
State of Art of Cancer Pharmacogenomics in Latin American Populations

 ,
,  ,
, 
Abstract
:
1. Introduction
2. Somatic Genomics in Oncology
3. Germline Pharmacogenomics in Oncology
3.1. Single Nucleotide Polymorphisms
3.2. Drug Transporters
3.3. Drug Targets
3.4. Drug-Metabolizing Enzymes
3.5. Pharmacogenomics in Clinical Practice
4. Germline Cancer Predisposition in Latin America
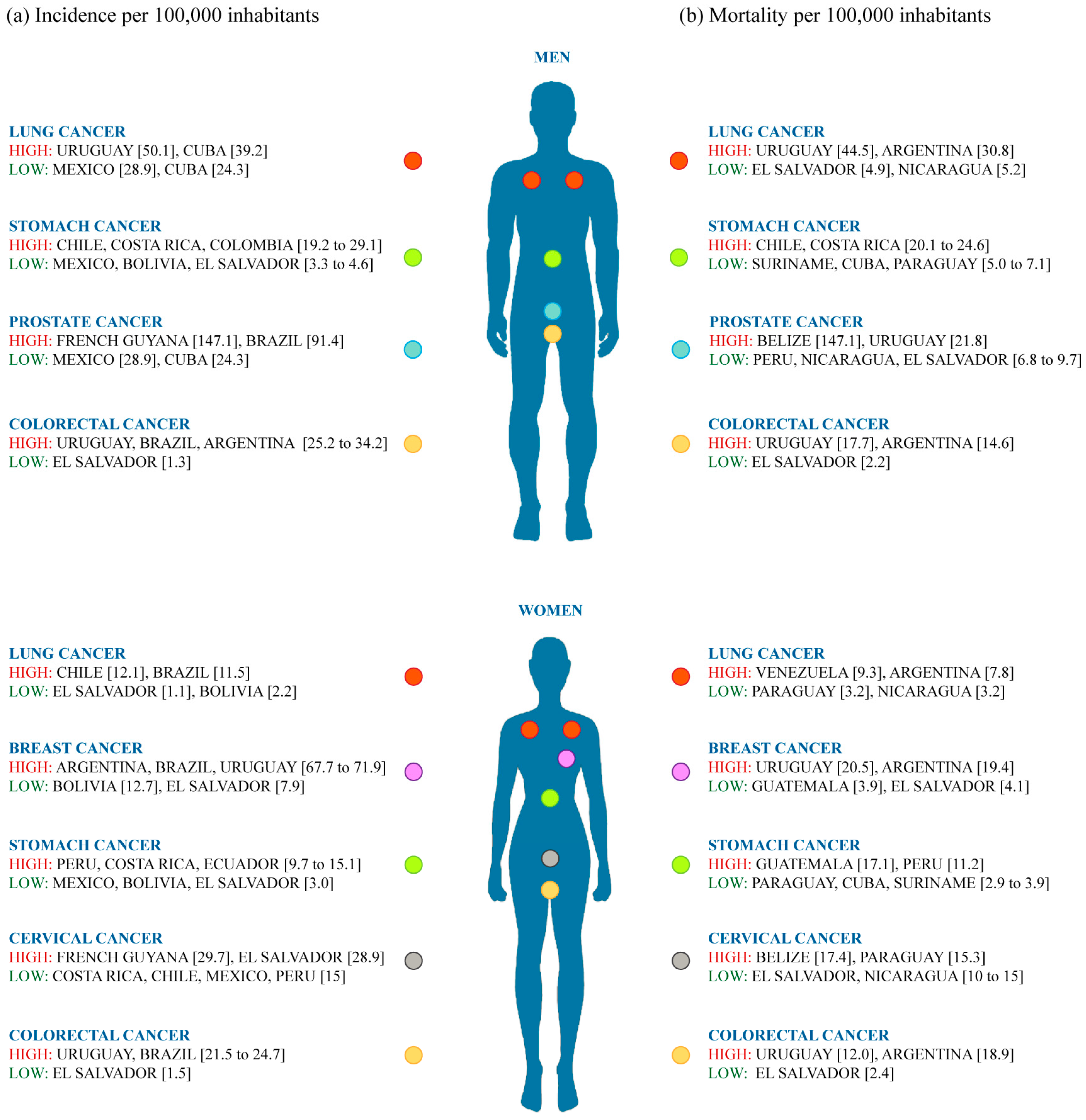
4.1. Cancer Incidence and Mortality in Latin America
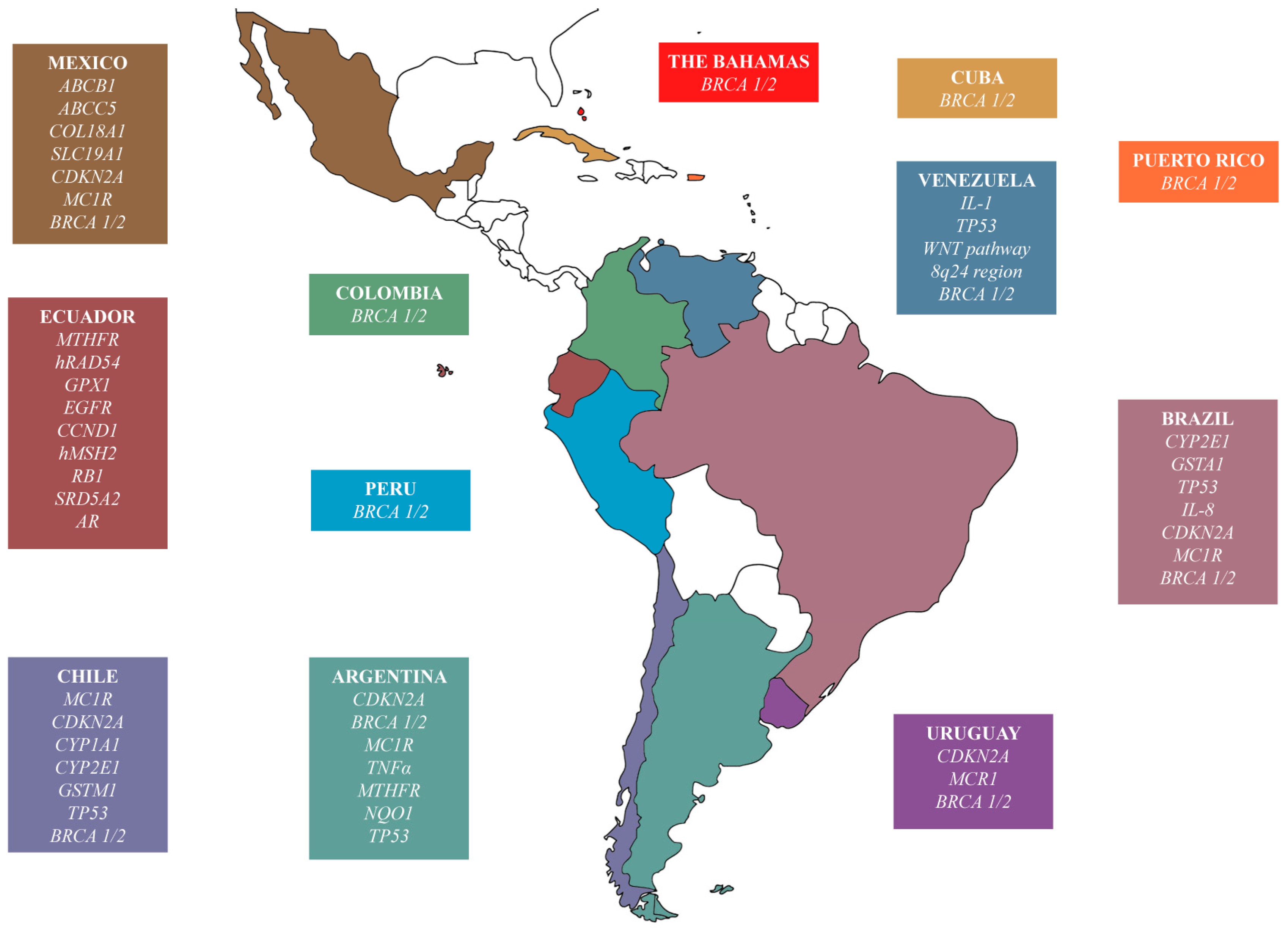
4.2. Genotyping the Latin American Populations with Cancer
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Sierra, M.; Forman, D. Cancer in Central and South America: Methodology. Cancer Epidemiol. 2016, 44, S11–S22. [Google Scholar] [CrossRef] [PubMed]
- McLeod, H.L. Cancer pharmacogenomics: Early promise, but concerned effort needed. Science 2013, 339, 1563–1566. [Google Scholar] [CrossRef] [PubMed]
- Gerlinger, M.; Rowan, A.J.; Horswell, S.; Larkin, J.; Endesfelder, D.; Gronroos, E.; Martinez, P.; Matthews, N.; Stewart, A.; Tarpey, P.; et al. Intratumor heterogeneity and branched evolution revealed by multiregion sequencing. N. Engl. J. Med. 2013, 366, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Wurth, L.; Panagiotis, P.; Olmeda, D.; Bley, N.; Calvo, G.; Guerrero, S.; Cerezo-Wallis, D.; Martinez-Useros, J.; García-Fernández, M.; Hüttelmaier, S.; et al. UNR/CSDE1 drives a post-transcriptional program to promote melanoma invasion and metastasis. Cancer Cell 2016, 30, 694–707. [Google Scholar] [CrossRef] [PubMed]
- Hertz, D.; McLeod, H.L. Use of pharmacogenetics for predicting cancer prognosis and treatment exposure, response and toxicity. J. Hum. Genet. 2013, 58, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.B.; Hui, E.P.; Mok, T.S. Population-based differences in treatment outcome following anticancer drug therapies. Lancet Oncol. 2010, 11, 75–84. [Google Scholar] [CrossRef]
- Collins, D.C.; Sundar, R.; Lim, J.S.J.; Yap, T.A. Towards precision medicine in the clinic: From biomarker discovery to novel therapeutics. Trends Pharmacol. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Rocco, G.; Morabito, A.; Leone, A.; Muto, P.; Fiore, F.; Budillon, A. Management of non-small cell lung cancer in the era of personalized medicine. Int. J. Biochem. Cell Biol. 2016, 78, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.S.W.; Mok, T.S.K.; Rebbeck, T.R. Cancer genomics: Diversity and disparity across ethnicity and geography. J. Clin. Oncol. 2016, 34, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhang, Y.; Parra, E.; Rodriguez, J.; Behrens, C.; Akbani, R.; Lu, Y.; Kurie, J.M.; Ginnons, D.L.; Mills, G.B.; et al. Multiplatform-based molecular subtypes of non-small-cell lung cancer. Oncogene 2016. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, P.F.; Arnoletti, J.P.; Benson, A.B.; Chen, Y.J.; Choti, M.A.; Cooper, H.S.; Covey, A.; Dilawari, R.A.; Early, D.S.; Enzinger, P.C.; et al. NCCN clinical practice guidelines in oncology: Colon cancer. J. Natl. Compr. Cancer Netw. 2009, 7, 778–831. [Google Scholar]
- Grothey, A.; Allegra, C.J. Antiangiogenesis therapy in the treatment of metastatic colorectal cancer. Ther. Adv. Med. Oncol. 2012, 4, 301–319. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.; Meade, A.; Wasan, H.; Griffiths, G.; Maughan, T. Cetuximab therapy in first-line metastatic colorectal cancer and intermittent palliative chemotherapy: Review of the COIN trial. Expert Rev. Anticancer Ther. 2008, 8, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Amado, R.G.; Wolf, M.; Peeters, M.; van Cutsem, E.; Siena, S.; Freeman, D.J.; Juan, T.; Sikorski, R.; Suggs, S.; Radinsky, R.; et al. Wild-type KRAS is required for panitumumab efficacy in patients with metastatic colorectal cancer. J. Clin. Oncol. 2008, 26, 1626–1634. [Google Scholar] [CrossRef] [PubMed]
- Karapetis, C.S.; Khambata-Ford, S.; Jonker, D.J.; O’Callaghan, C.J.; Tu, D.; Tebbutt, N.C.; Simen, R.J.; Chalchal, H.; Shapiro, J.D.; Robitaille, S.; et al. K-Ras mutations and benefit from cetuximab in advanced colorectal cancer. N. Engl. J. Med. 2008, 359, 1757–1765. [Google Scholar] [CrossRef] [PubMed]
- Barbacid, M. Ras genes. Annu. Rev. Biochem. 1987, 56, 779–827. [Google Scholar] [CrossRef] [PubMed]
- Macara, I.G.; Lounsbury, K.M.; Richards, S.A.; McKiernan, C.; Bar-Sagi, D. The Ras superfamily of GTPases. FASEB J. 1996, 10, 625–630. [Google Scholar] [PubMed]
- Cárdenas-Ramos, S.G.; Alcázar-González, G.; Reyes-Cortés, L.M.; Torres-Grimaldo, A.A.; Calderón-Garcidueñas, A.L.; Morales-Casas, J.; Flores-Sánchez, P.; de León-Escobedo, R.; Gómez-Díaz, A.; Moreno-Bringas, C.; et al. The frequency and type of K-RAS mutations in Mexican patients with colorectal cancer. Am. J. Clin. Oncol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Rodenhuis, S.; Wetering, M.; Mooi, W.; Evers, S.; Zandwijk, N.; Bos, J. Mutational activation of the K-Ras oncogene. N. Engl. J. Med. 1987, 317, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Zocche, D.; Ramirez, C.; Fontao, F.; Costa, L.; Redal, M. Global impact of KRAS mutation patterns in FOLFOX treated metastatic colorectal cancer. Front. Genet. 2015, 6, 116. [Google Scholar] [CrossRef] [PubMed]
- Sameer, A.S.; Chowdhri, N.A.; Abdullah, S.; Shah, Z.A.; Siddiqi, M.A. Mutation pattern of K-Ras gene in colorectal cancer patients of Kashmir: A report. Indian J. Cancer 2009, 46, 219–225. [Google Scholar] [PubMed]
- Elsamany, S.A.; Alzahrani, A.S.; Mohamed, M.M.; Elmorsy, S.A.; Zekri, J.E.; Al-Shehri, A.S.; Haggag, R.M.; El Taani, H.A. Clinico-pathological patterns and survival outcome of colorectal cancer in young patients: Western Saudi Arabia experience. Asian Pac. J. Cancer Prev. 2014, 15, 5239–5243. [Google Scholar] [CrossRef] [PubMed]
