Lactobacillus bulgaricus, Lactobacillus rhamnosus and Lactobacillus paracasei Attenuate Salmonella Enteritidis, Salmonella Heidelberg and Salmonella Typhimurium Colonization and Virulence Gene Expression In Vitro



Abstract
:1. Introduction
2. Results and Discussion
2.1. Sub-Inhibitory Concentrations (SICs) of LAB Cell-Free Supernatant (CFSN)
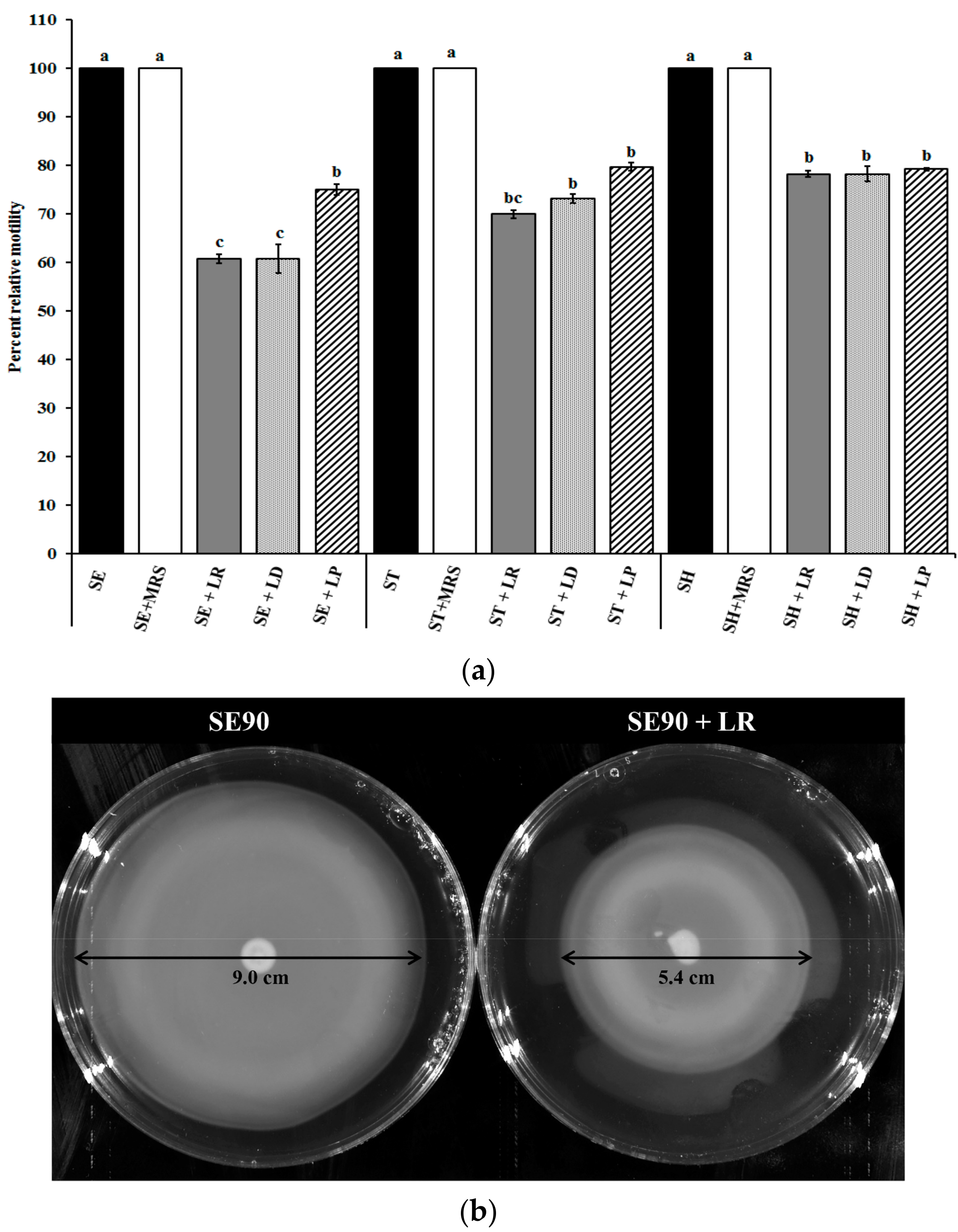
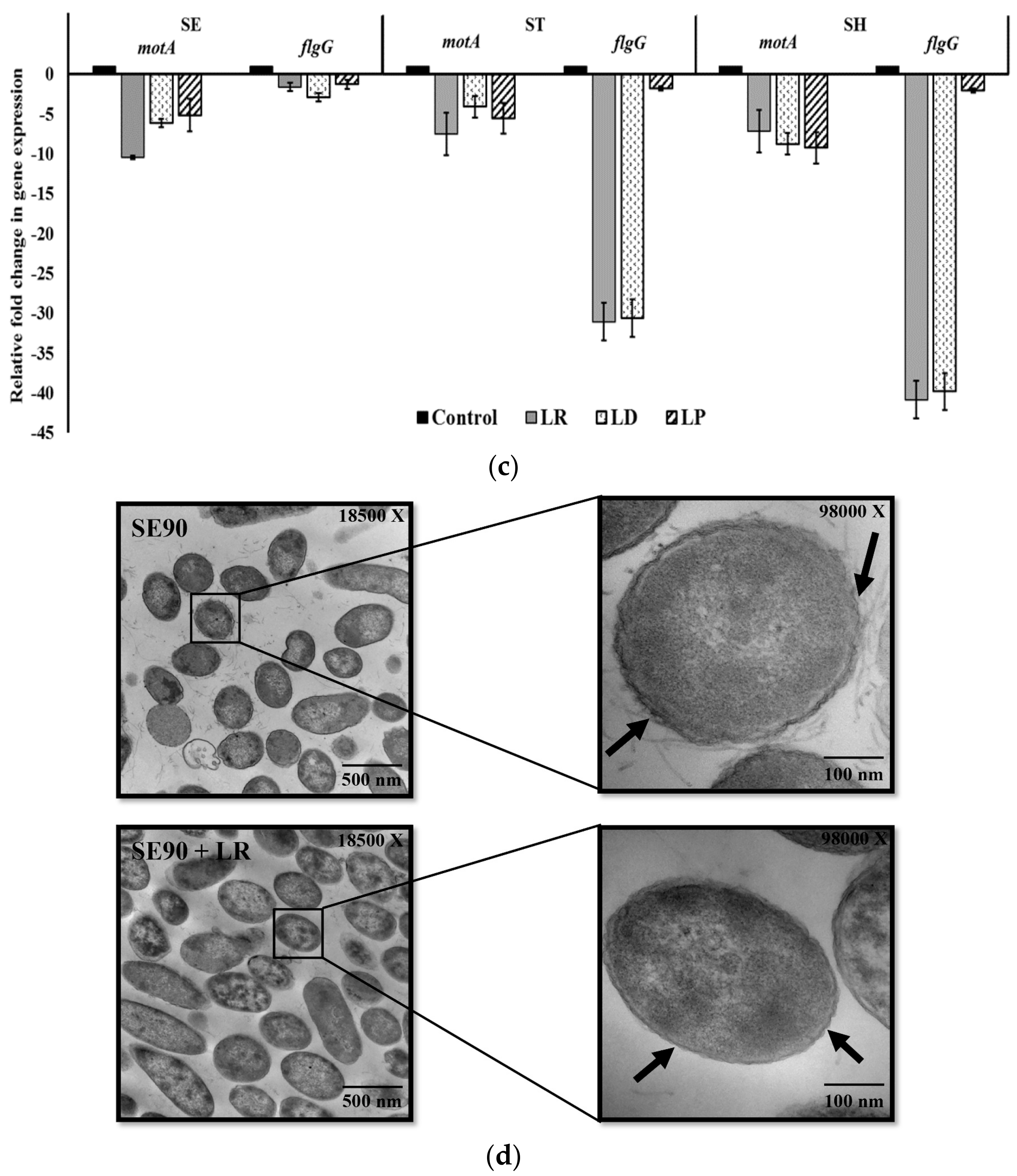
2.2. Motility Assay
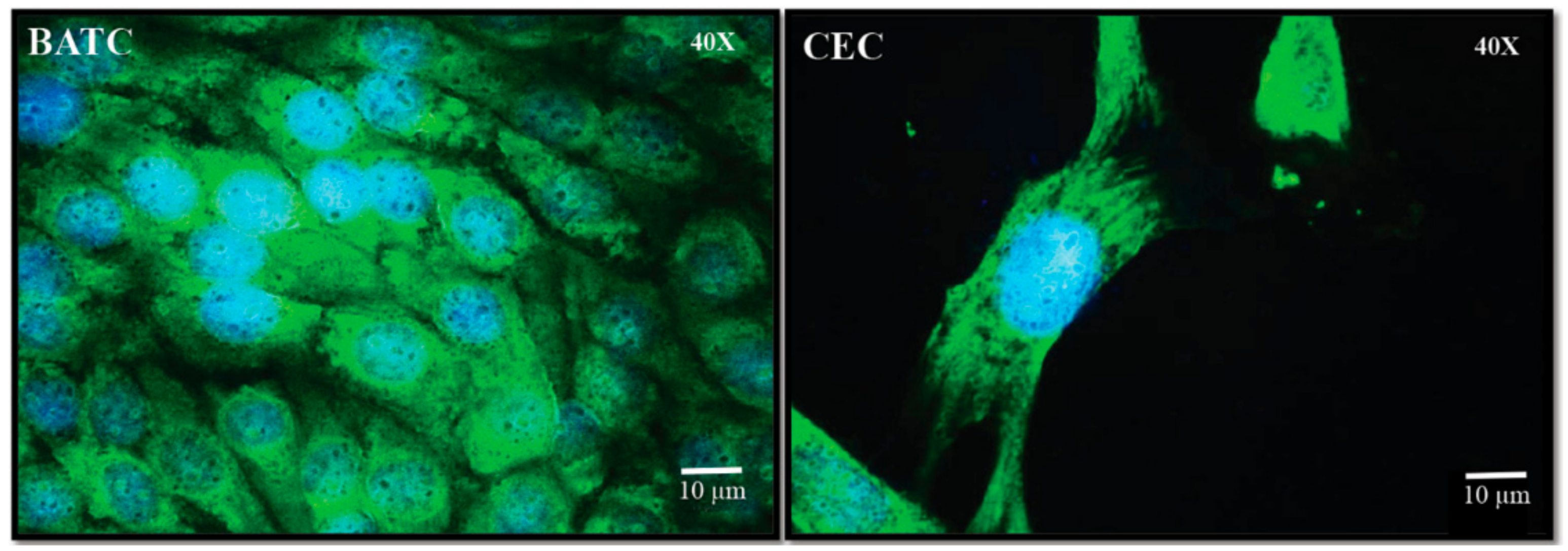
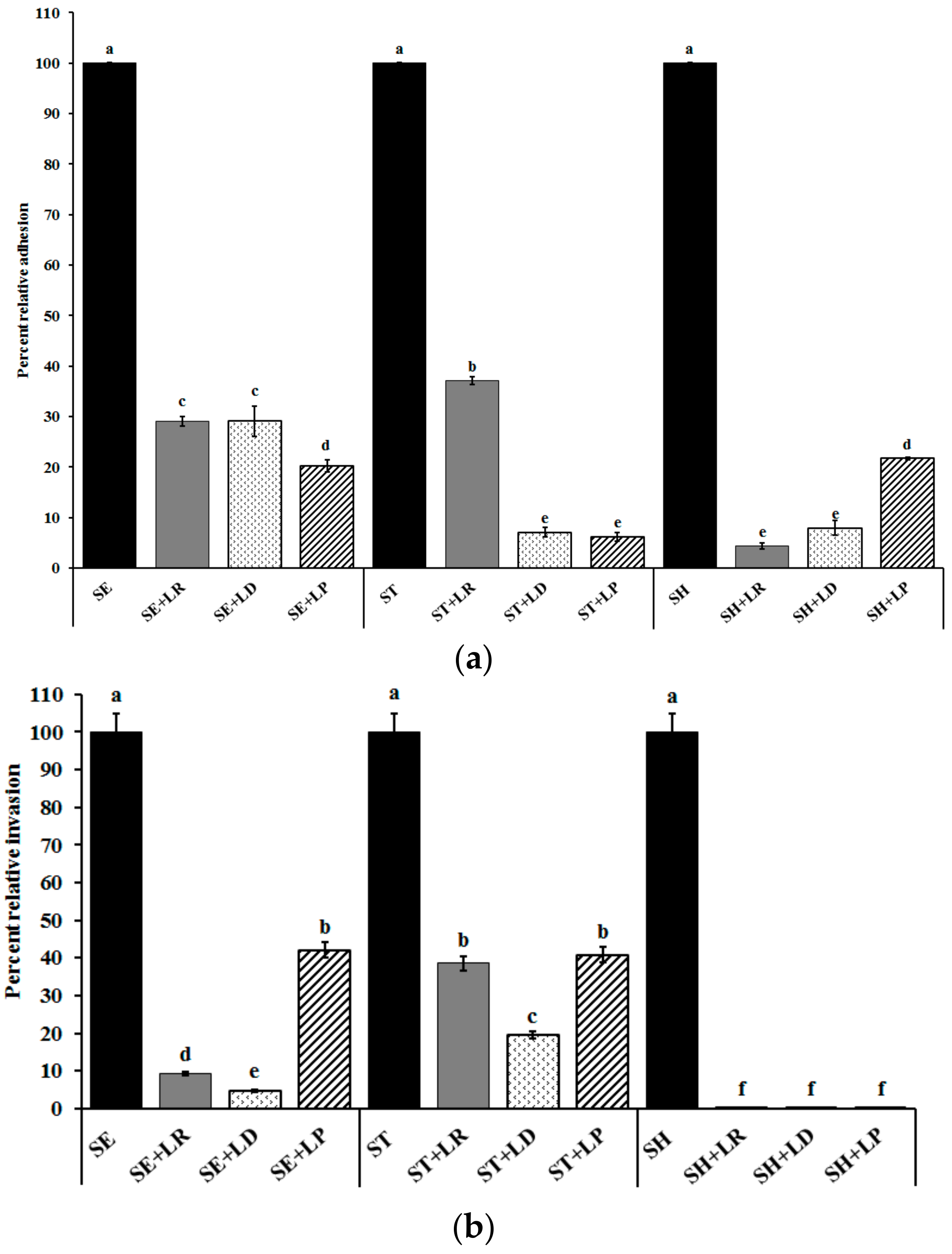
2.3. Inhibition of Salmonella Adhesion and Invasion in Cecal Epithelial Cells (CEC)
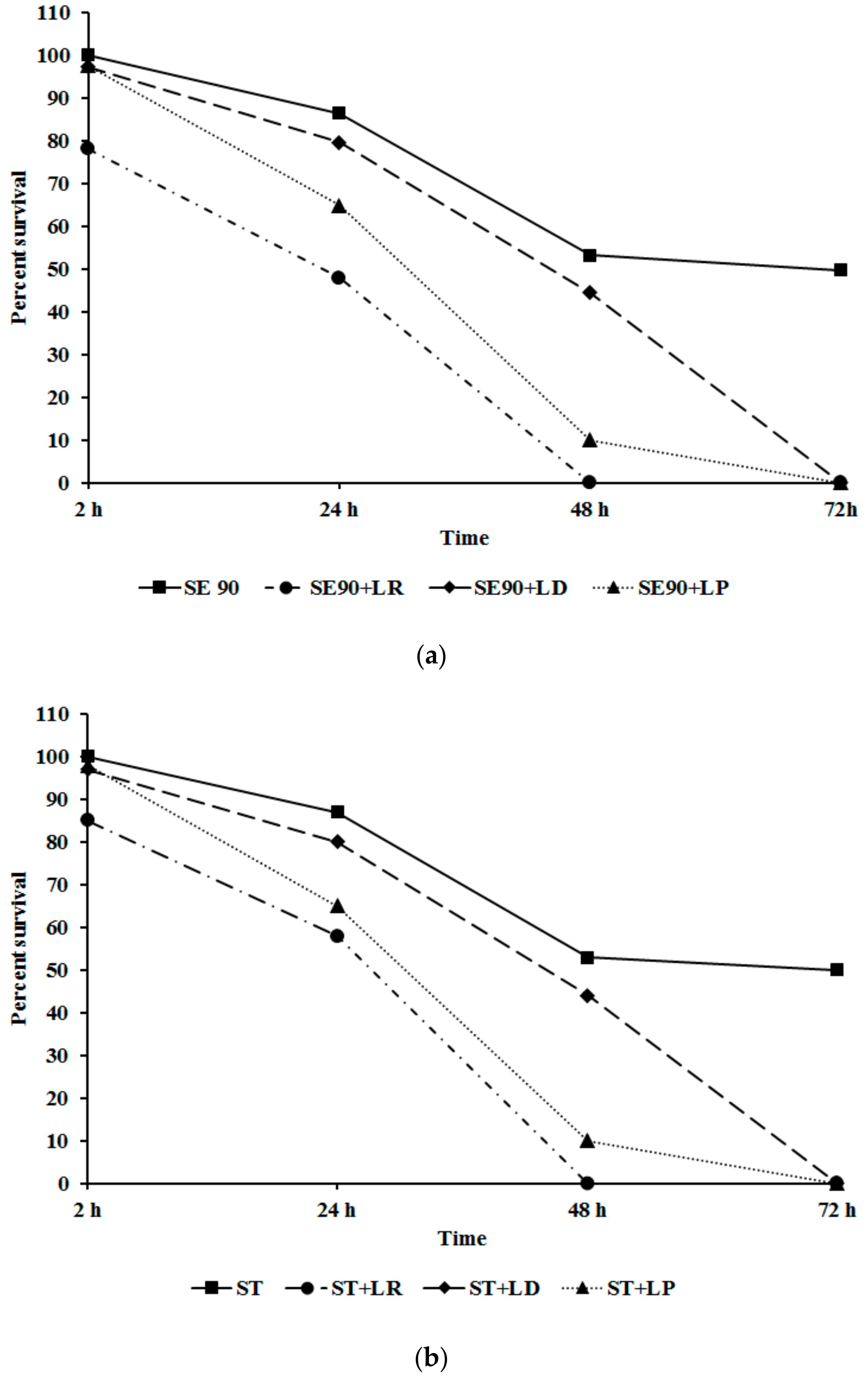
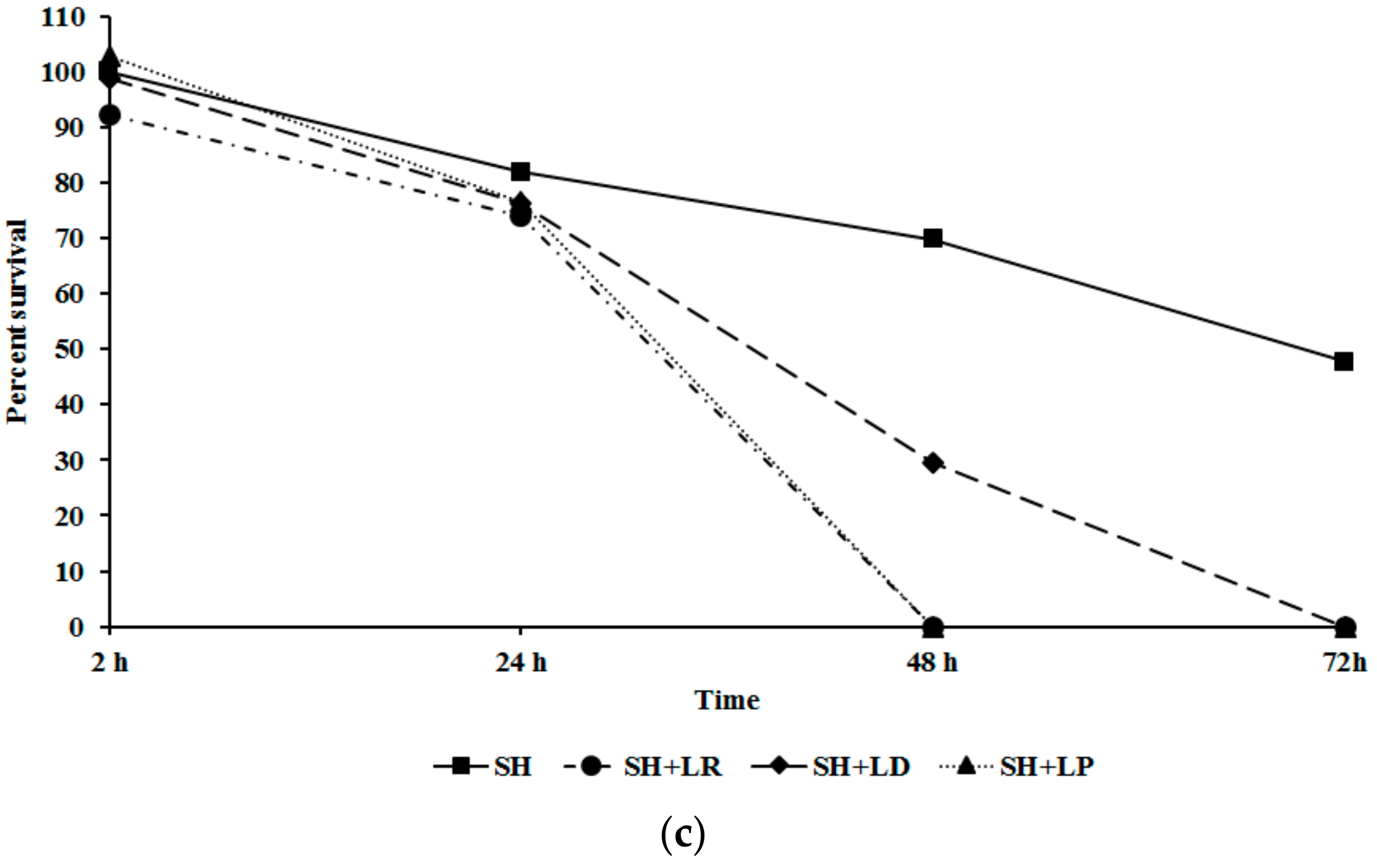
2.4. Inhibition of Salmonella Invasion and Survival in Chicken Macrophage
3. Materials and Methods
3.1. Bacterial Isolates and Growth Conditions
3.2. Estimation of Sub-Inhibitory Concentration of LAB Cell-Free Supernatant
3.3. Motility Assay
3.4. Isolation of Chicken Primary CEC
3.5. Inhibition of Salmonella Adhesion to CEC
3.6. Inhibition of Salmonella Invasion in CEC
3.7. Inhibition of Salmonella Invasion and Survival in HTC
3.8. Effect of LAB on Salmonella Virulence Gene Expression
3.9. Transmission Electron Microscopy (TEM) Analysis
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. International Collaboration on Enteric Disease ‘Burden of Illness’ Studies: The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [PubMed]
- Antunes, P.; Mourao, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [PubMed]
- Jackson, B.R.; Griffin, P.M.; Cole, D.; Walsh, K.A.; Chai, S.J. Outbreak-associated Salmonella enterica serotypes and food Commodities, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 1239–1244. [Google Scholar] [PubMed]
- Andino, A.; Hanning, I. Salmonella Enterica: Survival, colonization, and virulence differences among serovars. Sci. World J. 2015, 2015, 520179. [Google Scholar] [CrossRef] [PubMed]
- Rajan, K.; Shi, Z.; Ricke, S.C. Current aspects of Salmonella contamination in the US poultry production chain and the potential application of risk strategies in understanding emerging hazards. Crit. Rev. Microbiol. 2017, 43, 370–392. [Google Scholar] [PubMed]
- Chousalkar, K.; Gast, R.; Martelli, F.; Pande, V. Review of egg-related salmonellosis and reduction strategies in United States, Australia, United Kingdom and New Zealand. Crit. Rev. Microbiol. 2017, 14, 1–14. [Google Scholar]
