2.1. Physico-Chemical Properties of PMMA-UA Composites
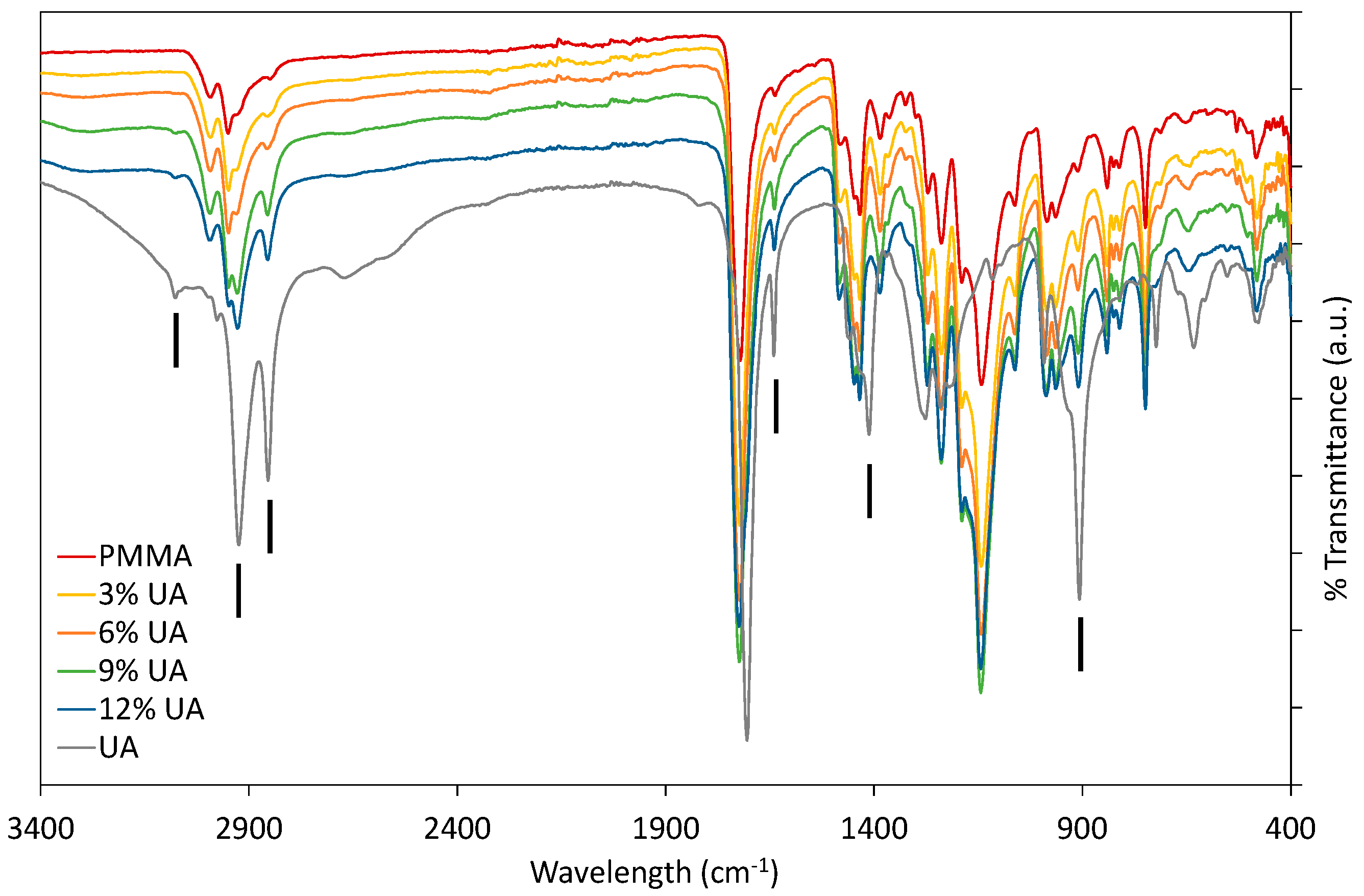
Firstly, PMMA-UA composites were studied by FTIR by comparing the spectra of native components (UA and PMMA) with composites containing different UA concentrations (
Figure 1). As expected, composites had all the characteristic peaks of PMMA. Typically the characteristic bands of the carboxylic group are in the 1570–1610 cm
−1 region for its antisymmetric vibrations, and in the 1350–1450 cm
−1 region for its symmetric vibrations [
23]. However, some peaks of PMMA and UA are similar, because of the similarity of some bonds, as for instance C=O and C–O. Therefore, the presence of UA in the PMMA-UA composites was indicated by the characteristic peaks of UA, which were not present in PMMA, such as: C=C (at 1642 cm
−1 and 908 cm
−1), =C–H (at 3078 cm
−1) and O–H (at 1411 cm
−1). Both characteristic C=C peaks of UA were evident in PMMA-UA composites, confirming the presence of UA in the composites. Nevertheless, the =C–H bend of UA was visible in the composites with 9% and 12% of UA only as a week peak shifted towards lower wavelengths (3074 cm
−1), while it was not seen in composites with 3% and 6% of UA. This indicates the interaction of the unsaturated UA tail with PMMA, probably via hydrogen bonding, because C=C peaks were clearly visible. However, the O–H peak of UA at 1411 cm
−1 was not observed in the composites, suggesting the interaction of this part of the carboxylic group with PMMA. Moreover, a broad peak with the appearance of multiple peaks in the 3300–2500 cm
−1 region (which corresponds to both O–H and various C–H vibrations) was observed in both PMMA and UA. However, the CH
3 peak is more characteristic of PMMA (see peaks at 2985 cm
−1 and 2964 cm
−1), while CH
2 is more characteristic of UA, due to a long C
11 chain (see peaks at 2927 cm
−1 and 2854 cm
−1) [