- Schubbert, S.; Shannon, K.; Bollag, G. Hyperactive Ras in developmental disorders and cancer. Nat. Rev. Cancer 2007, 7, 295–308. [Google Scholar] [CrossRef] [PubMed]
- De Roock, W.; de Vriendt, V.; Normanno, N.; Ciardiello, F.; Tejpar, S. KRAS, BRAF, PIK3CA, and PTEN mutations: Implications for targeted therapies in metastatic colorectal cancer. Lancet Oncol. 2011, 12, 594–603. [Google Scholar] [CrossRef]
- Douillard, J.Y.; Olineret, K.S.; Siena, S.; Tabernero, J.; Burkes, R.; Barugel, M.; Humblet, Y.; Bodoky, G.; Cunningham, D.; Jassem, J.; et al. Panitumumab-FOLFOX4 treatment and Ras mutations in colorectal cancer. N. Engl. J. Med. 2013, 369, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Akbani, R.; Akdemir, K.C.; Aksoy, B.A.; Albert, M.; Ally, A.; Amin, S.B.; Arachchi, H.; Arora, A.; Auman, J.T.; Ayala, B.; et al. Genomic classification of cutaneous melanoma. Cell 2015, 161, 1681–1696. [Google Scholar] [CrossRef] [PubMed]
- Alioto, T.; Buchhalter, I.; Derdak, S.; Hutter, B.; Eldridge, M.; Hoving, E.; Heisler, L.E.; Beck, T.A.; Simpson, J.T.; Tonon, L.; et al. A comprehensive assessment of somatic mutation detection in cancer using whole-genome sequencing. Nat. Commun. 2015, 6, 10001. [Google Scholar] [CrossRef] [PubMed]
- International Cancer Genome Consortium; Hudson, T.; Anderson, W.; Artez, A.; Barker, A.; Bell, C.; Bernabé, R.R.; Bhan, M.K.; Calvo, F.; Eerola, I.; et al. International network of cancer genome projects. Nature 2010, 464, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Carr, D.F.; Alfirevic, A.; Pirmohamed, M. Pharmacogenomics: Current state-of-the-art. Genes 2014, 5, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Relling, M.V.; Evans, W.E. Pharmacogenomics in the clinic. Nature 2015, 526, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.A.; Cavallari, L.H. Pharmacogenetics and cardiovascular disease—Implications for personalized medicine. Pharmacol. Rev. 2013, 65, 987–1009. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Phage therapy: Eco-physiological pharmacology. Scientifica 2014, 2014, 581639. [Google Scholar] [CrossRef] [PubMed]
- Nuchnoi, P.; Nantakomol, D.; Chumchua, V.; Plabplueng, C.; Isarankura-Na-Ayudhya, C. The identification of functional non-synonymous SNP in human ATPbinding cassette (ABC), subfamily member 7 gene: Application of bioinformatics tools in biomedicine. J. Bioanal. Biomed. 2011, 3, 26–31. [Google Scholar] [CrossRef]
- The 1000 Genomes Project Consortium. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [Green Version]
- Paz-y-Miño, C.; López-Cortés, A. Genética Molecular y Citogenética Humana: Fundamentos, Aplicaciones e Investigaciones en el Ecuador, 1st ed.; Yachay EP: Quito, Ecuador, 2014; p. 400. [Google Scholar]
- Wiwanitkit, V. Possible single nucleotide polymorphism (SNP) in the nucleic sequence of a-kinase-anchoring protein 9. J. Proteom. Bioinform. 2008, 1, 227–229. [Google Scholar] [CrossRef]
- Katara, P. Single nucleotide polymorphism and its dynamics for pharmacogenomics. Interdiscip. Sci. Comput. Life Sci. 2014, 6, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.M.; Prakash, N.; Pragya, Ch.; Krishna, M. Computational analysis of SNPs in 10 kb region of human chromosome 1. J. Comput. Sci. Syst. Biol. 2011, 4, 33–34. [Google Scholar] [CrossRef]
- Huang, T.; Shu, Y.; Cai, Y.D. Genetic differences among ethnic groups. BMC Genom. 2015, 16, 1093. [Google Scholar] [CrossRef] [PubMed]
- Reimer, T.; Kempert, S.; Gerber, B.; Hans-Jürgen, T.; Hartmann, S.; Koczan, D. SLCO1B1*5 polymorphism (rs4149056) is associated with chemotherapy-induced amenorrhea in premenopausal women with breast cancer: A prospective-cohort study. BMC Cancer 2016, 16, 337. [Google Scholar] [CrossRef] [PubMed]
- König, J. Pharmacogenomics of human OATP transporters. Arch. Pharmacol. 2006, 372, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Radtke, S.; Zolk, R.; Paulides, R.; Zimmermann, M.; Möricke, A.; Stanulla, M.; Schrappe, M.; Langer, T. Germline genetic variations in methotrexate candidate genes are associated with pharmacokinetics, toxicity, and outcome in childhood acute lymphoblastic leukemia. Blood 2013, 121, 5145–5153. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, L.; Johnson, S.; Caudle, K.; Haidar, C.; Voora, D.; Wilke, R.; Maxwell, W.; McLeod, H.; Krauss, R.; Roden, D.; et al. The clinical pharmacogenetics implementation consortium guideline for SLCO1B1 and simvastatin-induced myopathy: 2014 update. Clin. Pharmacol. Ther. 2014, 96, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, L.; Panetta, J.; Smith, C.; Yang, W.; Fan, Y.; Winick, N.; Martin, P.; Cheng, C.; Devidas, M.; Pui, C.; et al. Genome-wide study of methotrexate clearance replicates SLCO1B1. Blood 2013, 121, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, L.; Bruun, G.; Yang, W.; Treviño, L.; Vattathil, S.; Scheet, P.; Cheng, C.; Rosner, G.; Giacomini, K.; Fan, Y.; et al. Rare versus common variants in pharmacogenetics: SLCO1B1 variation and methotrexate disposition. Genome Res. 2012, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Grem, J.L. 5-Fluorouracil: Forty-plus and still ticking. A review of its preclinical and clinical development. Investig. New Drugs 2000, 18, 299–313. [Google Scholar] [CrossRef]
- Leichman, C.G.; Lenz, H.J.; Leichman, L.; Danenberg, K.; Baranda, J.; Groshen, S.; Boswell, W.; Metzger, R.; Tan, M.; Denenberg, P.V. Quantitation of intratumoral thymidylate synthase expression predicts for disseminated colorectal cancer response and resistance to protracted-infusion fluorouracil and weekly leucovorin. J. Clin. Oncol. 1997, 15, 3223–3229. [Google Scholar] [CrossRef] [PubMed]
