1. Introduction
Cold and drought are the most common and primary environments that adversely affect plant growth and crop production worldwide. Because plants are sessile and have to endure one or more environmental challenges, plants have developed multiple complex signaling networks involved in stress perception, signal transduction, and the expression of stress-related genes to cope with adverse environmental conditions during their long-term adaptation [
1]. The calcium ion (Ca
2+) is recognized as a secondary messenger and appears to play crucial roles in signal transduction under both cold and drought stresses, via the changes in intracellular Ca
2+ concentration [
2]. Several kinds of Ca
2+ sensors, including calmodulin (CaM), CaM-like protein (CML), calcineurin B-like protein (CBL), and Ca
2+-dependent protein kinase (CDPK), are induced in response to cold and drought [
2]. Moreover,
Arabidopsis transgenic plants with over-expression of
GmCaM4 or
VaCDPK20 exhibit enhanced stress tolerance [
3,
4]. Mitogen-activated protein kinase (MAPK), another secondary messenger, is an important mediator in signal transmission, connecting the perception of external stimuli to cellular responses. Previous studies have demonstrated that MAPKs are involved in cold, drought, or both cold and drought stresses [
5,
6]. Reactive oxygen species (ROS), such as superoxide, hydrogen peroxide, and hydroxyl radicals, are also secondary messengers and accumulate under cold and drought stresses. They are signals contributing to stress injury in plants, as transgenic plants over-expressing ROS scavengers or mutants with higher ROS-scavenging ability show increased tolerance to environmental stresses [
7].
In addition to the secondary messengers, hormones have been demonstrated to be key players in the response to cold and drought. Of these, abscisic acid (ABA) plays a critical role in response to various stress signals [
8]. Under osmotic conditions imposed by drought, the most rapid response of plants is closure of the stomata, mediated by ABA, to maintain their water balance. Accordingly, expression of several ABA biosynthesis genes, such as
NCED3 and
LOS6/
ABA1, are up-regulated by drought stress [
9]. In addition to the ABA-dependent pathway, molecular analyses have revealed that ABA-independent pathways are also involved in transcriptional regulatory networks under drought stress [
8,
10]. However, under cold stress, the expression of genes is influenced largely through an ABA-independent pathway, because expression of some key genes (e.g.,
ZEP and
NCED) involved in ABA biosynthesis is not obviously up-regulated by cold and there is little or no increase in ABA content in plants subjected to cold treatment [
7,
11]. It is noteworthy that the expression of
Arabidopsis gene
RD29A was subject to both ABA-dependent and -independent regulation under both drought and cold conditions [
1]. In addition to ABA, hormones such as ethylene, salicylic acid (SA), and brassinosteroid (BR) are also involved in abiotic stress responses [
12,
13]. Moreover, these hormones interact with one another in regulating stress signaling and tolerance in plants [
13].
Transcription factors (TFs) are also key regulators involved in plant abiotic stress signaling. Many cold-regulated genes (
COR) have been identified as responding to cold treatment. The most characterized transcriptional cascade, with a large role in cold stress pathways, is called “ICE-CBF-COR” [
1]. Of these, C-repeat binding factors (CBFs) are members of the AP2/EREBP family and are involved in regulation of ABA-independent expression of
COR genes under cold stress. Constitutive- or over-expression of CBFs enhances cold stress tolerance in several plants [
14,
15]. Inducer of CBF expression (
ICE), a member of the bHLH family, can directly interact with CBFs and regulate their expression upon cold treatment [
16]. Moreover, the over-expression of
ICEs in transgenic plants results in improved freezing tolerance [
17]. ABA is a major player in response to drought stress, thus the TFs involved in ABA signaling pathways are related to drought stress. For example, ABRE-binding factor (ABF), of the bZIP family, can bind to the ABRE motif and activate ABA-dependent gene expression during drought stress [
18]. Other important TFs, such as the MYC and myeloblastosis (MYB) proteins, are also known to function as activators in ABA-dependent regulatory pathways. Transgenic plants over-expressing both
AtMYC2 and
AtMYB2 showed an ABA-hypersensitive phenotype and improved osmotic stress tolerance [
8,
19]. In addition to the cold- or drought-specific induced TFs, some others (e.g.,
WRKY45,
ZAT12, and
NAC6) are involved in both cold and drought stresses.
To further increase the understanding of signaling pathways for the improvement of crop stress tolerance, it is essential to know how plants respond to stresses and which genes and pathways are involved in the stress tolerance. Over recent decades, significant progress has been made regarding our knowledge of plant response to abiotic stresses, and thousands of genes and tens of signaling pathways have been identified concerning cold and drought [
8,
20]. Moreover, many of these genes were found to be regulated commonly or specifically in response to drought and cold stresses in several plants, including
Arabidopsis [
21], rice [
22], and cotton [
23] at the transcriptional level. However, in tropical plants (e.g., cassava), similar studies are very limited, and the molecular mechanisms underlying tolerance to both drought and cold stresses remain largely unknown.
Cassava (
Manihot esculenta Crantz) is an important cash crop and provides staple food for over 750 million people in tropical and sub-tropical areas [
24]. It is a major crop for starch, bio-fuel production, and animal feed, owing to its starch-enriched root [
25]. Cassava is generally thought to be drought-tolerant, however, similarly to other crops, this drought tolerance is usually at the cost of restrained growth and decreased economic yield [
25]. As a tropical root crop, cassava is native to a warm habitat and is very sensitive to cold, which greatly damages the plants and reduces yield [
26]. Thus, it is very important to increase our understanding of the molecular mechanisms of cold and drought stresses in cassava. With rapid development of next generation sequencing techniques and the release of the cassava draft genome [
27,
28], much progress has been made in large-scale identification of genes involved in cold [
26,
29] and drought [
24,
30,
31] responses, respectively, in cassava at the transcriptional or proteomic levels. However, very few studies have focused on comparisons of transcriptome profiles in response to cold and drought stresses in cassava, and the related signaling pathways have not been adequately explored. In this study, the same cassava cultivar leaves (including folded and full-expanded leaves) and roots were respectively collected at 0, 6, 24, and 48 h after cold treatment and 0, 4, 6, and 10 days after drought treatment, to investigate the genome-wide transcriptome changes through RNA-seq technology. The results will provide new insights into the signaling regulation networks responding to cold and drought in cassava, and will also provide solid bases for cassava improvement in regard to these two abiotic stresses.