- Alali, W.Q.; Hofacre, C.L. Preharvest food safety in broiler chicken production. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Chittick, P.; Sulka, A.; Tauxe, R.V.; Fry, A.M. A summary of national reports of foodborne outbreaks of Salmonella Heidelberg infections in the United States: Clues for disease prevention. J. Food Prot. 2006, 69, 1150–1153. [Google Scholar] [CrossRef] [PubMed]
- Gast, R.K.; Guraya, R.; Jones, D.R.; Anderson, K.E.; Karcher, D.M. Colonization of internal organs by Salmonella Enteritidis in experimentally infected laying hens housed in enriched colony cages at different stocking densities. Poult. Sci. 2016, 95, 1363–1369. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Tian, W.; Qian, N.; Cheng, A.; Deng, S. Quantitative studies of the distribution pattern for Salmonella Enteritidis in the internal organs of chicken after oral challenge by a real-time PCR. Vet. Res. Commun. 2010, 34, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, I.; Upadhyay, A.; Kollanoor-Johny, A.; Darre, M.J.; Venkitanarayanan, K. Effect of plant derived antimicrobials on Salmonella Enteritidis adhesion to and invasion of primary chicken oviduct epithelial cells in vitro and virulence gene expression. Int. J. Mol. Sci. 2013, 14, 10608–10625. [Google Scholar] [CrossRef] [PubMed]
- Harvey, P.C.; Watson, M.; Hulme, S.; Jones, M.A.; Lovell, M.; Berchieri, A., Jr.; Young, J.; Bumstead, N.; Barrow, P. Salmonella enterica serovar Typhimurium colonizing the lumen of the chicken intestine grows slowly and upregulates a unique set of virulence and metabolism genes. Infect. Immun. 2011, 79, 4105–4121. [Google Scholar] [CrossRef] [PubMed]
- Berry, E.D.; Wells, J.E. Reducing foodborne pathogen persistence and transmission in animal production environments: Challenges and opportunities. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Umaraw, P.; Prajapati, A.; Verma, A.K.; Pathak, V.; Singh, V.P. Control of Campylobacter in poultry industry from farm to poultry processing unit: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Lin, J. Novel approaches for Campylobacter control in poultry. Foodborne Pathog. Dis. 2009, 6, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Vandeplas, S.; Dubois Dauphin, R.; Beckers, Y.; Thonart, P.; Thewis, A. Salmonella in chicken: Current and developing strategies to reduce contamination at farm level. J. Food Prot. 2010, 73, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Fooks, L.J.; Gibson, G.R. In vitro investigations of the effect of probiotics and prebiotics on selected human intestinal pathogens. FEMS Microbiol. Ecol. 2002, 39, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Fuller, R. Probiotic in man and animals: A review. J. Appl. Bacteriol. 1989, 90, 352. [Google Scholar]