- Pullarkat, S.T.; Stoehlmacher, J.; Ghaderi, V.; Xiong, Y.P.; Ingles, S.A.; Sherrod, A.; Warren, R.; Tsao-Wei, D.; Groshen, S.; Lenz, H.J. Thymidylate synthase gene polymorphism determines response and toxicity of 5-FU chemotherapy. Pharmacogenom. J. 2001, 1, 65–70. [Google Scholar] [CrossRef]
- Niedzwiecki, D.; Hasson, R.; Lenz, H.; Ye, C.; Redston, M.; Ogino, S.; Fuchs, C.; Compton, C.; Mayer, R.; Goldberg, R.; et al. A study of thymidylate synthase expression as a biomarker for resectable colon cancer: Alliance (cancer and leukemia group B) 9581 and 89803. Oncologist 2016. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cai, L.; Huang, H.; Liu, B.; Wu, Q. Genetic variations and haplotype diversity of the UGT1 gene cluster in the Chinese population. PLoS ONE. 2012, 7, e33988. [Google Scholar] [CrossRef] [PubMed]
- Kupiec, T.; Raj, V.; Vu, N. Pharmacogenomics for the forensic toxicologist. J. Anal. Toxicol. 2006, 30, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Lockhart, A.C.; Kim, R.B.; Rothenberg, M.L. Cancer pharmacogenomics: Powerful tools in cancer chemotherapy and drug development. Oncologist 2005, 10, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.E.; Horner, M.; Chu, Y.Q.; Kalwinsky, D.; Roberts, W.M. Altered mercaptopurine metabolism, toxic effects, and dosage requirement in a thiopurine methyltransferase-deficient child with acute lymphocytic leukemia. J. Pediatr. 1991, 119, 985–989. [Google Scholar] [CrossRef]
- Laróvere, L.; de Kremer, R.; Lambooy, L.; de Abreu, R. Genetic polymorphism of thiopurine S-methyltransferase in Argentina. Ann. Clin. Biochem. 2003, 40, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Collie-Duguid, E.S.; Pritchard, S.C.; Powrie, R.H.; Sludden, J.; Collier, D.A.; Li, T.; McLeod, H.L. The frequency and distribution of thiopurine methyltransferase alleles in Caucasian and Asian populations. Pharmacogenetics 1999, 9, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Yates, C.R.; Krynetski, E.Y.; Loennechen, T.; Fessing, M.Y.; Tai, H.L.; Pui, C.H.; Relling, M.V.; Evans, W.E. Molecular diagnosis of thiopurine S-methyltransferase deficiency: Genetic basis for azathioprine and mercaptopurine intolerance. Ann. Intern. Med. 1997, 126, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Rothenberg, M.L.; Kuhn, J.G.; Burris, H.A.; Nelson, J.; Eckardt, J.R.; Tristan-Morales, M.; Hilsenbeck, S.G.; Weiss, G.R.; Smith, L.S.; Rodríguez, G.I. Phase I and pharmacokinetic trial of weekly CPT-11. J. Clin. Oncol. 1993, 11, 2194–2204. [Google Scholar] [CrossRef] [PubMed]
- Gupta, E.; Lestingi, T.M.; Mick, R.; Ramirez, J.; Vokes, E.E.; Ratain, M.J. Metabolic fate of irinotecan in humans: Correlation of glucuronidation with diarrhea. Cancer Res. 1994, 54, 3723–3725. [Google Scholar] [PubMed]
- Bosma, P.J.; Chowdhury, J.R.; Bakker, C.; Gantla, S.; de Boer, A.; Oostra, B.A.; Lindhout, D.; Tytgat, G.N.; Jansen, P.L.; Oude Elfernik, R.P. The genetic basis of the reduced expression of bilirubin UDP-glucuronosyltransferase 1 in Gilbert’s syndrome. N. Engl. J. Med. 1995, 333, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, F.; Undevia, S.D.; Iyer, L.; Chen, P.X.; Das, S.; Kocherginsky, M.; Karrison, T.; Janisch, L.; Ramírez, J.; Rudin, C.M.; et al. Genetic variants in the UDP-glucuronosyltransferase 1A1 gene predict the risk of severe neutropenia of irinotecan. J. Clin. Oncol. 2004, 22, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.E. Pharmacogenomics: Marshalling the human genome to individualise drug therapy. Gut 2003, 52, ii10–ii18. [Google Scholar] [CrossRef] [PubMed]
- Relling, M.; Klein, T. CPIC: Clinical Pharmacogenetics Implementation Consortium of the pharmacogenomics research network. Clin. Pharmacol. Ther. 2011, 89, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Swen, J.; Nijenhuis, M.; de Boer, A.; Grandia, L.; Maitland-van der Zee, A.; Mulden, H.; Ronger, G.; van Schaik, R.; Schalekamp, T.; Touw, D.; et al. Pharmacogenomics: From bench to byte—An update of guidelines. Clin. Pharmacol. Ther. 2011, 89, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.; Visscher, H.; Sistonen, J.; Brunham, L.; Pussegoda, K.; Loo, T.; Rieder, M.; Koren, G.; Carleston, B.; Hyden, M.; et al. The Canadian Pharmacogenomics Network for Drug Safety: A model for safety pharmacology. Thyroid 2010, 20, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, L.A.; Lavanderos, M.A.; Cayun, J.P.; García-Martín, E.; Agúndez, J.A.; Cáceres, D.D.; Roco, A.M.; Morales, J.E.; Herrera, L.; Encina, G.; et al. Perception of the usefulness of drug/gene pairs and barriers for pharmacogenomics in Latin America. Curr. Drug Metab. 2014, 15, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Atun, R.; de Andrade, L.O.M.; Almeida, G.; Cotlear, D.; Dmytraczenko, T.; Frenz, P.; Garcia, P.; Gómez-Dantés, O.; Knaul, F.M.; Muntaner, C.; et al. Health-system reform and universal health coverage in Latin America. Lancet 2015, 385, 1230–1247. [Google Scholar] [CrossRef]
- Goss, P.E.; Lee, B.L.; Badovinac-Crnjevic, T.; Strasser-Weippl, K.; Chavarri-Guerra, Y.; Louis, J.S.; Villarreal-Garza, C.; Unger-Saldaña, K.; Ferreyra, M.; Debiasi, M.; et al. Planning cancer control in Latin America and the Caribbean. Lancet Oncol. 2013, 14, 391–436. [Google Scholar] [CrossRef]
- Sierra, M.; Soerjomataram, I.; Antoni, S.; Laversanne, M.; Piñeros, M.; de Vries, E.; Forman, D. Cancer patterns and trends in Central and South America. Cancer Epidemiol. 2016, 44, S23–S42. [Google Scholar] [CrossRef] [PubMed]
- Country Profile: Human Development Indicators. United Nations Development Programme (UNDP), 2013. Available online: http://hdr.undp.org/en/data/profiles/ (accessed on 13 March 2017).