24]. All four peaks were observed in PMMA-UA composites, and as expected UA-related peaks were more pronounced in the composites with higher UA concentrations (9% and 12%).
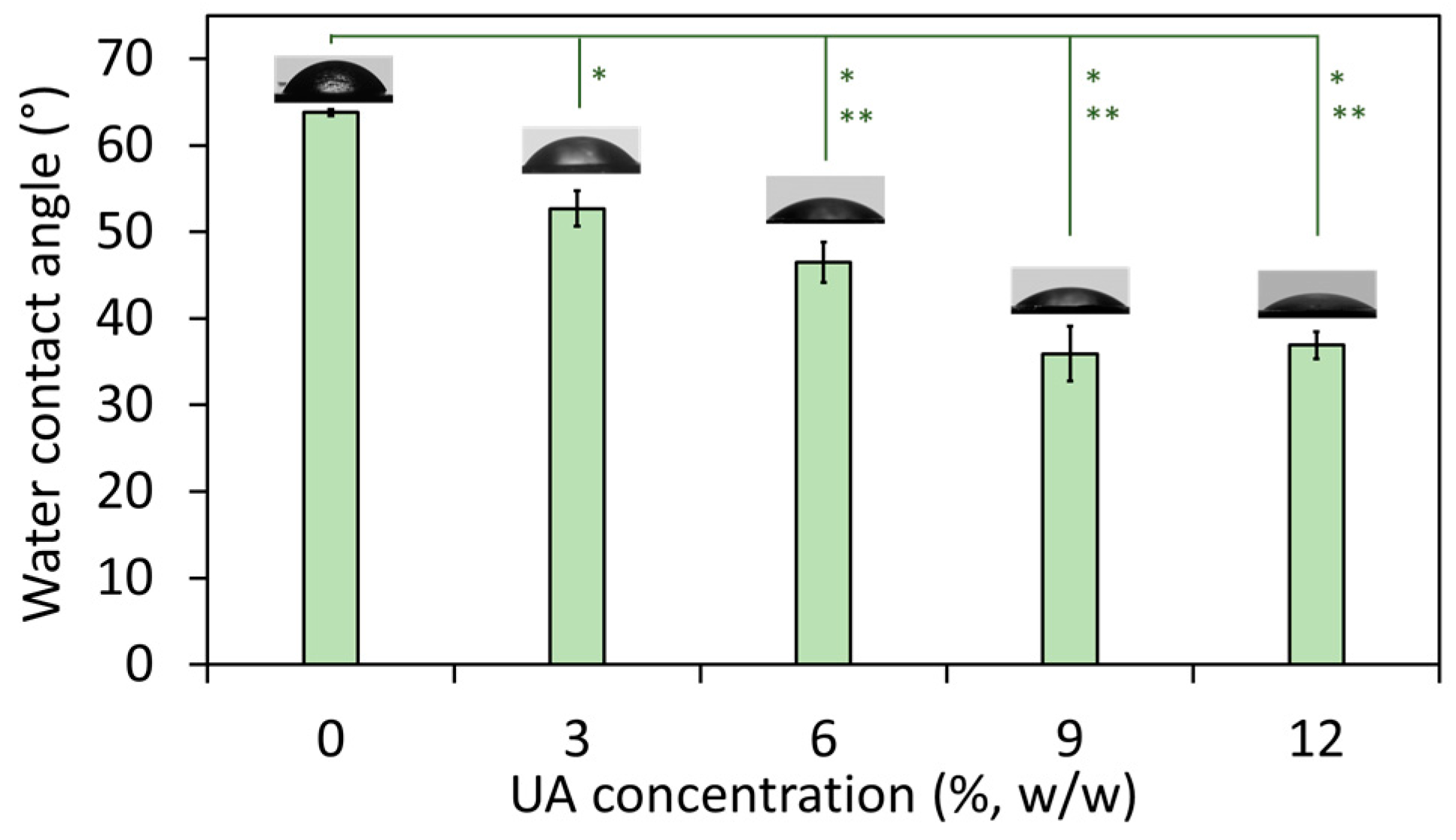
The wetting properties of the composites were studied by measuring their water contact angle. It has to be noted that all studies were done on the native (non-polished) composites’ surface and hence, there are no parallel lines and cracks characteristic for polished PMMA (therefore, polished PMMA has water contact angle ~90° [
25]). The results (
Figure 2) showed that with an increase of the UA concentration, the water contact angle decreased from 63.8° (for composite with 0% UA) to ~36° (for composites with 9% and 12% of UA). This confirms a change in the surface nature of the studied materials, which become more hydrophilic with an increase of the UA concentration in the composites. The result for PMMA is in agreement with previously reported values (such as ~68° [
26]), which has a more hydrophobic nature due to the presence of polar side-chain groups [
26]. However, in PMMA-UA composites, we expected that carboxylic groups of UA molecules would be oriented towards the side-chain groups of PMMA, while the nonpolar flexible part of the UA tail would be oriented outwards the PMMA-chain and thus outwards from the surface of the composite. In this way, an increase of the UA concentration would cause an increase of the number of nonpolar UA tails at the composites’ surface. Consequently, the top most layer of the surface of the PMMA-UA composites would appear as more hydrophilic as compared to the PMMA surface. This effect should be more pronounced with an increase of UA concentration. Such a behavior is indeed observed for composites with UA concentrations from 0% to 9%. However, the water contact angle in-between composites with 9% and 12% of UA did not change. This suggests that the presence of UA on the surface of composites was not the only reason for the observed increased hydrophilicity.