- Doré, J.; Corthier, G. Le microbiote intestinal humain. Gastroentérologie Clinique et Biologique 2010, 34, 7–16. [Google Scholar] [CrossRef]
- Gusils, C.; Bujazha, M.; Gonzalez, S. Preliminary studies to design a probiotic for use in swine feed. Interciencia 2002, 27, 409–413. [Google Scholar]
- Awad, W.A.; Ghareeb, K.; Abdel-Raheem, S.; Bohm, J. Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poult. Sci. 2009, 88, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Stavric, S.; Kornegay, E.T. Microbial probiotics for pigs and poultry. In Biotechnology in Animal Feeds and Feeding; Wallace, R.J., Chesson, A., Eds.; Wiley-VCH: New York, NY, USA, 1995; pp. 205–231. [Google Scholar]
- Rolfe, R.D. The role of probiotic cultures in the control of gastrointestinal health. J. Nutr. 2000, 130, 396S–402S. [Google Scholar] [PubMed]
- Upadhyaya, I.; Upadhyay, A.; Yin, H.B.; Nair, M.S.; Bhattaram, V.K.; Karumathil, D.; Kollanoor-Johny, A.; Khan, M.I.; Darre, M.J.; Curtis, P.A.; et al. Reducing Colonization and Eggborne Transmission of Salmonella Enteritidis in Layer Chickens by In-Feed Supplementation of Caprylic Acid. Foodborne Pathog. Dis. 2015, 12, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.H.; Benson, C.E.; Krotec, K.; Eckroade, R.J. Salmonella Enteritidis colonization of the reproductive tract and forming and freshly laid eggs of chickens. Infect. Immun. 1995, 63, 2443–2449. [Google Scholar] [PubMed]
- Gantois, I.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F.; Gast, R.; Humphrey, T.J.; Van Immerseel, F. Mechanisms of egg contamination by Salmonella Enteritidis. FEMS Microbiol. Rev. 2009, 33, 718–738. [Google Scholar] [CrossRef] [PubMed]
- Muramoto, K.; Macnab, R.M. Deletion analysis of MotA and MotB, components of the force-generating unit in the flagellar motor of Salmonella. Mol. Microbiol. 1998, 29, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, Y.V.; Che, Y.S.; Minamino, T.; Namba, K. Proton-conductivity assay of plugged and unplugged MotA/B proton channel by cytoplasmic pHluorin expressed in Salmonella. FEBS Lett. 2010, 584, 1268–1272. [Google Scholar] [CrossRef] [PubMed]