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Forman, D.; Sierra, M. Cancer in Central and South America: Introduction. Cancer Epidemiol. 2016, 44, S3–S10. [Google Scholar] [CrossRef] [PubMed]
- Di Sibio, A.; Abriata, G.; Forman, D.; Sierra, M. Female breast cancer in Central and South America. Cancer Epidemiol. 2016, 44, S110–S120. [Google Scholar] [CrossRef] [PubMed]
- Sierra, M.; Soerjomataram, I.; Forman, D. Prostate cancer burden in Central and South America. Cancer Epidemiol. 2016, 44, S131–S140. [Google Scholar] [CrossRef] [PubMed]
- Piñeros, M.; Sierra, M.; Forman, D. Descriptive epidemiology of lung cancer and current status to tobacco control measures in Central and South America. Cancer Epidemiol. 2016, 44, S90–S99. [Google Scholar] [CrossRef] [PubMed]
- Sierra, M.; Cueva, P.; Bravo, L.E.; Forman, D. Stomach cancer burden in Central and South America. Cancer Epidemiol. 2016, 44, S62–S73. [Google Scholar] [CrossRef] [PubMed]
- Sierra, M.; Forman, D. Burden of colorectal cancer in Central and South America. Cancer Epidemiol. 2016, 44, S74–S81. [Google Scholar] [CrossRef] [PubMed]
- Murillo, R.; Herrero, R.; Sierra, M.; Forman, D. Cervical cancer in Central and South America: Burden of disease and status of disease control. Cancer Epidemiol. 2016, 44, S121–S130. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, L.; Lucas, D.; Godoy, J.; Cáceres, D.; Berthou, F.; Varela, N.; Lee, K.; Acevedo, C.; Martínez, L.; Aguilera, A.M.; et al. CYP1A1, CYP2E1 and GSTM1 genetic polymorphisms. The effect of single and combined genotypes on lung cancer susceptibility in Chilean people. Cancer Lett. 2001, 174, 35–44. [Google Scholar] [CrossRef]
- Cáceres, D.D.; Quiñones, L.A.; Schroeder, J.C.; Gil, L.D.; Irarrázabal, C.E. Association between p53 codon 72 genetic polymorphism and tobacco use and lung cancer risk. Lung 2009, 187, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, H.; Hamada, G.S.; Suzuki, I.; Iwase, T.; Kiyokawa, E.; Kino, I.; Tsugane, S. CYP1A1 and CYP2E1 polymorphism and lung cancer, case-control study in Rio de Janeiro, Brazil. Pharmacogenetics 1995, 5, S145–S148. [Google Scholar] [CrossRef] [PubMed]
- Paz-y-Miño, C.; López-Cortés, A.; Muñoz, M.J.; Cabrera, A.; Castro, B.; Sánchez, M.E. Incidence of the L858R and G719S mutations of the epidermal growth factor receptor oncogene in an Ecuadorian population with lung cancer. Cancer Genet. Cytogenet. 2010, 196, 201–203. [Google Scholar] [CrossRef] [PubMed]
- Midha, A.; Dearden, S.; McCormack, R. EGFR mutation incidence in non-small-cell lung cancer of adenocarcinoma histology: A systematic review and global map by ethnicity (mutMapII). Am. J. Cancer Res. 2015, 5, 2892–2911. [Google Scholar] [PubMed]
- Hung, RJ.; Boffeta, P.; Brockmöller, J.; Butkiewicz, D.; Cascorbi, I.; Clapper, M.; Garte, S.; Hauger, A.; Hirvonen, A.; Anttila, S.; et al. CYP1A1 and GSTM1 genetic polymorphisms and lung cancer risk in Caucasian non-smokers: A pooled analysis. Carcinogenesis 2003, 24, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Shaffi, S.; Shah, M.; Bhat, I.; Koul, P.; Ahmad, S.; Siddiqi, M. CYP1A1 polymorphisms and risk of lung cancer in the ethnic Kashmiri population. Asian Pac. J. Cancer Prev. 2009, 10, 651–656. [Google Scholar] [PubMed]
- Lui, X.; Li, Z.; Zhang, Z.; Zhang, W.; Li, W.; Xiao, Z.; Liu, H.; Jiao, H.; Wang, Y.; Li, G. Meta-analysis of GSTM1 null genotype and lung cancer in Asians. Med. Sci. Monit. 2014, 20, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lan, X.; Tan, S.; Wang, S.; Li, Y. P53 codon 72 Arg/Pro polymorphism and lung cancer risk Asians: As updated meta-analysis. Tumor Biol. 2013, 34, 2511–2520. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Bu, Z.; Feng, J.; Peng, L.; Liao, X.; Zhu, X.; Sun, X.; Yu, H.; Yan, D.; Yan, S. Association between the TP53 polymorphisms and lung cancer risk: A meta-analysis. Mol. Biol. Rep. 2014, 41, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Ze-Tian, S.; Xin-Hu, W.; Bing, L.; Jun-shu, S.; Zhen, W.; Jing, L.; Xi-Xi, Z. CYP2E1 Rsa I/Pst I polymorphism and lung cancer susceptibility: A meta-analysis involving 10,947 subjects. J. Cell. Mol. Med. 2015, 19, 2136–2142. [Google Scholar]
- Gonzalez-Hormazabal, P.; Gutierrez-Enriquez, S.; Gaete, D.; Reyes, J.M.; Peralta, O.; Waugh, E.; Gomez, F.; Margarit, S.; Bravo, T.; Diez, O.; et al. Spectrum of BRCA1/2 point mutations and genomic rearrangements in high-risk breast/ovarian cancer Chilean families. Breast Cancer Res. Treat. 2011, 126, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, M.; Silva, A.; Rubio, L.; Alvarez, C.; Torrealba, C.; Salinas, M.; Tapia, T.; Faundez, P.; Palma, L.; Riccio, M.E.; et al. Incidence of BRCA1 and BRCA2 mutations in 54 Chilean families with breast/ovarian cancer, genotype-phenotype correlations. Breast Cancer Res. Treat. 2006, 95, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Lara, K.; Consigliere, N.; Pérez, J.; Porco, A. BRCA1 and BRCA2 mutations in breast cancer patients from Venezuela. Biol. Res. 2012, 45, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.E.; Llacuachaqui, M.; Palacio, G.V.; Figueroa, J.D.; Madrid, J.; Lema, M.; Royer, R.; Li, S.; Larson, G.; Weitzel, J.N.; et al. Prevalence of BRCA1 and BRCA2 mutations in unselected breast cancer patients from Medellín, Colombia. Hered. Cancer Clin. Pract. 2014, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Garza, C.; Alvarez-Gómez, R.M.; Pérez-Plasencia, C.; Herrera, L.A.; Herzog, J.; Castillo, D.; Mohar, A.; Castro, C.; Gallardo, L.N.; Gallardo, D.; et al. Significant clinical impact of recurrent BRCA1 and BRCA2 mutations in Mexico. Cancer 2015, 121, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Solano, A.R.; Aceto, G.M.; Delettieres, D.; Veschi, S.; Neuman, M.I.; Alonso, E.; Chialina, S.; Chacón, R.D.; Renato, M.C.; Podestá, E.J. BRCA1 and BRCA2 analysis of Argentinean breast/ovarian cancer patients selected for age and family history highlights a role for novel mutations of putative south-American origin. Springerplus 2012, 25, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.C.; Costa, M.M.; Borojevic, R.; Monteiro, A.N.; Vieira, R.; Koifman, S.; Koifman, R.J.; Li, S.; Royer, R.; Zhang, S.; et al. Prevalence of BRCA1 and BRCA2 mutations in breast cancer patients from Brazil. Breast Cancer Res. Treat. 2007, 103, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.C.; Esperon, A.A.; Ropero, R.; Rubio, M.C.; Rodriguez, R.; Ortiz, R.M.; Anta, J.J.; de los Rios, M.; Carnesolta, D.; del Olivera, M.C.; et al. Prevalence of BRCA1 and BRCA2 mutations in breast cancer patients from Cuba. Fam. Cancer. 2008, 7, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Dutil, J.; Colon-Colon, J.L.; Matta, J.L.; Sutphen, R.; Echenique, M. Identification of the prevalent BRCA1 and BRCA2 mutations in the female population of Puerto Rico. Cancer Genet. 2012, 205, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Dutil, J.; Golubeva, V.A.; Pacheco-Torres, A.L.; Diaz-Zabala, H.J.; Matta, J.L.; Monteiro, A.N. The spectrum of BRCA1 and BRCA2 alleles in Latin America and the Caribbean: A clinical perspective. Breast Cancer Res. Treat. 2015, 154, 441. [Google Scholar] [CrossRef] [PubMed]