2. Results
2.1. Stress Responses and Transcriptional Profiling of Cassava
Compared with controls, there were respective distinct symptoms in the cold- and drought-treated plants as the treatment time progressed (
Figure S1). For the cold treatment, the plants were badly wilted at 6 h, their top leaves were curved at 24 h, and symptoms were more serious at 48 h. The drought-treated plants were badly wilted at 4 days, their lower leaves began to fall off at 6 days, and only a few upper leaves remained at 10 days (
Figure S1).
To investigate the spatial and temporal gene expression in response to abiotic stresses in cassava at the transcriptional level, different tissues, including folded leaves (FL, the first three folded leaves), fully expanded leaves (FEL, three most newly fully expanded leaves), and fibrous roots (RT), were respectively collected at 0, 6, 24, and 48 h after cold treatment and 0, 4, 6, and 10 days after drought treatment. Overall, 88.2 million raw reads of 35 bp in length were obtained by single end sequencing with the Illumina GAII platform. After sequence pre-processing, 83.4 million adapter-trimmed clean reads were kept, and 95.8% of them were mapped to the cassava reference genome (Available online:
http://www.phytozome.net) with a maximum of two mismatches.
Overall, 18,271 genes, approximately two-thirds of the annotated genes in the reference genome, were expressed across drought-treated samples. Comparatively, a similar number of genes (17,352) were expressed across cold-treated samples. Interestingly, in either cold or drought treatment, the numbers of expressed genes were quite similar in the samples of all examined time points within the same tissue, as well as in different tissues when considering all the time points together (i.e., a gene was counted if it was expressed in at least one of four time points within the same tissue) (
Figure S2). In addition, in both cold and drought treatments, most of these genes were commonly expressed while only a few were exclusively expressed in FL, FEL, and RT, respectively (
Figure S2). The results indicated that there were similar amounts of expressed genes in different tissues (e.g., FL, FEL, and RT) of cassava to maintain a kind of housekeeping effect during cassava growth and development, regardless of experiencing abiotic stress or not. Once subjected to abiotic stress, the expression of these genes in cassava would be up- or down-regulated to cope with the adverse environments.
In addition, seven genes of interest were selected and their expression patterns were validated with high correlation coefficient (r = 0.94) between RNA-seq and qRT-PCR methods (
Table S1).
2.2. Transcriptome Changes in Response to Drought and Cold
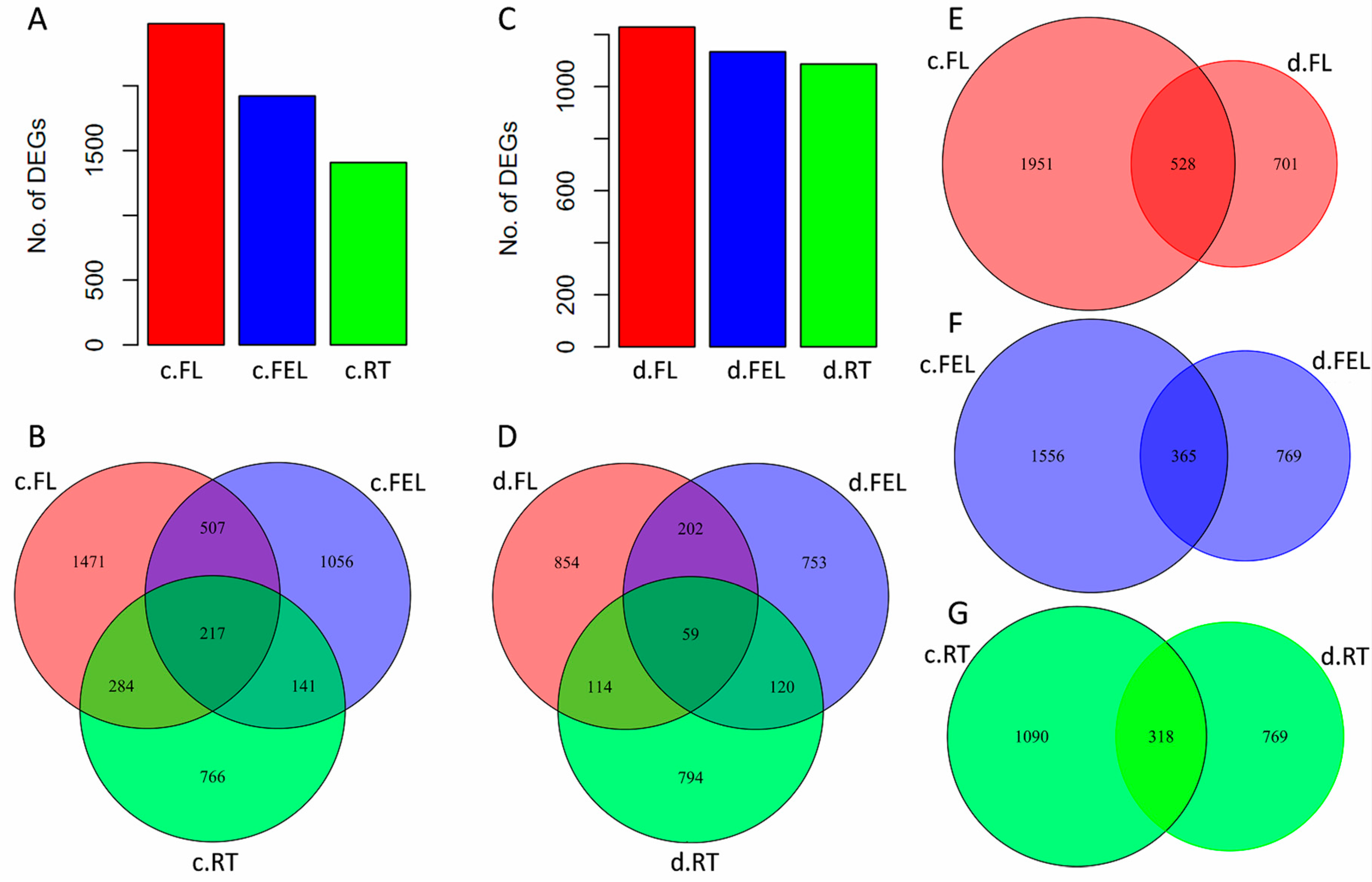
To reveal the transcriptome changes affected by abiotic stresses, differentially expressed (DE) genes were identified under cold and drought, respectively. Under cold treatment (signified by prefix “c.”), there were 2479, 1921, and 1408 DE genes in c.FL, c.FEL, and c.RT, respectively, with an increased gradient change of DE gene number from bottom to top of a plant (
Figure 1A). Unlike expressed genes, most of these DE genes were exclusively identified in c.FL (1471), c.FEL (1056), and c.RT (766), but only 217 were commonly identified in all of these tissues (
Figure 1B). Under drought treatment (signified by prefix “d.”), a similar gradient change of DE gene number was observed: d.FL (1229) > d.FEL (1134) > d.RT (1087), although the numbers were slightly lower compared with those of the cold treatment (
Figure 1C). Similarly, most of these DE genes were also exclusively identified in d.FL (854), d.FEL (753), and d.RT (794), and only a few (59) were commonly identified (
Figure 1D). Thus, in response to cold and drought, expression of DE genes was more influenced in cassava leaves than in roots, and the majority exhibited tissue-specific expression patterns while only a small proportion were commonly influenced across tissue types.