- Misselwitz, B.; Barrett, N.; Kreibich, S.; Vonaesch, P.; Andritschke, D.; Rout, S.; Weidner, K.; Sormaz, M.; Songhet, P.; Horvath, P.; et al. Near surface swimming of Salmonella Typhimurium explains target-site selection and cooperative invasion. PLoS Pathog. 2012, 8, e1002810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorkowski, M.; Felipe-Lopez, A.; Danzer, C.A.; Hansmeier, N.; Hensel, M. Salmonella enterica invasion of polarized epithelial cells is a highly cooperative effort. Infect. Immun. 2014, 82, 2657–2667. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, Y.V.; Nakamura, S.; Kami-ike, N.; Namba, K.; Minamino, T. Charged residues in the cytoplasmic loop of MotA are required for stator assembly into the bacterial flagellar motor. Mol. Microbiol. 2010, 78, 1117–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saijo-Hamano, Y.; Matsunami, H.; Namba, K.; Imada, K. Expression, purification, crystallization and preliminary X-ray diffraction analysis of a core fragment of FlgG, a bacterial flagellar rod protein. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Amy, M.T.; Virlogeux-Payant, I.; Bottreau, E.; Mompart, F.; Pardon, P.; Velge, P. Precise excision and secondary transposition of TnphoA in non-motile mutants of a Salmonella enterica serovar Enteritidis clinical isolate. FEMS Microbiol. Lett. 2005, 245, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; De Buck, J.; De Smet, I.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R. Interactions of butyric acid- and acetic acid-treated Salmonella with chicken primary cecal epithelial cells in vitro. Avian Dis. 2004, 48, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Lim, C.Y.; Teng, W.L.; Ouwehand, A.C.; Tuomola, E.M.; Salminen, S. Quantitative approach in the study of adhesion of lactic acid bacteria to intestinal cells and their competition with enterobacteria. Appl. Environ. Microbiol. 2000, 66, 3692–3697. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.B.; Curtiss, R. Effect of Inv Mutations on Salmonella Virulence and Colonization in 1-Day-Old White Leghorn Chicks. Avian Dis. 1997, 41, 45. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Z.; Pace, L.; Lillehoj, H.; Zhang, S. Functions exerted by the virulence-associated type-three secretion systems during Salmonella enterica serovar Enteritidis invasion into and survival within chicken oviduct epithelial cells and macrophages. Avian Pathol. 