- Delgado, L.; Fernández, G.; Grotiuz, G.; Cataldi, S.; González, A.; Lluveras, N.; Heguaburu, M.; Fresco, R.; Lens, D.; Sabini, G.; et al. BRCA1 and BRCA2 germline mutations in Uruguayan breast and breast-ovarian cancer families. Identification of novel mutations and unclassified variants. Breast Cancer Res. Treat. 2011, 128, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Abugattas, J.; Llacuachaqui, M.; Allende, Y.S.; Velásquez, A.A.; Velarde, R.; Cotrina, J.; Garcés, M.; León, M.; Calderón, G.; de la Cruz, M.; et al. Prevalence of BRCA1 and BRCA2 mutations in unselected breast cancer patients from Peru. Clin. Genet. 2015, 88, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Buleje, J.L.; Huaman, F.; Guevara, M.; Acosta, O.; Pinto, J.A.; Araujo, J.; Ponce, J.; León, J.; Lizaraso, F.; Gómez, H.; et al. Detección de reordenamientos genómicos en los genes BRCA1 y BRCA2 en 16 familias peruanas con cáncer de mama mediante Amplificación de Sondas dependiente de Ligamiento Múltiple (MLPA). Carcinos 2015, 5, 34–38. [Google Scholar]
- Akbari, M.R.; Donenberg, T.; Lunn, J.; Curling, D.; Turnquest, T.; Krill-Jackson, E.; Zhang, S.; Narod, S.A.; Hurley, J. The spectrum of BRCA1 and BRCA2 mutations in breast cancer patients in the Bahamas. Clin. Genet. 2014, 85, 64–67. [Google Scholar] [CrossRef] [PubMed]
- López-Cortés, A.; Echeverría, C.; Oña-Cisneros, F.; Sánchez, M.E.; Herrera, C.; Cabrera-Andrade, A.; Rosales, F.; Ortiz, M.; Paz-y-Miño, C. Breast cancer risk associated with gene expression and genotype polymorphisms of the folate-metabolizing MTHFR gene: A case-control study in a high altitude Ecuadorian mestizo population. Tumor Biol. 2015, 36, 6451–6461. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Reid, J.; Burbidge, L.; Pruss, D.; Deffenbaugh, A.; Frye, C.; Wenstrup, R.; Ward, B.; Scholl, T.; Noll, W. BRCA1 and BRCA2 mutations in women of different ethnicities undergoing testing for hereditary breast-ovarian cancer. Cancer 2009, 115, 2222–2233. [Google Scholar] [CrossRef] [PubMed]
- Haeyoung, K.; Doo Ho, C. Distribution of BRCA1 and BRCA2 mutations in Asian patients with breast cancer. J. Breast Cancer 2013, 16, 357–365. [Google Scholar]
- Zhang, X.; Li, Y.; Yang, S.; Li, W. Meta-analysis on MTHFR polymorphism and lung cancer susceptibility in East Asian populations. Biomed. Rep. 2013, 1, 440–446. [Google Scholar] [PubMed]
- Chen, L.; Lu, N.; Zhang, B.; Weng, B.; Weng, L.; Lu, J. Association between the MTHFR C677T polymorphism and gastric cancer susceptibility: A meta-analysis of 5757 cases and 8501 controls. Oncol. Lett. 2015, 10, 1159–1165. [Google Scholar] [PubMed]
- Acevedo, C.; Opazo, J.L.; Huidobro, C.; Cabezas, J.; Iturrieta, J.; Quiñones Sepúlveda, L. Positive correlation between single or combined genotypes of CYP1A1 and GSTM1 in relation to prostate cancer in Chilean people. Prostate 2003, 57, 111–117. [Google Scholar] [CrossRef] [PubMed]
- López-Cortés, A.; Jaramillo-Koupermann, G.; Muñoz, M.J.; Cabrera, A.; Echeverría, C.; Rosales, F.; Vivar, N.; Paz-y-Miño, C. Genetic polymorphisms in MTHFR (C677T, A1298C), MTR (A2756G) and MTRR (A66G) genes associated with pathological characteristics of prostate cancer in the Ecuadorian population. Am. J. Med. Sci. 2013, 346, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Paz-y-Miño, C.; Witte, T.; Robles, P.; Llumipanta, W.; Díaz, M.; Arévalo, M. Association among polymorphisms in the steroid 5α-reductase type II (SRD5A2) gene, prostate cancer risk, and pathologic characteristics of prostate tumors in an Ecuadorian population. Cancer Genet. Cytogenet. 2009, 189, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Paz-y-Miño, C.; Robles, P.; Salazar, C.; Leone, P.E.; García-Cárdenas, J.M.; Naranjo, M.; López-Cortés, A. Positive association of the androgen receptor CAG repeat length polymorphism with the risk of prostate cancer. Mol. Med. Rep. 2016, 14, 1791–1798. [Google Scholar] [PubMed]
- Sá, R.A.; Moreira, A.D.S.; Cabello, P.H.; Ornellas, A.A.; Costa, E.B.; Matos, C.; Alves, G. Human glutathione S-transferase polymorphisms associated with prostate cancer in the Brazilian population. Int. Braz. J. Urol. 2014, 40, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Xu, W.; Liu, H.; Zhang, M.; Huang, Q.; Liao, Z. CYP1A1 MspI polymorphism is associated with prostate cancer susceptibility: Evidence from a meta-analysis. Mol. Biol. Rep. 2013, 40, 3483–3491. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Kazmi, Z.; Fatima, I.; Shabbir, R.; Perveen, S.; Masood, N. Genetic polymorphism of GSTM1 and GSTT1 and risk of prostatic carcinoma—A Meta-analysis of 7281 prostate cancer cases and 9082 healthy controls. Asian Pac. J. Cancer Prev. 2016, 17, 2629–2635. [Google Scholar] [PubMed]
- Chen, P.; Li, W.; Wang, J.; Jiang, Y.; Wu, P.; Chen, T.; Zheng, S. Association between MTHFR gene polymorphism (C677T, A1298C) and genetic susceptibility to prostate cancer: A meta-analysis. Genet. Mol. Res. 2015, 14, 19191–19202. [Google Scholar] [CrossRef] [PubMed]
- Neslund-Dudas, C.; Bock, C.; Monaghan, K.; Nock, N.; Yang, N.; Yang, J.; Rundle, A.; Tang, D.; Rybicki, B. SRD5A2 and HSD3B2 polymorphisms are associated with prostate cancer risk and aggressiveness. Prostate 2007, 67, 1654–1663. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Tao, W.; Chen, Q.; Hu, H.; Wen, X.; Han, R. SRD5A2 V89L polymorphism and prostate cancer risk: A meta-analysis. Prostate 2010, 70, 170–178. [Google Scholar] [PubMed]