The DE genes were compared between cold and drought treatments within the same tissue to explore their roles and expression responses to different abiotic stresses. In FL, 1951 (61.4%) DE genes were uniquely identified under cold treatment (
Figure 1E). Functional category enrichment showed that these genes were significantly enriched in calcium signaling and ABC transporters (
Figure S3). As compared, 701 (22.0%) DE genes were uniquely identified under drought and 528 (16.6%) were identified under both cold and drought (
Figure 1E). These two groups of genes were commonly enriched in several categories, including cell wall, photosynthesis, and abiotic stress (
Figure S3). In FEL, 1556 (57.8%) and 769 (28.6%) DE genes were uniquely identified in cold and drought, and they were enriched in photosynthesis and tetrapyrrole synthesis, and redox and abiotic stress, respectively (
Figure S3). In total, 365 (13.6%) DE genes were commonly found in response to cold and drought (
Figure 1F), however, no significantly enriched categories were observed. In RT, 1090 (50.0%) DE genes were uniquely identified under cold treatment (
Figure 1G), but there were no significantly enriched categories. A total of 769 (35.3%) DE genes were uniquely identified under drought, and 318 (14.6%) were commonly identified under both cold and drought (
Figure 1G); these genes were significantly enriched in secondary metabolism and abiotic stress, respectively (
Figure S3). These results also revealed that the expression of genes was more influenced by cold (~55%) than drought (~30%), and only a few of genes (~15%) were commonly influenced by both cold and drought in FL, FEL, or RT (
Figure 1E–G).
To further characterize the functions of DE genes that were uniquely or commonly identified in cold and drought stress, fold-change-based clustering analysis and functional category enrichment were performed for each group, respectively.
2.3. DE Genes Specifically Responding to Cold
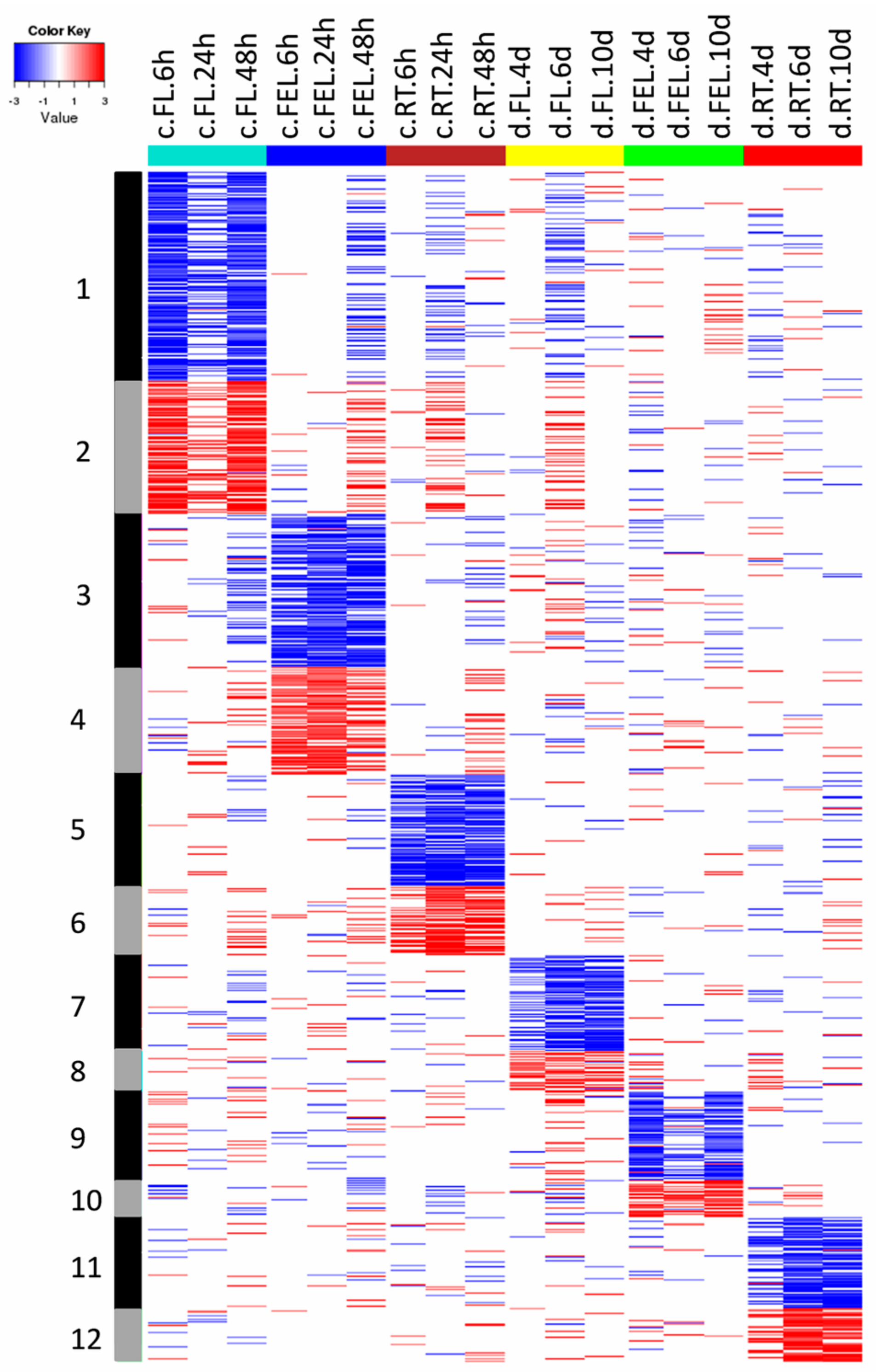
In total, 38 groups (G1–G38) comprising 5030 DE genes that were differentially expressed in at least one tissue, were identified (
Table S2). As shown in
Figure 2, the expression of genes from groups G1–G6 was significantly changed in only one tissue in response to cold. For example, the expression of 604 genes in G1 and 383 genes in G2 was significantly decreased and increased in FL, respectively. Category enrichment showed that these genes were mainly involved in cell-related metabolism, and protein degradation and ABC transporters, respectively (
Figure 3). There were 439 genes in G3 and 311 in G4 of which the expression was significantly decreased and increased in FEL, respectively. The enriched categories in G3 included Calvin cycle and light reaction of photosynthesis, and tetrapyrrole synthesis, however, no enriched categories were found in G4 (
Figure 3). Likewise, there were 319 genes in G5 and 201 in G6 whose expression was significantly decreased and increased in RT, respectively (
Figure 2). No enriched categories were found in G5, however, the genes of G6 were mainly involved in trehalose metabolism (
Figure 3).