2009, 38, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Akbar, S.; Schechter, L.M.; Lostroh, C.P.; Lee, C.A. AraC/XylS family members, HilD and HilC, directly activate virulence gene expression independently of HilA in Salmonella Typhimurium. Mol. Microbiol. 2003, 47, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.W.; Jones, M.A.; Watson, P.R.; Hedges, S.; Wallis, T.S.; Galyov, E.E. Identification of a pathogenicity island required for Salmonella enteropathogenicity. Mol. Microbiol. 1998, 29, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Santos, R.L.; Tsolis, R.M.; Stender, S.; Hardt, W.-D.; Baumler, A.J.; Adams, L.G. The Salmonella enterica serotype Typhimurium effector proteins SipA, SopA, SopB, SopD, and SopE2 act in concert to induce diarrhea in calves. Infect. Immun. 2002, 70, 3843–3855. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.R.; Galyov, E.E.; Paulin, S.M.; Jones, P.W.; Wallis, T.S. Mutation of invH, but not stn, reduces Salmonella-induced enteritis in cattle. Infect. Immun. 1998, 66, 1432–1438. [Google Scholar] [PubMed]
- Eichelberg, K.; Galan, J.E. Differential regulation of Salmonella Typhimurium type III secreted proteins by pathogenicity island 1 (SPI-1)-encoded transcriptional activators InvF and HilA. Infect. Immun. 1999, 67, 4099–4105. [Google Scholar] [PubMed]
- Lucas, R.L.; Lee, C.A. Roles of hilC and hilD in regulation of hilA expression in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2001, 183, 2733–2745. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.H.; Lee, M.; Park, J.; Lee, J.; Eo, S.; Kwon, J.; Chae, J. Identification of Salmonella Gallinarum virulence genes in a chicken infection model using PCR-based signature-tagged mutagenesis. Microbiology 2005, 151, 3957–3968. [Google Scholar] [CrossRef] [PubMed]
- Pati, N.B.; Vishwakarma, V.; Jaiswal, S.; Periaswamy, B.; Hardt, W.D.; Suar, M. Deletion of invH gene in Salmonella enterica serovar Typhimurium limits the secretion of Sip effector proteins. Microbes Infect. 2013, 15, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Fields, P.I.; Swanson, R.V.; Haidaris, C.G.; Heffron, F. Mutants of Salmonella Typhimurium that cannot survive within the macrophage are avirulent. Proc. Natl. Acad. Sci. USA 1986, 83, 5189–5193. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, D.M.; Valdivia, R.H.; Monack, D.M.; Falkow, S. Macrophage-dependent induction of the Salmonella pathogenicity island 2 type III secretion system and its role in intracellular survival. Mol. Microbiol. 