In fact, besides the chemical groups at the materials’ surface, other factors, such as the surface topology, influence the hydrophilic/hydrophobic appearance of that surface. For instance, the surface with convex structures (ideally periodic) would appear as hydrophobic (the phenomenon called lotus effect), while the smooth non-structured surface behaves as hydrophilic. Therefore, we examined the surface of the studied composites by HR SEM. In fact, it has been often reported that a polished PMMA surface is either rough or contains voids due to the lines and cracks from the polishing process [
25,
27,
28]. On the contrary, non-polished PMMA does not have lines and cracks and indeed, HR SEM micrographs of the native non-polished PMMA surface showed a smooth but not-ideally-flat surface (
Figure 3a). More precisely, the PMMA surface has a wavy topology and this is an additional reason for the more hydrophobic nature of the PMMA’s surface besides the polar side-chain groups. The surface of the composites with three percent of UA was also non-flat, but more irregular in the surface profile than PMMA (
Figure 3b). However, composites with 6%, 9% and 12% of UA had similar surfaces (see for instance
Figure 4), which were flatter than those of PMMA and the composite with 3% UA. Only a small difference could be observed in-between the composites with 9% and 12% of UA. Namely, the composite’s surface with 12% UA appeared with a slightly irregular topography and surface protrusions. Since this would increase the hydrophobicity, while an increase of the UA concentration would increase the surface hydrophilicity, it is possible that the compensation of these two opposing effects accounts for the same water wetting angle in-between composites with 9% and 12% of UA (see
Figure 2).