The expression of genes from groups G13–G17 was significantly changed in more than one tissue exclusively in response to cold (
Figure 4). For example, the expression of genes in G13 was significantly decreased in both FL and FEL, and these genes mainly participated in hormone metabolism (e.g., BR), light reaction of photosynthesis, and tetrapyrrole synthesis (
Figure 3). On the contrary, the expression of G15 genes increased in both FL and FEL. Similarly, genes decreased in G16 but increased in G17 in both FL and RT, genes decreased in G23 but increased in G24 in both FEL and RT, and genes decreased but increased in all the three tissues, respectively (
Figure 4). However, no significantly enriched categories were found in most of these groups (
Figure 3).
Together, the results indicated that DE genes, which exclusively responded to cold, were mainly involved in cell related metabolism, protein degradation, Calvin cycle and light reaction of photosynthesis, tetrapyrrole synthesis, and BR metabolism.
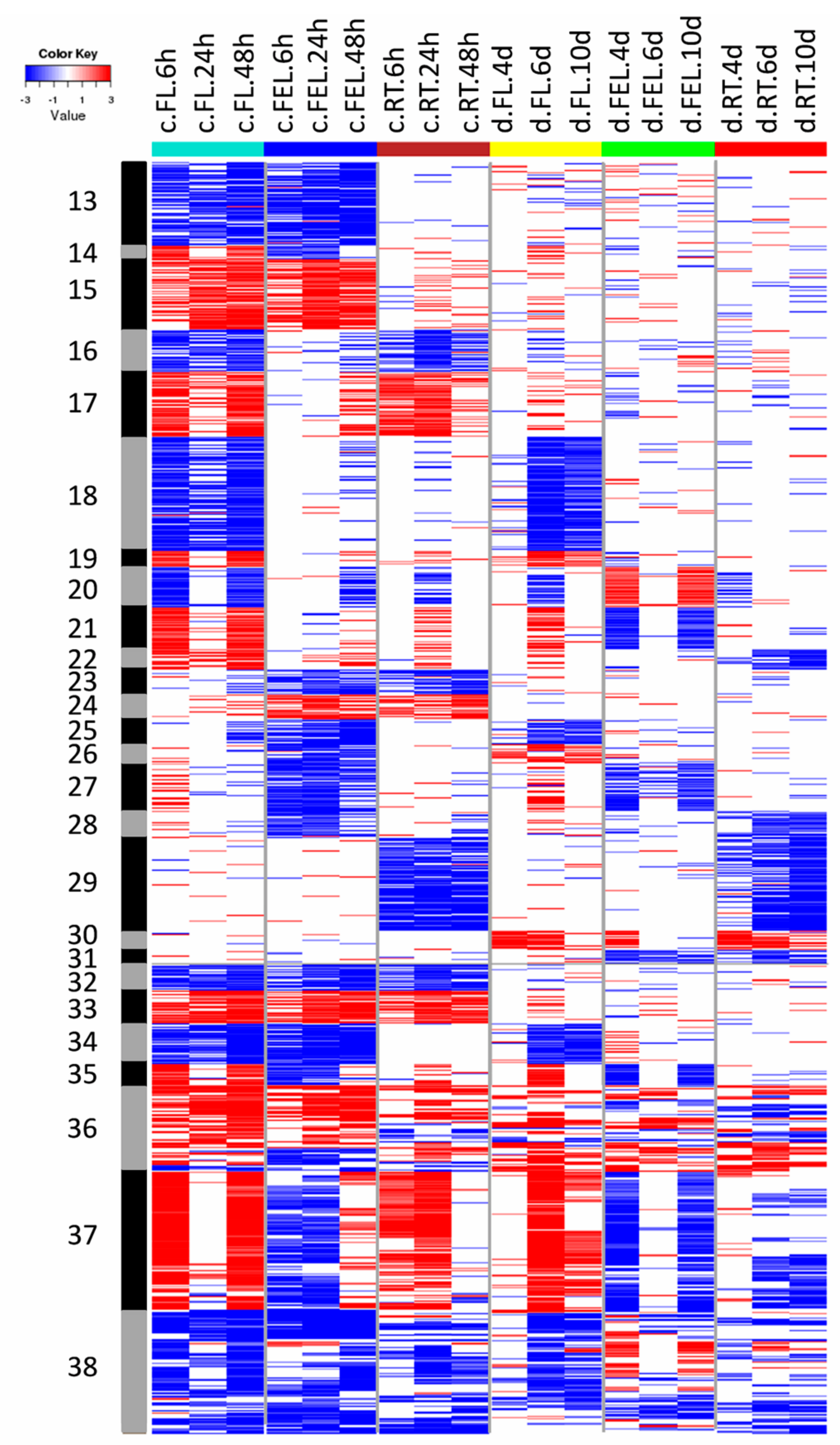
2.4. DE Genes Specifically Responding to Drought
The expression of genes from groups G7–G12 was significantly changed in only one tissue in response to drought (
Figure 2). The expression of genes of G7 and G8 was significantly decreased and increased in FL, respectively. The enriched categories in G7 included cell wall and light reaction of photosynthesis, and those in G8 included protein synthesis and abiotic stress (
Figure 3), which was quite different to those of cold responses in the same tissue. Similarly, the gene expression of G9 and G10 was significantly decreased and increased in FEL, respectively. No enriched categories were found in these two groups (
Figure 3). The expression of 259 genes in G11 and 156 in G12 was significantly decreased and increased in RT, respectively (
Figure 2). The genes in G11 were enriched in ethylene metabolism and secondary metabolism, but no enriched categories were found in G12 (
Figure 3).