- Gsur, A.; Preyer, M.; Haidinger, G.; Zidek, T.; Madersbacher, S.; Schatzl, G.; Marberger, M.; Vutuc, C.; Micksche, M. Polymorphic CAG repeats in the androgen receptor gene, prostate-specific antigen polymorphism and prostate cancer risk. Carcinogenesis 2002, 23, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Irvine, R.; Yu, M.; Ross, R.; Coetzee, G. The CAG and GGC microsatellites of the androgen receptor gene are in linkage disequilibrium in men with prostate cancer. Cancer Res. 1995, 55, 1937–1940. [Google Scholar] [PubMed]
- Komiya, Y.; Tsukino, H.; Nakao, H.; Kuroda, Y.; Imai, H.; Katoh, T. Human glutathione S-transferase A1, T1, M1, and P1 polymorphisms and susceptibility to prostate cancer in the Japanese population. J. Cancer Res. Clin. Oncol. 2005, 131, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Ning, B.; Wang, C.; More, F.; Nowell, S.; Ratnasinghe, D.; Carter, W.; Kadlubar, F.; Coles, B. Human glutathione S-transferase A2 polymorphisms: Variant expression, distribution in prostate cancer cases/controls and a novel form. Pharmacogenetics 2004, 14, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Chiurillo, M.A. Role of gene polymorphisms in gastric cancer and its precursor lesions: Current knowledge and perspectives in Latin American countries. World J. Gastroenterol. 2014, 20, 4503–4515. [Google Scholar] [CrossRef] [PubMed]
- Cañas, M.; Morán, Y.; Camargo, M.E.; Rivero, M.B.; Bohórquez, A.; Villegas, V.; Ramírez, E.; Rendón, Y.; Suárez, A.; Morales, L.; et al. TP53 codon 72 polymorphism and gastric cancer risk: A case-control study in individuals from the central-western region of Venezuela. Investig. Clin. 2009, 50, 153–161. [Google Scholar]
- Chiurillo, M.A. Role of the Wnt/β-catenin pathway in gastric cancer: An in-depth literature review. World J. Exp. Med. 2015, 5, 84–102. [Google Scholar] [CrossRef] [PubMed]
- Labrador, L.; Torres, K.; Camargo, M.; Santiago, L.; Valderrama, E.; Chiurillo, M.A. Association of common variants on chromosome 8q24 with gastric cancer in Venezuelan patients. Gene 2015, 566, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Felipe, A.V.; Silva, T.D.; Pimenta, C.A.; Kassab, P.; Forones, N.M. lnterleukin-8 gene polymorphism and susceptibility to gastric cancer in a Brazilian population. Biol. Res. 2012, 45, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Camargo, M.; Mera, R.; Correa, P.; Peek, R.; Fontham, E.; Goodman, K.; Piazuelo, M.; Sicinschi, L.; Zabaleta, J.; Schneider, B. Interleukin-1β and interleukin-1 receptor antagonista gene polymorphisms and gastric cancer: A meta-analysis. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1674–1687. [Google Scholar] [CrossRef] [PubMed]
- Peirce, L. Gastric cancer review. J. Carcinog. 2014, 13, 14. [Google Scholar]
- Mocellin, S.; Verdi, D.; Pooley, K.; Nitti, D. Genetic variation and gastric cancer risk: A field synopsis and meta-analysis. Gut 2015, 64, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Pan, H.; Hu, Y.; Zhu, Y.; He, Q. Polymorphism of IL-8 in 251 allele and gastric cancer susceptibility: A meta-analysis. Dig. Dis. Sci. 2010, 55, 1818–1823. [Google Scholar] [CrossRef] [PubMed]
- Puig, S.; Potrony, M.; Cuellar, F.; Puig-Butille, J.A.; Carrera, C.; Aguilera, P.; Nagore, E.; García-Casado, Z.; Requena, C.; Kumar, R.; et al. Characterization of individuals at high risk of developing melanoma in Latin America: Bases for genetic counseling in melanoma. Genet. Med. 2015, 18, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Hayward, N. Genetics of melanoma predisposition. Oncogene 2003, 22, 3053–3062. [Google Scholar] [CrossRef] [PubMed]
- Ibarrola-Villava, M.; Hu, H.-H.; Guedj, M.; Fernandez, L.; Descamps, V.; Basset-Seguin, N.; Bagot, M.; Benssussan, A.; Saiag, P.; Fargnoli, M.; et al. MC1R, SLC45A2 and TYR genetic variants involved in menaloma susceptibility in Southern European populations: Results from a meta-analysis. Eur. J. Cancer 2012, 48, 2183–2191. [Google Scholar] [CrossRef] [PubMed]
- Paz-y-Miño, C.; Muñoz, M.J.; López-Cortés, A.; Cabrera, A.; Palacios, A.; Castro, B.; Paz-y-Miño, N.; Sánchez, M.E. Frequency of polymorphisms pro198leu in GPX-1 gene and ile58thr in MnSOD gene in the altitude Ecuadorian population with bladder cancer. Oncol. Res. 2010, 18, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.E.; Wiencke, J.K.; Bates, M.N.; Zheng, S.; Rey, O.A.; Smith, A.H. Investigation of genetic polymorphisms and smoking in a bladder cancer case-control study in Argentina. Cancer Lett. 2004, 211, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, Y.; Habuchi, T.; Tsuchiya, N.; Wang, L.; Oyama, C.; Sato, K.; Nishiyama, H.; Ogawa, O.; Kato, T. Increased risk of bladder cancer associated with a glutathione peroxidase 1 codon 198 variant. J. Urol. 2004, 172, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Kucukgergin, C.; Sanli, O.; Amasyah, A.; Tefik, T.; Seckin, S. Genetic variants of MnSOD and GPX1 and susceptibility to bladder cancer in a Turkish population. Med. Oncol. 2012, 29, 1928–1934. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Dong, W.; An, R. Glutathione S-transferase T1 polymorphism contributes to bladder cancer risk: A meta-analysis involving 50 studies. DNA Cell Biol. 2012, 31, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Su, H.; Zhu, J.; Wang, D.; Zhu, S.; Li, S.; He, J.; Chen, Y. The association between NQO1 pro187ser polymorphism and bladder cancer susceptibility: A meta-analysis of 15 studies. PLoS ONE 2015, 10, e0116500. [Google Scholar] [CrossRef] [PubMed]
- Paz-y-Miño, C.; Salazar, C.; Zurita, T.; López-Cortés, A.; Hidalgo, R.; Rosales, F.; Montalvo, A.; Leone, P.E. Positive association between the polymorphic variant CCND1 A870G and colorectal cancer in Ecuadorian mestizo population. J. Cancer Res. Updat. 2015, 4, 163–170. [Google Scholar] [CrossRef]
- Pérez, L.O.; Abba, M.C.; Dulout, F.N.; Golijow, C.D. Evaluation of p53 codon 72 polymorphism in adenocarcinomas of the colon and rectum in La Plata, Argentina. World J. Gastroenterol. 2006, 12, 1426–1429. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yang, P.; Zhou, Y.; Wu, X. Cyclin D1 (CCND1) G870A gene polymorphism is an ethnicity-dependent risk factor for digestive tract cancers: A meta-analysis comprising 20,271 subjects. Cancer Epidemiol. 2012, 36, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Ni, X.; Yang, G.; Luo, Z.; Niu, Y.; Shen, M. CCND1 G870A polymorphism and colorectal cancer: An updated meta-analysis. Mol. Clin. Oncol. 2016, 4, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qin, H.; Zhang, Y.; Shi, T.; Liu, B.; Sun, Y.; Ma, Y. P53 codon 72 polymorphism and colorectal cancer: A meta-analysis of epidemiological studies. Hepatogastroenterology 2011, 58, 1926–1929. [Google Scholar] [PubMed]
