII | P. glaucum, O. sativa, Arabidopsis, and C. arietinum |
| [108] |
| O. sativa | OsglyII | O. sativa |
| [67] |
| N. tabaccum | gly I and gly II | gly I from B. juncea, gly II gene isolated from O. sativa |
| [68] |
| N. tabacum | gly I and gly II | gly I from B. juncea, gly II gene isolated from O. sativa |
| [88] |
| N. tabacum | OsGLYI-11.2 | O. sativa |
| [110] |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasanuzzaman, M.; Nahar, K.; Hossain, M.S.; Mahmud, J.A.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Coordinated Actions of Glyoxalase and Antioxidant Defense Systems in Conferring Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2017, 18, 200. https://doi.org/10.3390/ijms18010200
Hasanuzzaman M, Nahar K, Hossain MS, Mahmud JA, Rahman A, Inafuku M, Oku H, Fujita M. Coordinated Actions of Glyoxalase and Antioxidant Defense Systems in Conferring Abiotic Stress Tolerance in Plants. International Journal of Molecular Sciences. 2017; 18(1):200. https://doi.org/10.3390/ijms18010200
Chicago/Turabian StyleHasanuzzaman, Mirza, Kamrun Nahar, Md. Shahadat Hossain, Jubayer Al Mahmud, Anisur Rahman, Masashi Inafuku, Hirosuke Oku, and Masayuki Fujita. 2017. "Coordinated Actions of Glyoxalase and Antioxidant Defense Systems in Conferring Abiotic Stress Tolerance in Plants" International Journal of Molecular Sciences 18, no. 1: 200. https://doi.org/10.3390/ijms18010200
APA StyleHasanuzzaman, M., Nahar, K., Hossain, M. S., Mahmud, J. A., Rahman, A., Inafuku, M., Oku, H., & Fujita, M. (2017). Coordinated Actions of Glyoxalase and Antioxidant Defense Systems in Conferring Abiotic Stress Tolerance in Plants. International Journal of Molecular Sciences, 18(1), 200. https://doi.org/10.3390/ijms18010200