In addition to genes that were differentially expressed in only one tissue, many genes whose expression was significantly changed in more than one tissue were also observed exclusively in response to drought. For example, the expression of genes in G30 was significantly increased in FL and RT under drought, and these genes mainly participated in abiotic stress (
Figure 3 and
Figure 4). The expression of genes in G31 was significantly decreased in FEL and RT under drought, but no enriched categories were observed (
Figure 3 and
Figure 4).
Together, the results indicated that DE genes, which exclusively responded to drought, were mainly involved in cell wall, protein synthesis, light reaction of photosynthesis, abiotic stress, and ethylene metabolism.
2.5. DE Genes Responding to Both Cold and Drought
Many genes that showed significantly changed expression in both cold and drought were also observed. The expression of genes in G18 and G19 was significantly decreased and increased, respectively, in FL under both cold and drought (
Figure 4). Category enrichment showed that these genes were involved in cell wall and DNA synthesis/chromatic structure, and co-factor and vitamin metabolism, respectively (
Figure 3). In addition, groups of genes whose expression was changed under both cold and drought stresses were also observed in FEL (e.g., G27) and RT (e.g., G29) (
Figure 4). The enriched categories of G29 included hormone metabolism of ABA and SA, and FA synthesis and FA elongation, but no enriched categories were observed in G27 (
Figure 3). Interestingly, the expression of photosynthesis-related genes was significantly decreased in response to both cold and drought, although there were different expression patterns of these genes (e.g., G25, G34, and G38) (
Figure 4). In addition, the expression of genes involved in Ca
2+ signaling and abiotic stress (e.g., G37) was also significantly changed in response to both cold and drought.
In the following subsections, a few special and interesting pathways were further inspected to illustrate their respective responses to cold and drought stresses.
2.6. Photosynthesis and Carbohydrate Metabolism
Genes related to photosynthesis, including both light reaction and Calvin cycle, were dramatically influenced by either cold or drought, or both of these stresses (
Figure 3). It is noteworthy that the expression of all photosystem I and II DE genes was significantly decreased in leaves (
Table S2). Consistently, DE genes related to electron carrier (e.g., ferredoxin-NADP
+ oxidoreductase 2), cyclic electron flow-chlororespiration, and ATP synthase of light reaction were also decreased (
Table S2), although they exhibited stress-specific response patterns (e.g., cyclic electron flow-chlororespiration for cold and ATP synthase for drought). Likely, the expression of Calvin-cycle-related genes, such as three rubisco small subunit, four glyceraldehyde 3-phosphate dehydrogenase (two
GAPA and two
GAPB), two fructose-1,6-bisphosphatase (
FBPase), two sedoheptulose bisphosphatase (
SBPase), and two phosphoribulokinase (
PRK), was significantly decreased in response to cold and drought (
Table S2). In contrast to light reaction and Calvin cycle, the expression of only a few genes related to photorespiration was changed (
Table S2).
Tetrapyrroles play vital roles in photosynthesis and respiration. There are three branches, including the chlorophyll branch, siroheme branch, and heme branch, in the tetrapyrrole biosynthesis pathway in plants. However, only the expression of genes located in the chlorophyll branch was significantly influenced, especially in FL and FEL in response to cold, in which the expression of magnesium chelatase and protochlorophyllide reductase was dramatically decreased (
Table S2). These results suggested that photosynthesis metabolism was greatly influenced by tetrapyrrole biosynthesis through the chlorophyll branch, specifically in response to cold.
Starch and sucrose metabolisms are closely linked to photosynthesis. As expected, the expression of starch and sucrose metabolism genes was greatly depressed by cold and drought. For example, the expression of ADP-glucose pyrophosphorylase (
AGPase) and starch synthase (
SS) in starch biosynthesis [
32], as well as alpha amylase, beta amylase, and glycosyl transferase in the starch degradation pathway [
32] was significantly decreased in response to cold and drought (
Table S2). Similar changes were observed in sucrose-degradation-related genes such as fructokinase, hexokinase, sucrose synthase (
SUS), and sucrose transporters (
SUT) (
Table S2).
Together, the results indicated that cold and drought dramatically depressed the expression of photosynthesis genes mainly through reduced accumulation of chlorophyll and then inhibited the processes of carbohydrate metabolism.
2.7. Response of Abiotic-Stress-Related Genes
In total, 78 genes related to abiotic stress were identified in response to either cold or drought after manual curation. The most dominant category was related to heat stress (53 genes), followed by drought/salt (18 genes), cold stress (five genes), and touch/wounding (two genes,
Table S3).
Hierarchical clustering analysis grouped these genes into four main clusters (
Figure S4). Overall, the expression of genes in cluster A1 (14%, 11 genes) was significantly decreased under both stresses (except for c.FL), and the expression of genes in cluster A3 (33%, 26 genes) was dramatically increased, especially in the early response to drought.
ERD4, which encoded an early-responsive to dehydration stress protein [
33], was included in cluster A1, while most genes in cluster A3 were related to heat stress (
Table S3). In comparison, the expression of genes in cluster A2 (32%, 25 genes) was significantly decreased under cold but increased under drought stress, whereas cluster A4 (21%, 16 genes) showed opposite trends, except for those induced in FL under drought stress. As expected, several dehydration-responsive genes, including
ERD3,
QUL2, and
DI19 [
34,
35], were included in these two clusters. These results suggested that abiotic-stress-related genes had different responses to cold and drought.
