
Commitment of Scaffold Proteins in the Onco-Biology of Human Colorectal Cancer and Liver Metastases after Oxaliplatin-Based Chemotherapy

 ,
,  , and
, and 
Abstract
:
1. Introduction
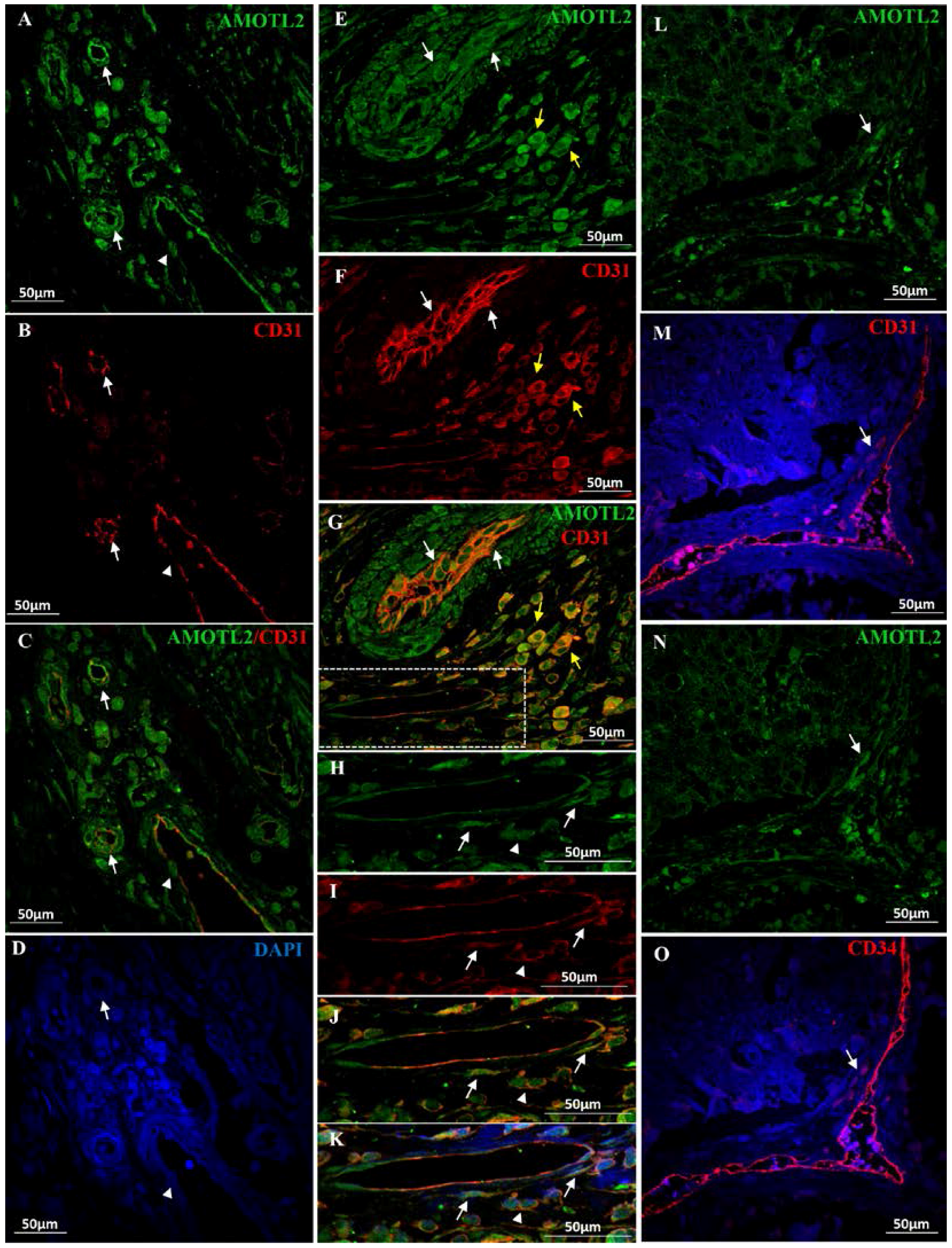
2. Results
2.1. AmotL2 in Healthy Colon and in CRC Tissue Samples
2.2. AmotL2 in Healthy Liver and in CRC Metastasized Liver Tissue Samples
2.3. AmotL2 in Blood Vessels
2.4. FKBP51
2.5. IQGAP1
3. Discussion
3.1. AmotL2 Expression in Healthy Colon and in CRC
3.2. AmotL2 Expression in Healthy Colon, Liver and in CRC-Metastasized Liver
3.3. FKBP51 Expression in Healthy Colon, Liver, CRC and Metastasized Liver
3.4. IQGAP1 Expression in Healthy Colon, Liver, CRC and Metastasized Liver
3.5. Final Considerations and Future Research Directions
4. Materials and Methods
4.1. Patients, Tumor Tissue and Controls
4.2. Antibodies
4.3. Immunohistochemistry
4.4. Double Immunofluorescence Simultaneous Staining
4.5. Image Analysis and Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Amot | Angiomotin |
| APC | Adenomatous polyposis coli |
| CLIP-170 | Cytoplasmic linker protein CLIP-170 |
| CT | Chemotherapy |
| EMT | Epithelial mesenchymal transition |
| FKBP | FK506 binding protein |
| FOLFOX | FOL—Folinic acid, leucovorin, F—Fluorouracil, 5-FU, OX—Oxaliplatin |
| HSP90 | Heat shock protein 90 |
| IQGAP1 | IQ-motif containing GTPase activating protein 1 |
| MAPK | Mitogen-activated protein kinase |
| PCNA | Proliferating cell nuclear antigen |
| TEAD | Transcriptional enhancer factors |
| YAP | Yes-associated protein |
References
- Bendris, N.; Schmid, S.L. Endocytosis, metastasis and beyond: Multiple facets of SNX9. Trends Cell Biol. 2017, 27, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Garcia, E.; O’Bryan, J.P. Intersectin scaffold proteins and their role in cell signaling and endocytosis. Biochim. Biophys. Acta 2017, 1864, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Garbett, D.; Bretscher, A. The surprising dynamics of scaffolding proteins. Mol. Biol. Cell 2014, 25, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Calvo, M.; Concha, A.; Figueroa, A.; Garrido, F.; Valladares-Ayerbes, M. Colorectal Cancer Classification and Cell Heterogeneity: A Systems Oncology Approach. Int. J. Mol. Sci. 2015, 16, 13610–13632. [Google Scholar] [CrossRef] [PubMed]
- Trinh, A.; Trumpi, K.; De Sousa, E.M.; Wang, X.; de Jong, J.H.; Fessler, E.; Kuppen, P.J.; Reimers, M.S.; Swets, M.; Koopman, M.; et al. Practical and Robust Identification of Molecular Subtypes in Colorectal Cancer by Immunohistochemistry. Clin. Cancer Res. 2017, 23, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Andre, T.; Boni, C.; Mounedji-Boudiaf, L.; Navarro, M.; Tabernero, J.; Hickish, T.; Topham, C.; Zaninelli, M.; Clingan, P.; Bridgewater, J.; et al. fluorouracil, and leucovorin as adjuvant treatment for colon cancer. N. Engl. J. Med. 2004, 350, 2343–2351. [Google Scholar] [CrossRef] [PubMed]