- Economopoulos, K.; Sergentanis, T.; Zagouri, F.; Zografos, G. Association between p53 Arg72Pro polymorphism and colorectal cancer risk: A meta-analysis. Onkologie 2010, 33, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Rahal, M.; Herrera, M.J.; Quiñones, L.; Farfán, N.; Cáceres, D.; Roco, A. Frequency of CYP1A1*2a and GSTM1 gene polymorphisms in Chilean patients with squamous cell carcinoma of the larynx in relation to smoking habit: A pilot study. Rev. Otorrinolaringol. Cir. Cabeza Cuello 2013, 73, 7–16. [Google Scholar]
- Liu, X.; Fan, Q.; Ni, L.; Liu, F.; Huang, S.; Gao, J.; Chen, B. GSTM1 null genotype is a risk factor for laryngeal cancer. Int. J. Clin. Exp. Med. 2015, 8, 7661–7666. [Google Scholar] [PubMed]
- Zhuo, W.; Wang, Y.; Zhuo, X.; Zhu, B.; Zhu, Y.; Chen, Z. Polymorphisms of CYP1A1 and GSTM1 and laryngeal cancer risk: Evidence-based meta-analyses. J. Cancer Res. Clin. Oncol. 2009, 135, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Cordero, K.; Espinoza, I.; Cáceres, D.; Roco, A.; Miranda, C.; Squicciarini, V.; Santander, P.; Lee, K.; Saavedra, I.; Quiñones, L. Oral cancer susceptibility associated with the CYP1A1 and GSTM1 genotypes in Chilean individuals. Oncol. Lett. 2010, 1, 549–553. [Google Scholar] [PubMed]
- Cha, I.-H.; Park, J.; Chung, W.; Choi, M.; Kim, H.; Park, K. Polymorphisms of CYP1A1 and GSTM1 genes and susceptibility to oral cancer. Yonsei Med. J. 2007, 48, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Luo, C.; Shan, X.; Zhao, S.; He, J.; Cai, Z. CYP1A1 MspI polymorphism and the risk of oral squamous cell carcinoma: Evidence from a meta-analysis. Mol. Clin. Oncol. 2016, 4, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, W.; Wang, Y.; Zhuo, X.; Zhu, Y.; Wang, W.; Zhu, B.; Li, D.; Chen, Z. CYP1A1 and GSTM1 polymorphisms and oral cancer risk: Association studies via evidence-based meta-analyses. Cancer Investig. 2009, 27, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Paz-y-Miño, C.; López-Cortés, A.; Muñoz, M.J.; Castro, B.; Cabrera, A.; Sánchez, M.E. Relationship of an hRAD54 gene polymorphism (2290 C/T) in an Ecuadorian population with chronic myelogenous leukemia. Genet. Mol. Biol. 2010, 33, 646–649. [Google Scholar] [CrossRef] [PubMed]
- Paz-y-Miño, C.; Fiallo, B.F.; Morillo, S.A.; Acosta, A.; Giménez, P.; Ocampo, L.; Leone, P.E. Analysis of the polymorphism [gIVS12-6T > C] in the hMSH2 gene in lymphoma and leukemia. Leuk. Lymphoma 2003, 44, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Zaruma-Torres, F.; Lares-Asseff, I.; Reyes-Espinoza, A.; Loera-Castañeda, V.; Chairez-Hernández, I.; Sosa-Macías, M.; Galaviz-Hernández, C.; Almanza-Reyes, H. Association of ABCB1, ABCC5 and xanthine oxidase genetic polymorphisms with methotrexate adverse reactions in Mexican pediatric patients with ALL. Drug Metab. Pers. Ther. 2015, 30, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Zaruma-Torres, F.; Lares-Asseff, I.; Lima, A.; Reyes-Espinoza, A.; Loera-Castañeda, V.; Sosa-Macías, M.; Galaviz-Hernández, C.; Arias-Peláez, M.C.; Reyes-López, M.A.; Quiñones, L.A. Genetic polymorphisms associated to folate transport as predictors of increased risk for acute lymphoblastic leukemia in Mexican children. Front. Pharmacol. 2016, 5, 238. [Google Scholar] [CrossRef] [PubMed]
- Hishida, A.; Matsuo, K.; Hamajima, N.; Ito, O.; Ogura, M.; Kagami, Y.; Taji, H.; Morishima, Y.; Emi, N.; Tajima, K. Polymorphism in the hMSH2 gene (gIVS 12-67→C) and risk of non-Hodgkin lymphoma in a Japanese population. Cancer Genet. Cytogenet. 2003, 147, 71–74. [Google Scholar] [CrossRef]
- Lowsky, R.; DeCoteau, J.; Reitmair, A.; Ichinohasama, R.; Dong, W.; Xu, Y.; Mak, T.; Kadin, M.; Minden, M. Defects of the mismatch repair gene MSH2 are implicated in the development of murine and human lymphoblastic lymphomas and are associated with the aberrant expression of rhombotin-2 (Lmo-2) and Tal-1 (SLC). Blood 1997, 89, 2276–2282. [Google Scholar] [PubMed]
- Ma, L.; Ruan, L.; Liu, H.; Yang, H.; Feng, Y. ABCB1 C3435T polymorphism is associated with leukemia susceptibility: Evidence from a meta-analysis. Onco Targets Ther. 2015, 8, 1009–1015. [Google Scholar] [PubMed]
- Zhang, H.; Zhang, Z.; Li, G. ABCB1 polymorphism and susceptibility to acute lymphoblastic leukemia: A meta analysis. Int. J. Clin. Exp. Med. 2015, 8, 7585–7591. [Google Scholar] [PubMed]
- Leone, P.E.; Vega, M.E.; Jervis, P.; Pestaña, A.; Alonso, J.; Paz-y-Miño, C. Two new mutations and three novel polymorphisms in the RB1 gene in Ecuadorian patients. J. Hum. Genet. 2003, 48, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Mohd Khalid, M.; Yakob, Y.; Yasin, R.; Teik, K.; Siew, C.G.; Rahmat, J.; Ramasamy, S.; Alagaratnam, J. Spectrum of germ-line RB1 gene mutations in Malaysian patients with retinoblastoma. Mol. Vis. 2015, 21, 1185–1190. [Google Scholar] [PubMed]
- Badano, I.; Stietz, S.M.; Schurr, T.G.; Picconi, A.M.; Fekete, D.; Quintero, I.M.; Cabrera, M.D.; Campos, R.H.; Liotta, J.D. Analysis of TNFα promoter SNPs and the risk of cervical cancer in urban populations of Posadas (Misiones, Argentina). J. Clin. Virol. 2012, 53, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Brenna, S.M.F.; da Silva, I.D.C.G.; Zeferino, L.C.; Pereira, J.; Martinez, E.Z.; Syrjänen, K.J. Prevalence of codon 72 P53 polymorphism in Brazilian women with cervix cancer. Genet. Mol. Biol. 2004, 27, 496–499. [Google Scholar] [CrossRef] [Green Version]
- Pan, F.; Tian, J.; Ji, C.; He, Y.; Han, X.; Wang, Y.; Du, J.; Jiang, F.; Zhang, Y.; Pan, Y.; Hu, B. Association of TNF-α-308 and -238 polymorphisms with risk of cervical cancer: A meta-analysis. Asian Pac. J. Cancer Prev. 2012, 13, 5777–5783. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yang, X.; Chen, X.; Kan, T.; Shen, Y.; Chen, Z.; Hu, Z. Association between TNF- α polymorphisms and cervical cancer risk: A meta-analysis. Mol. Biol. Rep. 2012, 39, 2683–2688. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, X.; Chen, H.; Shang, L.; Wu, N. TP53 codon 72 polymorphism and susceptibility to cervical cancer in the Chinese population: An update meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 9055–9062. [Google Scholar] [PubMed]