1998, 30, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Knodler, L.A.; Vallance, B.A.; Hensel, M.; Jäckel, D.; Finlay, B.B.; Steele-Mortimer, O. Salmonella type III effectors PipB and PipB2 are targeted to detergent-resistant microdomains on internal host cell membranes. Mol. Microbiol. 2003, 49, 685–704. [Google Scholar] [CrossRef] [PubMed]
- Charles, S. The Proteomic Response of the Probiotic Pediococcus acidilactici R1001 after Exposure to the Cell-Free Supernatant of the Pathogen Listeria monocytogenes ATCC 7644; MSc: UGuelph, ON, Canada, 2016. [Google Scholar]
- Kollanoor-Johny, A.; Mattson, T.; Baskaran, S.A.; Amalaradjou, M.A.; Babapoor, S.; March, B.; Valipe, S.; Darre, M.; Hoagland, T.; Schreiber, D.; et al. Reduction of Salmonella enterica serovar Enteritidis colonization in 20-day-old broiler chickens by the plant-derived compounds trans-cinnamaldehyde and eugenol. Appl. Environ. Microbiol. 2012, 78, 2981–2987. [Google Scholar] [CrossRef] [PubMed]
- Molina-Quiroz, R.C.; Silva, C.A.; Molina, C.F.; Leiva, L.E.; Reyes-Cerpa, S.; Contreras, I.; Santiviago, C.A. Exposure to sub-inhibitory concentrations of cefotaxime enhances the systemic colonization of Salmonella Typhimurium in BALB/c mice. Open Biol. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Gilbert, E.S. Colorimetric method for identifying plant essential oil components that affect biofilm formation and structure. Appl. Environ. Microbiol. 2004, 70, 6951–6956. [Google Scholar] [CrossRef] [PubMed]
- Amalaradjou, M.A.; Kim, K.S.; Venkitanarayanan, K. Sub-inhibitory concentrations of trans-cinnamaldehyde attenuate virulence in Cronobacter sakazakii in vitro. Int. J. Mol. Sci. 2014, 15, 8639–8655. [Google Scholar] [CrossRef] [PubMed]
- Dodson, S.V.; Maurer, J.J.; Holt, P.S.; Lee, M.D. Temporal changes in the population genetics of Salmonella Pullorum. Avian Dis. 1999, 43, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Henderson, S.C.; Bounous, D.I.; Lee, M.D. Early events in the pathogenesis of avian salmonellosis. Infect. Immun. 1999, 67, 3580–3586. [Google Scholar] [PubMed]
- Hudson, C.R.; Quist, C.; Lee, M.D.; Keyes, K.; Dodson, S.V.; Morales, C.; Sanchez, S.; White, D.G.; Maurer, J.J. Genetic relatedness of Salmonella isolates from nondomestic birds in Southeastern United States. J. Clin. Microbiol. 2000, 38, 1860–1865. [Google Scholar] [PubMed]