- Pachman, D.R.; Loprinzi, C.L.; Grothey, A.; Ta, L.E. The search for treatments to reduce chemotherapy-induced peripheral neuropathy. J. Clin. Investig. 2014, 124, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Seo, A.N.; Kim, H. Sinusoidal obstruction syndrome after oxaliplatin-based chemotherapy. Clin. Mol. Hepatol. 2014, 20, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Avila, J.; Gonzalez-Fernandez, R.; Boronat, L.; Soriano, M.L.; Martin-Vasallo, P. Differential transcriptome profile of peripheral white cells to identify biomarkers involved in oxaliplatin induced neuropathy. J. Pers. Med. 2014, 4, 282–296. [Google Scholar] [CrossRef] [PubMed]
- Bratt, A.; Wilson, W.J.; Troyanovsky, B.; Aase, K.; Kessler, R.; Van Meir, E.G.; Holmgren, L. Angiomotin belongs to a novel protein family with conserved coiled-coil and PDZ binding domains. Gene 2002, 298, 69–77. [Google Scholar] [CrossRef]
- Mojallal, M.; Zheng, Y.; Hultin, S.; Audebert, S.; van, H.T.; Johnsson, P.; Lenander, C.; Fritz, N.; Mieth, C.; Corcoran, M.; et al. AmotL2 disrupts apical-basal cell polarity and promotes tumour invasion. Nat. Commun. 2014, 5, 4557. [Google Scholar] [CrossRef] [PubMed]
- Erlejman, A.G.; De Leo, S.A.; Mazaira, G.I.; Molinari, A.M.; Camisay, M.F.; Fontana, V.; Cox, M.B.; Piwien-Pilipuk, G.; Galigniana, M.D. NF-κB transcriptional activity is modulated by FK506-binding proteins FKBP51 and FKBP52: A role for peptidyl-prolyl isomerase activity. J. Biol. Chem. 2014, 19, 26263–26276. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.E. Peptidyl-prolyl isomerases: A new twist to transcription. EMBO Rep. 2002, 3, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Hubler, T.R.; Denny, W.B.; Valentine, D.L.; Cheung-Flynn, J.; Smith, D.F.; Scammell, J.G. The FK506-binding immunophilin FKBP51 is transcriptionally regulated by progestin and attenuates progestin responsiveness. Endocrinology 2003, 144, 2380–2387. [Google Scholar] [CrossRef] [PubMed]
- Barik, S. Immunophilins: For the love of proteins. Cell. Mol. Life Sci. 2006, 63, 2889–2900. [Google Scholar] [CrossRef] [PubMed]
- Galigniana, M.D.; Radanyi, C.; Renoir, J.M.; Housley, P.R.; Pratt, W.B. Evidence that the peptidylprolyl isomerase domain of the hsp90-binding immunophilin FKBP52 is involved in both dynein interaction and glucocorticoid receptor movement to the nucleus. J. Biol. Chem. 2001, 276, 14884–14889. [Google Scholar] [CrossRef] [PubMed]