- Zhour, R.; Wei, C.; Liu, J.; Luo, Y.; Tang, W. The prognostic value of p53 expression for patients with cervical cancer: A meta analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 195, 210–213. [Google Scholar]
- Hyman, D.; Taylor, B.; Baselga, J. Implementing genome-driven oncology. Cell 2017, 168, 584–599. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Perez, C.; Tamborero, D.; Schroeder, M.; Antolín, A.; Deu-Pons, J.; Perez-Llamas, C.; Mestres, J.; Gonzalez-Perez, A.; Lopez-Bigas, N. In silico prescription of anticancer drugs to cohort of 28 tumor types reveals targeting opportunities. Cancer Cell 2015, 27, 382–396. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Research Network; Weinstein, J.; Collisson, E.; Mills, G.; Mills, K.; Ozenberger, B.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar]
- The International HapMap Consortium. A haplotype map of the human genome. Nature 2005, 437, 1299–1320. [Google Scholar]
- The ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Gene | Polymorphism | Human Populations | ||
|---|---|---|---|---|
| Latin America | Caucasian | Asian | ||
| SLCO1B1 | T37041C (rs4149056) | Colombia: 0.08 (C) **; | Spain: 0.12 (C); | Han Chinese: 0.14 (C); |
| Mexico: 0.08 (C); | British: 0.14 (C); | Bangladesh: 0.05 (C); | ||
| Peru: 0.14 (C); | Finland: 0.18 (C); | Japan: 0.12 (C); | ||
| Puerto Rico: 0.12 (C) | Italy: 0.21 (C) | Vietnam: 0.10 (C) | ||
| TYMS | TSER*2/*3/*4 (rs34743033) | Self-described Hispanic: 0.41 (*2) ***; | Self-described Caucasian: 0.45 (*2); | Pacific Rim: 0.11 (*2); 0.89 (*3) |
| Self-described Hispanic: 0.59 (*3); | ||||
| Peru: 0.66 (*4); | Self-described Caucasian: 0.55 (*3) | |||
| Mexico: 0.65 (*4) | ||||
| TPMT | TPMT*2 (rs1800462) | Colombia: 0.01 (G); | Spain: 0.01 (G); | Han Chinese: 0.00 (G); |
| Mexico: 0.00 (G); | British: 0.01 (G); | Bangladesh: 0.00 (G); | ||
| Peru: 0.01 (G); | Finland: 0.00 (G); | Japan: 0.00 (G); | ||
| Puerto Rico: 0.01 (G) | Italy: 0.99 (C), 0.01 (G) | Vietnam: 0.00 (G) | ||
| TMPT*3A (rs1800460) | Colombia: 0.01 (T); | Spain: 0.04 (T); | Han Chinese: 0.00 (T); | |
| Mexico: 0.04 (T); | British: 0.03 (T); | Bangladesh: 0.02 (T); | ||
| Peru: 0.06 (T); | Finland: 0.03 (T); | Japan: 0.00 (T); | ||
| Puerto Rico: 0.05 (T) | Italy: 0.01 (T) | Vietnam: 0.00 (T) | ||
| TPMT*3C (rs1142345) | Colombia: 0.02 (C); | Spain: 0.04 (C); | Han Chinese: 0.01 (C); | |
| Mexico: 0.05 (C); | British: 0.03 (C); | Bangladesh: 0.03 (C); | ||
| Peru: 0.06 (C); | Finland: 0.03 (C); | Japan: 0.02 (C); | ||
| Puerto Rico: 0.10 (C) | Italy: 0.01 (C) | Vietnam: 0.03 (C) | ||
| TPMT*4A (rs1800584) | Argentina: 0.03 (A); Mexico: 0.04 (A) | Utah residents with Northern and Western European ancestry: 0.01 (A); Italy: 0.01 (A) | - | |
| UGT1A1 | UGT1A1*6 (rs4148323) | Colombia: 0.03 (A); | Spain: 0.00 (A); | Han Chinese: 0.23 (A); |
| Mexico: 0.02 (A); | British: 0.00 (A); | Bangladesh: 0.03 (A); | ||
| Peru: 0.00 (A); | Finland: 0.04 (A); | Japan: 0.13 (A); | ||
| Puerto Rico: 0.00 (A) | Italy: 0.00 (A) | Vietnam: 0.07 (A) | ||
| Disease | Gene | Human Populations | Reference | ||
|---|---|---|---|---|---|
| Latin American (Country) | Caucasian * | Asian * | |||
| Lung cancer | CYP1A1 | √ (Chile) | √ | √ | [78,83,84] |
| GSTM1 | √ (Chile) | √ | √ | [78,83,85] | |
| TP53 | √ (Chile) | √ | √ | [79,86,87] | |
| CYP2E1 | √ (Brazil) | Ø | Ø | [80,88] | |
| EGFR | √ (Ecuador) | √ | √ | [81,82] | |
| Breast cancer/ovarian cancer | BRCA1 | √ (Chile, Venezuela, Colombia, Mexico, Argentina, Brazil, Puerto Rico, Uruguay, Peru, The Bahamas) | √ | √ | [89,90,91,92,93,94,95,96,97,98,99,100,101,102,104,105] |
| BRCA2 | √ (Chile, Venezuela, Colombia, Mexico, Argentina, Brazil, Puerto Rico, Uruguay, Peru, The Bahamas) | √ | √ | [89,90,91,92,93,94,95,96,97,98,99,100,101,102,104,105] | |
| MTHFR | √ (Ecuador) | Ø | √ | [103,106,107] | |
| Prostate cancer | CYP1A1 | √ (Chile) | Ø | √ | [108,113] |
| GSTM1 | √ (Chile) | Ø | Ø | [108,114] | |
| MTHFR | √ (Ecuador) | √ | √ | [109,115] | |
| SRD5A2 | √ (Ecuador) | √ | Ø | [110,116,117] | |
| AR | √ (Ecuador) | √ | √ | [111,118,119] | |
| GSTA1 | √ (Brazil) | √ | √ | [96,120,121] | |
| Gastric cancer | IL-1 | √ (Venezuela) | √ | Ø | [122,127] |
| TP53 | √ (Venezuela) | √ | √ | [123,128] | |
| WNT | √ (Venezuela) | - | - | [124] | |
| 8q24 | √ (Venezuela) | √ | √ | [125,129] | |
| IL-8 | √ (Brazil) | Ø | √ | [126,130] | |
| Melanoma | CDKN2A | √ (Chile, Mexico, Argentina, Brazil, Uruguay) | √ | - | [131,132] |
| MC1R | √ (Chile, Mexico, Argentina, Brazil, Uruguay) | √ | - | [131,133] | |
| Bladder cancer | GPX1 | √ (Ecuador) | √ | √ | [134,136,137] |
| GST | √ (Argentina) | √ | √ | [135,138] | |
| NQO1 | √ (Argentina) | Ø | √ | [135,139] | |
| Colorectal cancer | CCND1 | √ (Ecuador) | √ | √ | [140,142,143] |
| TP53 | √ (Argentina) | Ø | √ | [141,144,145] | |
| Larynx cancer | GSTM1 | √ (Chile) | Ø | √ | [146,147,148] |
| Oral cancer | CYP1A1 | √ (Chile) | Ø | √ | [141,142,143] |
| GSTM1 | √ (Chile) | Ø | √ | [149,150,152] | |
| Lymphoma/leukemia | hRAD54 | √ (Ecuador) | - | - | [153] |
| hMSH2 | √ (Ecuador) | √ | Ø | [154,157,158] | |
| ABCB1/5 | √ (Mexico) | √ | √ | [155,156,159,160] | |
| COL18A1 | √ (Mexico) | - | - | [156] | |
| SLC19A1 | √ (Mexico) | - | - | [156] | |
| Retinoblastoma | RB1 | √ (Ecuador) | - | √ | [161,162] |
| Cervical cancer | TNFα | √ (Argentina) | √ | √ | [163,165,166] |
| TP53 | √ (Brazil) | √ | √ | [164,167,168] | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Cortés, A.; Guerrero, S.; Redal, M.A.; Alvarado, A.T.; Quiñones, L.A. State of Art of Cancer Pharmacogenomics in Latin American Populations. Int. J. Mol. Sci. 2017, 18, 639. https://doi.org/10.3390/ijms18060639
López-Cortés A, Guerrero S, Redal MA, Alvarado AT, Quiñones LA. State of Art of Cancer Pharmacogenomics in Latin American Populations. International Journal of Molecular Sciences. 2017; 18(6):639. https://doi.org/10.3390/ijms18060639
Chicago/Turabian StyleLópez-Cortés, Andrés, Santiago Guerrero, María Ana Redal, Angel Tito Alvarado, and Luis Abel Quiñones. 2017. "State of Art of Cancer Pharmacogenomics in Latin American Populations" International Journal of Molecular Sciences 18, no. 6: 639. https://doi.org/10.3390/ijms18060639
APA StyleLópez-Cortés, A., Guerrero, S., Redal, M. A., Alvarado, A. T., & Quiñones, L. A. (2017). State of Art of Cancer Pharmacogenomics in Latin American Populations. International Journal of Molecular Sciences, 18(6), 639. https://doi.org/10.3390/ijms18060639