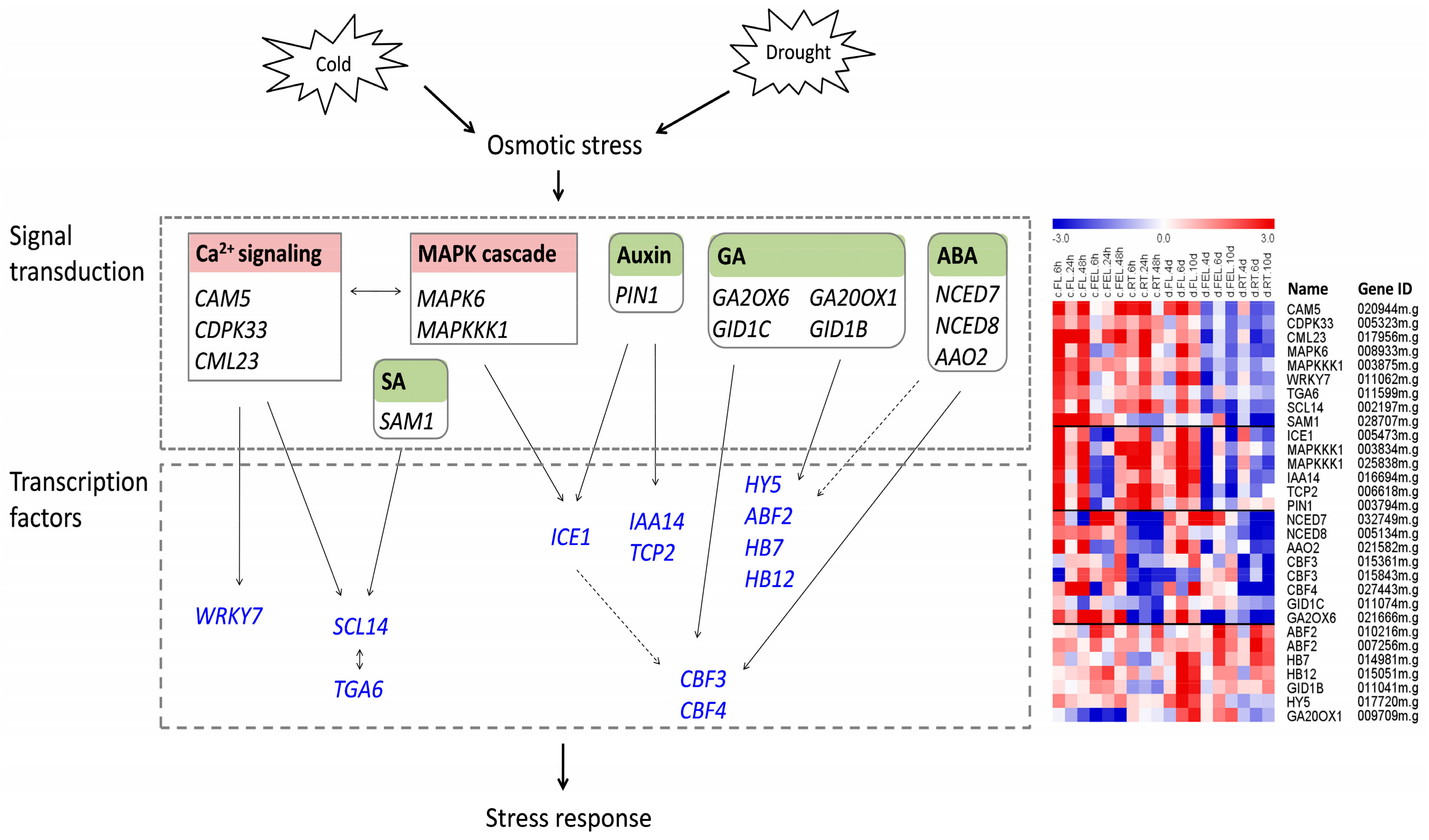
2.8. Response of Genes Related to Ca2+ Signaling, MAPK, and ROS
Ca
2+ sensors, MAPKs, and ROS-scavenging enzymes are important secondary messengers involved in abiotic stresses [
2,
5,
7], thus it is of great interest to determine their expression changes in response to cold and drought in cassava. In total, 13 Ca
2+ sensor-related genes, including three
CaMs, six
CMLs, one
CBL, and three
CDPKs, were identified (
Table S4). Four of them were responsive to both cold and drought, while the remaining nine were exclusively responsive to cold. Interestingly, most of the genes that responded to cold were up-regulated, whereas those responsive to drought were down-regulated (
Table S4). A total of 13
MAPKs, including six
MAPKs and seven
MAPKKK members, were identified. Similar to Ca
2+ sensors, most
MAPKs were differentially expressed in response to cold, or both cold and drought, but very few exclusively responded to drought (
Table S4). In total, 10 ROS-scavenging enzymes, including two superoxide dismutase (
SOD), two catalase (
CAT), four ascorbate peroxidase (
APX), and two glutathione reductase (
GR), were identified. Unlike Ca
2+ sensors and
MAPKs, most ROS-scavenging enzymes exclusively responded to cold or drought, and only one responded to both of these stresses (
Table S4). Together, these results implied that Ca
2+ sensors, MAPKs, and ROS-scavenging enzymes played important roles in cassava abiotic stress responses, but with different emphases for cold and drought.
2.9. Roles of Genes Related to Hormone Biosynthesis
Hormones usually play important roles in plant developmental processes and adaptation to changing environments. In this study, a total of 111 hormone-related genes, referring to ABA, auxin, BR, ethylene, gibberellic acid (GA), jasmonic acid (JA), SA, and cytokinin (CK), were differentially expressed in response to cold and drought. The top four hormones with most abundant genes were ethylene (35 genes), auxin (27), ABA (13), and GA (13), followed by BR (11) and SA (7) (
Table S5).
To reveal the role of hormones responding to cold and drought, the expression of genes related to biosynthesis of each hormone was examined, respectively. Nine-
cis-epoxycarotenoid dioxygenase (NCED) and ABA-aldehyde oxidase (AAO) are two key enzymes in the ABA biosynthetic pathway [
9]. The expression of
NCED4 and
NCED5 was significantly decreased in FL and FEL under cold stress, while the expression of
NCED7,
NCED8, and
AAO2 was significantly decreased in RT under both cold and drought. Besides this, the expression of
AAO4 was greatly induced in FEL exclusively in response to cold (
Table S5).
In the auxin biosynthetic pathway, the expression of
ILL6 (IAA-leucine resistant like gene 6), which encodes IAA-amino acid conjugate hydrolase involved in generating IAA from the hydrolysis of IAA-amino acid conjugates, was significantly depressed in RT under cold stress (
Table S5). Consistently,
PIN1, which encodes an auxin efflux carrier, showed a very similar expression trend. In addition, the expression of
PIN3 was also decreased, although it preferentially responded to cold in FL and FEL.