- Rein, T. FK506 binding protein 51 integrates pathways of adaptation: FKBP51 shapes the reactivity to environmental change. Bioessays 2016, 38, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Mukaide, H.; Adachi, Y.; Taketani, S.; Iwasaki, M.; Koike-Kiriyama, N.; Shigematsu, A.; Shi, M.; Yanai, S.; Yoshioka, K.; Kamiyama, Y.; et al. FKBP51 expressed by both normal epithelial cells and adenocarcinoma of colon suppresses proliferation of colorectal adenocarcinoma. Cancer Investig. 2008, 26, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Rotoli, D.; Morales, M.; Del Carmen, M.M.; Del Pino, G.M.; Morales, A.; Avila, J.; Martin-Vasallo, P. Expression and localization of the immunophilin FKBP51 in colorectal carcinomas and primary metastases, and alterations following oxaliplatin-based chemotherapy. Oncol. Lett. 2016, 12, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Toneatto, J.; Guber, S.; Charo, N.L.; Susperreguy, S.; Schwartz, J.; Galigniana, M.D.; Piwien-Pilipuk, G. Dynamic mitochondrial-nuclear redistribution of the immunophilin FKBP51 is regulated by the PKA signaling pathway to control gene expression during adipocyte differentiation. J. Cell Sci. 2013, 126, 5357–5368. [Google Scholar] [CrossRef] [PubMed]
- Storer, C.L.; Dickey, C.A.; Galigniana, M.D.; Rein, T.; Cox, M.B. FKBP51 and FKBP52 in signaling and disease. Trends Endocrinol. Metab. 2011, 22, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Pei, H.; Li, L.; Fridley, B.L.; Jenkins, G.D.; Kalari, K.R.; Lingle, W.; Petersen, G.; Lou, Z.; Wang, L. FKBP51 affects cancer cell response to chemotherapy by negatively regulating Akt. Cancer Cell 2009, 16, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; D’Angelillo, A.; D’Arrigo, P.; Staibano, S.; Greco, A.; Brunetti, A.; Scalvenzi, M.; Bisogni, R.; Scala, I.; Romano, M.F. FKBP51 increases the tumour-promoter potential of TGF-β. Clin. Transl. Med. 2014, 3, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Gallo, L.I.; Lagadari, M.; Piwien-Pilipuk, G.; Galigniana, M.D. The 90-kDa heat-shock protein (Hsp90)-binding immunophilin FKBP51 is a mitochondrial protein that translocates to the nucleus to protect cells against oxidative stress. J. Biol. Chem. 2011, 286, 30152–30160. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lou, Z.; Wang, L. The role of FKBP5 in cancer aetiology and chemoresistance. Br. J. Cancer 2011, 104, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Abel, A.M.; Schuldt, K.M.; Rajasekaran, K.; Hwang, D.; Riese, M.J.; Rao, S.; Thakar, M.S.; Malarkannan, S. IQGAP1: Insights into the function of a molecular puppeteer. Mol. Immunol. 2015, 65, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Erickson, J.W.; Cerione, R.A.; Hart, M.J. Identification of an actin cytoskeletal complex that includes IQGAP and the Cdc42 GTPase. J. Biol. Chem. 1997, 272, 24443–24447. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Li, Z.; Sacks, D.B. IQGAP1 is a scaffold for mitogen-activated protein kinase signaling. Mol. Cell. Biol. 2005, 25, 7940–7952. [Google Scholar] [CrossRef] [PubMed]