- Nair, D.V.T.; Kollanoor-Johny, A. Effect of Probionibacterium freudenreichii on Salmonella multiplication, motility, and association with avian epithelial cells. Poult. Sci. 2017, 96, 1513. [Google Scholar] [CrossRef] [PubMed]
- Koo, O.K.; Amalaradjou, M.A.; Bhunia, A.K. Recombinant probiotic expressing Listeria adhesion protein attenuates Listeria monocytogenes virulence in vitro. PLoS ONE 2012, 7, e29277. [Google Scholar] [CrossRef] [PubMed]
- Kollanoor-Johny, A.; Mattson, T.; Baskaran, S.A.; Amalaradjou, M.A.; Hoagland, T.A.; Darre, M.J.; Khan, M.I.; Schreiber, D.T.; Donoghue, A.M.; Donoghue, D.J.; et al. Caprylic acid reduces Salmonella Enteritidis populations in various segments of digestive tract and internal organs of 3- and 6-week-old broiler chickens, therapeutically. Poult. Sci. 2012, 91, 1686–1694. [Google Scholar] [CrossRef] [PubMed]
- Kannan, L.; Rath, N.C.; Liyanage, R.; Lay, J.O., Jr. Identification and characterization of thymosin beta-4 in chicken macrophages using whole cell MALDI-TOF. Ann. N. Y. Acad. Sci. 2007, 1112, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Bayoumi, M.A.; Griffiths, M.W. In vitro inhibition of expression of virulence genes responsible for colonization and systemic spread of enteric pathogens using Bifidobacterium bifidum secreted molecules. Int. J. Food Microbiol. 2012, 156, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Bookout, A.L.; Mangelsdorf, D.J. Quantitative real-time PCR protocol for analysis of nuclear receptor signaling pathways. Nucl. Recept. Signal. 2003, 1, e012. [Google Scholar] [CrossRef] [PubMed]
- Berton, V.; Montesi, F.; Losasso, C.; Facco, D.R.; Toffan, A.; Terregino, C. Study of the interaction between silver nanoparticles and Salmonella as revealed by transmission electron microscopy. J. Probiotics Health 2015, 3, 123. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | invH | hilA | hilD | sipA | sipB | sopB | spvB |
|---|---|---|---|---|---|---|---|
| SE Ctrl | 1 a | 1 a | 1 a | 1 a | 1 a | 1 a | 1 a |
| SE + LR | –1.85 ± 0.08 b | –4.58 ± 1.13 b–d | –2.90 ± 0.15 b,c | –5.01 ± 0.76 b–d | –18.69 ± 0.46 g | –6.67 ± 1.39 b–d | –8.17± 1.55 b–d |
| SE + LD | –2.66± 0.14 b,c | –3.54 ± 0.71 b,c | –2.20 ± 0.10 b | –1.87 ± 0.15 b | –1.91 ± 0.19 b | –2.93 ± 0.44 b,c | –17.27 ± 3.26 f,g |
| SE + LP | –4.07± 0.84 a,b | –4.07 ± 0.38 b–d | –2.81 ± 0.28 b,c | –11.19± 0.36 d,e | –27.19 ± 0.66 i | –15.70± 0.54 e–g | –25.46 ± 0.91 h,i |