The expression of three genes
DWF1,
DWF5, and
DWF7, which are involved in different steps of the BR biosynthesis pathway [
36], was dramatically decreased in FL and FEL under cold stress (
Table S5).
ACC synthetase (ACS) and ACC oxidase (ACO) are two key enzymes that catalyze the final two steps of ethylene biosynthesis [
37]. Although the expression of
ACS8,
ACO4, and
ACO5 was greatly depressed, the former two responded to both cold and drought in FL and FEL, whereas
ACO5 preferentially responded to cold in RT.
The expression of GA biosynthetic genes, including copalyl diphosphate synthase (CPS) and GA20OX1, was greatly induced in response to cold and drought. In contrast, the expression of JA biosynthetic genes, including lipoxygenase (LOX), allene oxide synthase (AOS), and oxophytodienoate reductase (OPR), was all significantly depressed, although LOX2 and OPR3 preferentially responded to drought, and LOX6 and AOS1 responded to both cold and drought.
Together, these results suggested diverse roles of hormone genes in response to cold and drought stresses.
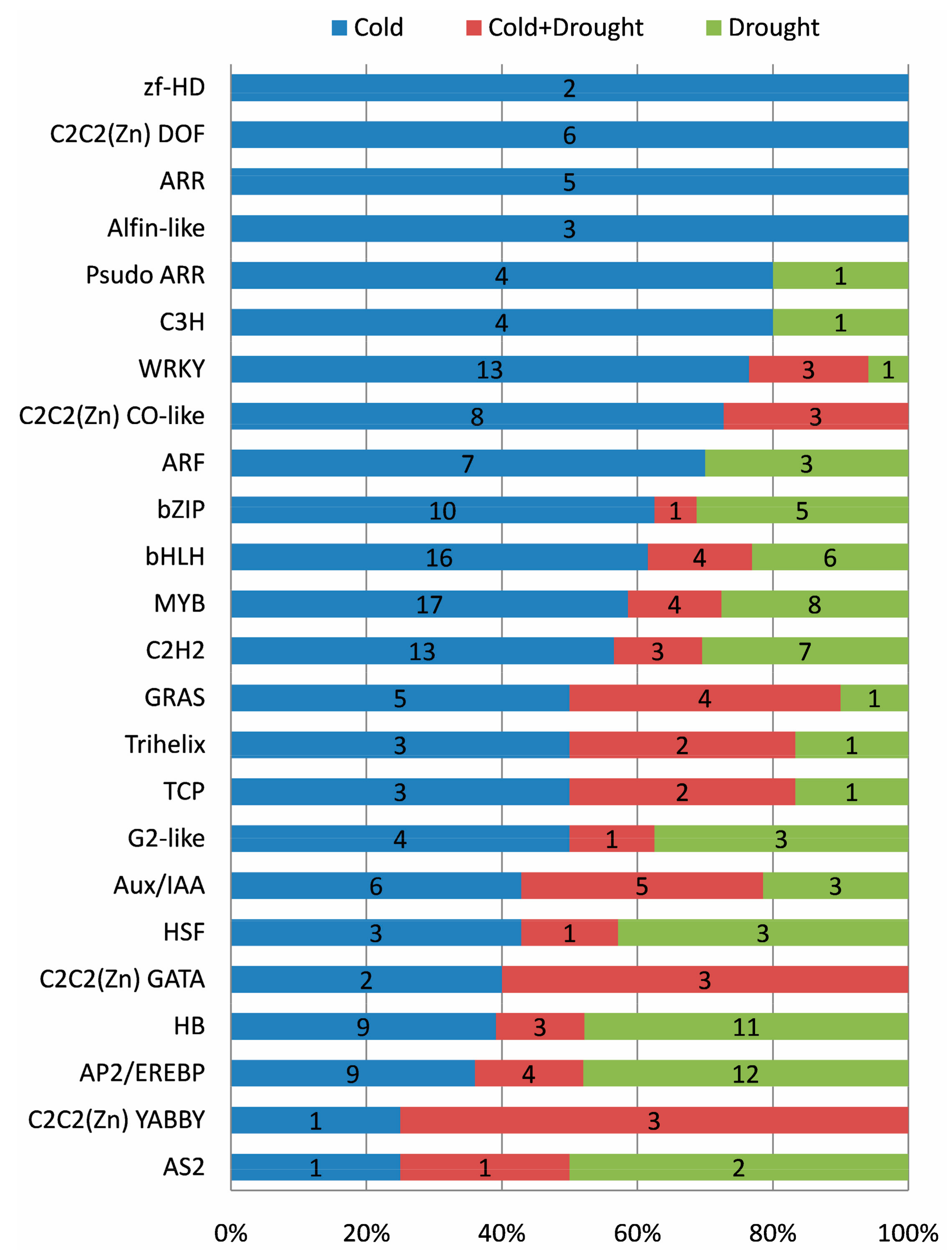
2.10. Roles of Transcription Factor (TF)
The expression changes of TF responding to cold and drought were well resolved in our RNA-seq data. In total, 1810 TF genes were expressed in at least one sample, and 270 (14.9%, representing 24 families) of them were differentially expressed in response to cold and drought stresses (
Table S6). The three most abundant TF families were MYB (29 genes), bHLH (26), and AP2/EREBP (25), followed by HB (23), C2H2 (23), WRKY (17), and bZIP (16). Of the 270 differentially expressed TFs, most (57.0%) of them were exclusively in response to cold, whereas 25.6% were specifically identified in drought, and about 17.4% were identified in both cold and drought (
Table S6).
Stress-specific expression trends were also inspected in each TF family, respectively. All members of zf-HD, C2C2 (Zn) DOF, ARR, and Alfin-like families were identified to exclusively respond to cold (
Figure 5), of which,
ARR1 and
ARR12 were involved in low temperature mediated inhibition of root growth by reducing auxin accumulation. Moreover, four out of five DE genes from the ARR family were involved in CK signaling (
Table S6), suggesting that CK played major roles in response to cold stress in cassava.