- Malarkannan, S.; Awasthi, A.; Rajasekaran, K.; Kumar, P.; Schuldt, K.M.; Bartoszek, A.; Manoharan, N.; Goldner, N.K.; Umhoefer, C.M.; Thakar, M.S. IQGAP1: A regulator of intracellular spacetime relativity. J. Immunol. 2012, 188, 2057–2063. [Google Scholar] [CrossRef] [PubMed]
- White, C.D.; Erdemir, H.H.; Sacks, D.B. IQGAP1 and its binding proteins control diverse biological functions. Cell Signal. 2012, 24, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Sharma, M.; Henderson, B.R. IQGAP1 regulation and roles in cancer. Cell Signal. 2009, 21, 1471–1478. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, K.; Shimao, Y.; Inoue, T.; Koono, M. Immunohistochemical analysis of IQGAP1 expression in human colorectal carcinomas: Its overexpression in carcinomas and association with invasion fronts. Cancer Lett. 2002, 176, 101–109. [Google Scholar] [CrossRef]
- Rotoli, D.; Perez-Rodriguez, N.D.; Morales, M.; Maeso, M.D.; Avila, J.; Mobasheri, A.; Martin-Vasallo, P. IQGAP1 in podosomes/invadosomes is involved in the progression of glioblastoma multiforme depending on the tumor status. Int. J. Mol. Sci. 2017, 18, E150. [Google Scholar] [CrossRef] [PubMed]
- Rotoli, D.; Morales, M.; del Carmen, M.M.; del Pino, G.M.; Gutierrez, R.; Valladares, F.; Avila, J.; Díaz-Flores, L.; Mobasheri, A.; Martin-Vasallo, P. Alterations in IQGAP1 expression and localization in colorectal carcinomas and liver metastases following oxaliplatin based chemotherapy. Oncol. Lett. 2016, 12, 1315–1322. [Google Scholar] [PubMed]
- White, C.D.; Brown, M.D.; Sacks, D.B. IQGAPs in cancer: A family of scaffold proteins underlying tumorigenesis. FEBS Lett. 2009, 583, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Lu, F.I.; Jia, S.; Meng, S.; Cao, Y.; Wang, Y.; Ma, W.; Yin, K.; Wen, Z.; Peng, J.; et al. AmotL2 is essential for cell movements in zebrafish embryo and regulates c-Src translocation. Development 2007, 134, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Chun, A.; Cheung, K.; Rashidi, B.; Yang, X. Tumor suppressor LATS1 is a negative regulator of oncogene YAP. J. Biol. Chem. 2008, 283, 5496–5509. [Google Scholar] [CrossRef] [PubMed]
- Porazinski, S.; Wang, H.; Asaoka, Y.; Behrndt, M.; Miyamoto, T.; Morita, H.; Hata, S.; Sasaki, T.; Krens, S.F.; Osada, Y.; et al. YAP is essential for tissue tension to ensure vertebrate 3D body shape. Nature 2015, 521, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Hong, W. Angiomotin’g YAP into the nucleus for cell proliferation and cancer development. Sci. Signal. 2013, 6, e27. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Ye, X.; Yu, J.; Li, L.; Li, W.; Li, S.; Yu, J.; Lin, J.D.; Wang, C.Y.; Chinnaiyan, A.M.; et al. TEAD mediates YAP-dependent gene induction and growth control. Genes Dev. 2008, 22, 1962–1971. [Google Scholar] [CrossRef] [PubMed]