| ST Ctrl | 1 a | 1 a | 1 a | 1 a | 1 a | 1 a | 1 a |
| ST + LR | –11.31± 0.22 d,e | –2.31 ± 1.04 b,c | –2.74 ± 0.9 b,c | –2.89 ± 1.12 b | –19.35 ± 0.27 g | –3.55 ± 0.51 b,c | –7.16 ± 0.88 b–d |
| ST + LD | –3.98 ± 1.2 a,b | –3.98 ± 0.93 b–d | –2.09 ± 0.66 b | –1.37 ± 0.28 b | –1.78 ± 0.28 b | –1.89 ± 0.28 b | –10.40 ± 1.17 d,e |
| ST + LP | –3.05± 0.77 b,c | –1.45 ± 0.56 b | –2.72 ± 0.55 b,c | –12.13± 1.65 d–f | –28.92 ± 1.53 i | –6.00 ± 1.93 b–d | –25.46 ± 4.8 h,i |
| SH Ctrl | 1 a | 1 a | 1 a | 1 a | 1 a | 1 a | 1 a |
| SH + LR | –3.12 ± 0.64 b,c | –2.00 ± 0.2 a | –2.54 ± 0.6 b,c | –3.29 ± 0.16 b,c | –17.21 ± 3.45 f,g | –3.22 ± 0.33 b,c | –19.97 ± 0.22 g,h |
| SH+ LD | –4.42 ± 1.09 a,b | –5.76 ± 0.55 b–d | –2.04 ± 0.63 b | –1.67 ± 0.17 b | –3.29 ± 0.75 b,c | –2.22 ± 0.23 b | –19.61 ± 0.53 g,h |
| SH + LP | –5.45 ± 2.53 a,b | –12.11± 0.55 d–f | –2.68 ± 0.22 b,c | –10.53 ± 0.99 d,e | –27.96 ± 1.99 i | –4.69 ± 0.67 b–d | –30.86 ± 1.41 i |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muyyarikkandy, M.S.; Amalaradjou, M.A. Lactobacillus bulgaricus, Lactobacillus rhamnosus and Lactobacillus paracasei Attenuate Salmonella Enteritidis, Salmonella Heidelberg and Salmonella Typhimurium Colonization and Virulence Gene Expression In Vitro. Int. J. Mol. Sci. 2017, 18, 2381. https://doi.org/10.3390/ijms18112381
Muyyarikkandy MS, Amalaradjou MA. Lactobacillus bulgaricus, Lactobacillus rhamnosus and Lactobacillus paracasei Attenuate Salmonella Enteritidis, Salmonella Heidelberg and Salmonella Typhimurium Colonization and Virulence Gene Expression In Vitro. International Journal of Molecular Sciences. 2017; 18(11):2381. https://doi.org/10.3390/ijms18112381
Chicago/Turabian StyleMuyyarikkandy, Muhammed Shafeekh, and Mary Anne Amalaradjou. 2017. "Lactobacillus bulgaricus, Lactobacillus rhamnosus and Lactobacillus paracasei Attenuate Salmonella Enteritidis, Salmonella Heidelberg and Salmonella Typhimurium Colonization and Virulence Gene Expression In Vitro" International Journal of Molecular Sciences 18, no. 11: 2381. https://doi.org/10.3390/ijms18112381
APA StyleMuyyarikkandy, M. S., & Amalaradjou, M. A. (2017). Lactobacillus bulgaricus, Lactobacillus rhamnosus and Lactobacillus paracasei Attenuate Salmonella Enteritidis, Salmonella Heidelberg and Salmonella Typhimurium Colonization and Virulence Gene Expression In Vitro. International Journal of Molecular Sciences, 18(11), 2381. https://doi.org/10.3390/ijms18112381