Most members from Psudo ARR, C3H, WRKY, C2C2(Zn) CO-like, ARF, bZIP, bHLH, and MYB families preferentially responded to cold (
Figure 5), of which,
PRR7 from Psudo ARR family was an essential component contributing to a temperature-sensitive circadian system;
WRKY39 functioned as a positive regulator in response to heat stress through SA- and JA-activated signaling pathways;
GBF3 from the bZIP family was involved in both cold and drought responses;
AIF4,
bHLH1, and
ICE1 from the bHLH family were involved in cold or freezing tolerance response; and
RVE1 and
MYB14 from the MYB family participated in the regulation of cold tolerance or cold acclimation (
Table S6). In addition, multiple TF families involved in hormones were also identified, e.g., C3H family (
AtTZF2 and
AtTZF3), WRKY family (
WRKY33 and
WRKY40), bZIP family (
GBF3 and
ABI5), and bHLH family (
JAM2,
AKS2, and
AKS3) for ABA; WRKY family (
WRKY33) for ethylene; WRKY family (
WRKY51 and
WRKY70) and bHLH family (
JAM2) for JA; and WRKY family (
WRKY4 and
WRKY70) for SA (
Table S6), suggesting that these hormones played major roles in response to cold stress.
In comparison, majority of the members of the HB, AP2/EREBP, and AS2 families preferentially responded to drought stress, whereas most members of the C2C2 (Zn) GATA and C2C2 (Zn) YABBY families responded to both cold and drought (
Figure 5). Accordingly, we found that
HB7 and
HB12 from the HB family participated in drought response via an ABA-dependent manner, and
SHN1,
RAP2.4,
ANT,
CBF4, and
ABR1 from the AP2/EREBP family were involved in drought response through hormones such as ethylene, ABA, and auxin. Notably, an AP2/EREBP member (
CBF3) previously reported to be involved in low temperature and ABA response, was specifically differentially expressed in RT under drought stress in our study (
Table S6).
Overall, the results revealed that TF members (even within the same family) exhibited exclusive and common expression responses to cold and drought, which might be regulated through hormone signaling.
4. Materials and Methods
4.1. Plant Materials and Treatments
Stem segments with three nodes were extracted from 10-month-old cassava cv. SC124 plants, and inclined in 3 L pots filled with barren red soil/vermiculite (1:1,
v/v) fertilized with Hoagland’s solution [
57], to propagate and generate uniform cassava seedlings for experiments. The soil was supplied with 300 mL of normal-strength Hoagland’s solution once a week. The plants grew in natural conditions (light/dark of 13/11 h and 27–33/22–30 °C) for 60 days, and then uniform seedlings were tagged and subjected to cold and drought treatments.
For cold treatment, plants were transferred to an illuminated incubator at 24 °C for 2 days for a homogenous start, and temperature was then decreased from 24 to 14 °C at the rate of −2 °C/h to exert a moderate chilling stress. Temperature was then held constant at 14 °C for 5 days for chilling acclimation. After 5 days of chilling acclimation and growth at 14 °C, plants were watered once again with Hoagland’s solution, and cooled further to 4 °C at a rate of −2 °C/h, and cultivated at constant 4 °C for 5 days. Then, folded leaves (FL, the first three folded leaves), fully expanded leaves (FEL, three most newly fully expanded leaves), and fibrous roots (RT) were sampled from three individual plants at 6, 24, and 48 h, respectively. In parallel, plants grown under normal condition of 24 °C at the three corresponding time points served as controls.
For drought treatment, watering of uniform plants was stopped for 5 days to stimulate the acclimation effects. At 6 days, the plants were watered once more with Hoagland’s solution, and then watering was stopped for another 10 days (i.e., the second phase) for the drought treatment. After 4, 6, and 10 days of the second phase of drought treatment, FL, FEL, and RT were harvested respectively from three individual plants. At the corresponding times, the same tissues of control plants (maintained at 90% full field water content by supplying the amount of water determined from weight loss) were also sampled. All samples were immediately frozen in liquid nitrogen and stored at −80 °C.
4.2. RNA-Seq Library Preparation and Sequencing
Total RNA was isolated separately for each sample as previously described. The integrity and quality of the total RNA were examined using a Nanodrop ND-1000 spectrophotometer (Thermo Scientific Inc., Waltham, MA, USA) and an Agilent 2100 Bioanalyzer (Agilent, Santa, CA, USA). For each time point of the treatment, equal amounts of RNA of the same tissue from three individual plants were pooled for expression profiling. In total, 24 RNA-seq libraries were separately constructed and sequenced at Beijing Genomics Institute (Shenzhen, China) based on the Illumina GAII platform.
4.3. Data Analysis
Differentially expressed (DE) genes were identified through two steps. (1) For each tissue (which included one control and three treatment time points) in either cold or drought stress, we used similar previously reported criteria [
29] to determine whether a gene was a DE gene candidate. For example, given two samples of one treatment and one control, a gene was considered as a DE gene candidate if (a) the fold-change (FC) of CPM was >3 if the gene was expressed in two samples, or (b) the gene was not expressed (CPM < 10) in one sample but over-expressed (CPM > 30) in the other sample. (2) DE gene candidates that were significant in at least two of three time points during the first step were selected and finally considered as DE genes in response to stress in that tissue. This step could be considered as a complement to the lack of replicates, which used different time points instead.
To understand the gene interactions between cold and drought stresses, and among different tissues (e.g., FL, FEL, and RT), heatmaps were used to visualize the FC of gene expression, which were grouped by hierarchical clustering (with “complete” method according to the Euclidean distance) implemented in MeV [
61]. The FCs were all set to 1 for non-DE genes. For gene functional category enrichment, cassava loci were functionally annotated and classified into hierarchical categories based on MapMan [
62], and then significantly over-represented functional categories were identified based on Fisher’s exact test as previously reported [
30,
58].
4.4. Validation of Commonly and Specifically Expressed Genes Responding to Cold and Drought Treatments
To validate the results of RNA-seq, seven genes of interest, related to primary cell wall process, stress responsive transcriptional factors, and protein modification and degradation, were selected and confirmed by using qRT-PCR with the primers listed in
Table S1. The amplification programs were performed on an ABI PRISM 7900HT real-time PCR system. The amplification conditions were 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s. A thermal denaturing step that generated the melt curves was followed after the qRT-PCR cycles to verify the amplification specificity. Beta-actin was used as the endogenous control. Each sample was measured in triplicate, including its non-template control. The relative gene expression level across the samples was calculated by using the 2
–△△Ct method.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