- Landin-Malt, A.; Benhaddou, A.; Zider, A.; Flagiello, D. An evolutionary, structural and functional overview of the mammalian TEAD1 and TEAD2 transcription factors. Gene 2016, 591, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the maestro of the replication fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Bratt, A.; Birot, O.; Sinha, I.; Veitonmaki, N.; Aase, K.; Ernkvist, M.; Holmgren, L. Angiomotin regulates endothelial cell-cell junctions and cell motility. J. Biol. Chem. 2005, 280, 34859–34869. [Google Scholar] [CrossRef] [PubMed]
- Troyanovsky, B.; Levchenko, T.; Mansson, G.; Matvijenko, O.; Holmgren, L. Angiomotin: An angiostatin binding protein that regulates endothelial cell migration and tube formation. J. Cell Biol. 2001, 19, 1247–1254. [Google Scholar] [CrossRef]
- Shenoy, A.K.; Jin, Y.; Luo, H.; Tang, M.; Pampo, C.; Shao, R.; Siemann, D.W.; Wu, L.; Heldermon, C.D.; Law, B.K.; et al. Epithelial-to-mesenchymal transition confers pericyte properties on cancer cells. J. Clin. Investig. 2016, 126, 4174–4186. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Flores, L.; Gutierrez, R.; Madrid, J.F.; Varela, H.; Valladares, F.; Acosta, E.; Martin-Vasallo, P.; Diaz-Flores, L., Jr. Pericytes. Morphofunction, interactions and pathology in a quiescent and activated mesenchymal cell niche. Histol. Histopathol. 2009, 24, 909–969. [Google Scholar] [PubMed]
- Popescu, L.M.; Faussone-Pellegrini, M.S. TELOCYTES—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Flores, L.; Gutierrez, R.; Garcia, M.P.; Saez, F.J.; Aparicio, F.; Diaz-Flores, L., Jr.; Madrid, J.F. Uptake and intracytoplasmic storage of pigmented particles by human CD34+ stromal cells/telocytes: Endocytic property of telocytes. J. Cell. Mol. Med. 2014, 18, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Smythies, J. Intercellular signaling in cancer-the SMT and TOFT hypotheses, exosomes, telocytes and metastases: Is the messenger in the message? J. Cancer 2015, 6, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Shen, Z.; Stemmer-Rachamimov, A.; Dawany, N.; Troutman, S.; Showe, L.C.; Liu, Q.; Shimono, A.; Sudol, M.; Holmgren, L.; et al. The p130 isoform of angiomotin is required for Yap-mediated hepatic epithelial cell proliferation and tumorigenesis. Sci. Signal. 2013, 6, ra77. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Ortiz, A.; Shen, P.F.; Cheng, C.J.; Lee, Y.C.; Yu, G.; Lin, S.C.; Creighton, C.J.; Yu-Lee, L.Y.; Lin, S.H. Angiomotin regulates prostate cancer cell proliferation by signaling through the Hippo-YAP pathway. Oncotarget 2016, 8, 10145–10160. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.H.; Cho, H.D.; Lee, J.H.; Lee, H.J.; Hong, S.A.; Cho, J.; Kim, H.J.; Oh, M.H. Reduced angiomotin p130 expression correlates with poor prognosis in lung adenocarcinoma. BMJ J. 2016. [Google Scholar] [CrossRef]
- Nalbantoglu, I.L.; Tan, B.R., Jr.; Linehan, D.C.; Gao, F.; Brunt, E.M. Histological features and severity of oxaliplatin-induced liver injury and clinical associations. J. Dig. Dis. 2014, 15, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.H.; Ning, Y.M.; Sanchez, E.R. A new first step in activation of steroid receptors: Hormone-induced switching of FKBP51 and FKBP52 immunophilins. J. Biol. Chem. 2002, 277, 4597–4600. [Google Scholar] [CrossRef] [PubMed]
- Wochnik, G.M.; Ruegg, J.; Abel, G.A.; Schmidt, U.; Holsboer, F.; Rein, T. FK506-binding proteins 51 and 52 differentially regulate dynein interaction and nuclear translocation of the glucocorticoid receptor in mammalian cells. J. Biol. Chem. 2005, 280, 4609–4616. [Google Scholar] [CrossRef] [PubMed]
- Sidler, D.; Renzulli, P.; Schnoz, C.; Berger, B.; Schneider-Jakob, S.; Fluck, C.; Inderbitzin, D.; Corazza, N.; Candinas, D.; Brunner, T. Colon cancer cells produce immunoregulatory glucocorticoids. Oncogene 2011, 30, 2411–2419. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Mallardo, M.; Romano, M.F. FKBP51 and the NF-κB regulatory pathway in cancer. Curr. Opin. Pharmacol. 2011, 11, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Fukata, M.; Watanabe, T.; Noritake, J.; Nakagawa, M.; Yamaga, M.; Kuroda, S.; Matsuura, Y.; Iwamatsu, A.; Perez, F.; Kaibuchi, K. Rac1 and Cdc42 capture microtubules through IQGAP1 and CLIP-170. Cell 2002, 109, 873–885. [Google Scholar] [CrossRef]
- Watanabe, T.; Wang, S.; Noritake, J.; Sato, K.; Fukata, M.; Takefuji, M.; Nakagawa, M.; Izumi, N.; Akiyama, T.; Kaibuchi, K. Interaction with IQGAP1 links APC to Rac1, Cdc42, and actin filaments during cell polarization and migration. Dev. Cell. 2004, 7, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.A.; Henderson, B.R. The scaffolding protein IQGAP1 co-localizes with actin at the cytoplasmic face of the nuclear envelope: Implications for cytoskeletal regulation. Bioarchitecture 2012, 2, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Bielak-Zmijewska, A.; Kolano, A.; Szczepanska, K.; Maleszewski, M.; Borsuk, E. CDC42 protein acts upstream of IQGAP1 and regulates cytokinesis in mouse oocytes and embryos. Dev. Biol. 2008, 322, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Laorden, B.; Viros, A.; Marais, R. Mind the IQGAP. Cancer Cell 2013, 23, 715–717. [Google Scholar] [CrossRef] [PubMed]
- Tekletsadik, Y.K.; Sonn, R.; Osman, M.A. A conserved role of IQGAP1 in regulating TOR complex 1. J. Cell Sci. 2012, 125, 2041–2052. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka-Tojo, M.; Tojo, T.; Kim, H.W.; Hilenski, L.; Patrushev, N.A.; Zhang, L.; Fukai, T.; Ushio-Fukai, M. IQGAP1 mediates VE-cadherin-based cell–cell contacts and VEGF signaling at adherence junctions linked to angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1991–1997. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AmotL2 | IQGAP1 | FKBP51 | |
|---|---|---|---|
| Healthy Colon | |||
| Epithelial cells (Mucosae) | +/+++ | +++ | +++ |
| Stromal cells (Mucosae) | +++ | − | ++ |
| Immune system cells | + | ? | ++ |
| Blood vessel cells | +++ | ++ | − |
| Smooth muscle cells | +++ | ++ | + |
| Neurous (Myenteric plexus) | +++ | ++ | −/+ |
| Glia cells (Myenteric plexus) | +++ | ++ | −/+ |
| Colorectal Cancer | |||
| Tumor cells | +++ | −/+++ | −/++ |
| Budding tumor cells | +++ | −/+++ | −/+ |
| Tumor associated stromal cells | +++ | −/++ | −/+++ |
| Epithelial cells (Mucosae) | ++ | −/+++ | ++ |
| Immune system cells | +++ | +++ | +++ |
| Smooth muscle cells | ++ | − | ++ |
| Blood vessel cells | +++ | +++ | − |
| Neurous (Myenteric plexus) | ++ | − | ++ |
| Glia cells (Myenteric plexus) | ++ | − | ++ |
| Healthy Liver | |||
| Hepatocytes | +/+++ | −/+++ | −/++ |
| Epithelial cells (Bile duct) | + | + | − |
| Blood vessel cells | +++ | +++ | − |
| Immune system cells | − | + | + |
| Metastasized Liver | |||
| Tumor cells | +++ | −/++ | −/+ |
| Budding tumor cells | +++ | +++ | − |
| Hepatocytes | ++ | −/+++ | +/++ |
| Epithelial cells (Bile duct) | ++ | ++ | − |
| Immune system cells | +++ | +/++ | ++ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rotoli, D.; Morales, M.; Ávila, J.; Maeso, M.D.C.; García, M.D.P.; Mobasheri, A.; Martín-Vasallo, P. Commitment of Scaffold Proteins in the Onco-Biology of Human Colorectal Cancer and Liver Metastases after Oxaliplatin-Based Chemotherapy. Int. J. Mol. Sci. 2017, 18, 891. https://doi.org/10.3390/ijms18040891
Rotoli D, Morales M, Ávila J, Maeso MDC, García MDP, Mobasheri A, Martín-Vasallo P. Commitment of Scaffold Proteins in the Onco-Biology of Human Colorectal Cancer and Liver Metastases after Oxaliplatin-Based Chemotherapy. International Journal of Molecular Sciences. 2017; 18(4):891. https://doi.org/10.3390/ijms18040891
Chicago/Turabian StyleRotoli, Deborah, Manuel Morales, Julio Ávila, María Del Carmen Maeso, María Del Pino García, Ali Mobasheri, and Pablo Martín-Vasallo. 2017. "Commitment of Scaffold Proteins in the Onco-Biology of Human Colorectal Cancer and Liver Metastases after Oxaliplatin-Based Chemotherapy" International Journal of Molecular Sciences 18, no. 4: 891. https://doi.org/10.3390/ijms18040891
APA StyleRotoli, D., Morales, M., Ávila, J., Maeso, M. D. C., García, M. D. P., Mobasheri, A., & Martín-Vasallo, P. (2017). Commitment of Scaffold Proteins in the Onco-Biology of Human Colorectal Cancer and Liver Metastases after Oxaliplatin-Based Chemotherapy. International Journal of Molecular Sciences, 18(4), 891. https://doi.org/10.3390/ijms18040891