2.2. Anti-Candida Properties of PMMA-UA Composites
The surface of all composites was also studied by HR SEM after 24 h incubation of the composites with
C.a. Particular attention was given to the composites with zero percent and three percent of UA, because of the above observed relatively small differences in both the water contact angle (
Figure 2) and the surface topography (
Figure 3a,b). Therefore, these two composites were examined after their incubation with
C.a. A large difference could be observed between the PMMA surface almost fully covered by
C.a. (
Figure 3c) and the surface of the three percent UA composite almost without
C.a. cells (
Figure 3d). Interestingly, HR SEM micrographs with a higher magnification of the
C.a. biofilm on the native PMMA surface (
Figure 3e) show that
C.a. cells (typically as true hyphae) firstly grow in the “valley” by leaving “protrusions” unoccupied (see red arrows in
Figure 3c,e). This was also confirmed by the HR SEM micrographs with a higher magnification of the
C.a. cells on the surface of the composite with three percent UA (
Figure 3f), which shows that rare
C.a. cells (typically as yeast) were firstly found in the “valley” (see blue arrows in
Figure 3f and a micrograph of a higher magnification in
Figure 3g of the region marked in blue in
Figure 3f). In other words, it seems that
C.a. cells prefer to attach to all kinds of concave structures, such as holes and valleys. This is in agreement with previous reports of the tendency of
C.a. to attach to holes [
28]. These observations suggest that the hydrophobic nature of some materials attracts
C.a. cells (which are overall hydrophobic, hyphae more than yeast [
29]) not due to the chemical interactions, but rather because of their physical properties, such as the topology of the surface. Furthermore,
Figure 3g shows the magnified rarely observed
C.a. yeast cells on the surface of the composite with three percent of UA. These cells were not observed in the hyphal form, and more importantly the visible loss of yeast-cells’ integrity suggests that the mechanism of activity of UA is fungicidal. The deformed morphology of the surface of
C.a. cells observed by HR SEM has been reported in the presence of only one millimolar UA [
21], suggesting that the concentration of UA at the surface of the composite with three percent of UA was minimum one millimolar. This is a reasonable suggestion, because the concentration of three percent UA in PMMA-UA solid was 190 mM, and thus one millimolar UA at its surface would represent a release of only 0.5% UA. Similarly, for some other unsaturated fatty acids (polyunsaturated with C18 to C22 in contrast to UA, which is monounsaturated with C11) has been reported to cause nuclear condensation and fragmentation and thus apoptosis of
C.a. yeast cells [
30].
Moreover, rare
C.a. cells could be seen on the surface of the composites with 6%, 9% and 12% of UA (
Figure 4a–c). This is consistent with the above discussed relatively similar both water contact angles and surface topology. In all, clearly less
C.a. cells as yeast were observed on the surface of composites with any UA concentration as compared to the native PMMA surface. This observation suggests that the surface of the composites prevent the attachment of
C.a. cells, i.e., that the PMMA-UA surfaces are
C.a. repellent. These results showed that our material both: (a) prevent the attachment of
C.a. cells and (b) have fungicidal effect, because the rare sessile
C.a. cells were often dead.
An additional method to study the antifungal potential of materials is the Kirby–Bauer test (sometimes referred to as a disk diffusion method), where a disk of the studied material is placed on the surface of an agar plate seeded with microorganisms, here
C.a. cells. After 48 h of incubation at 37 °C, the inhibition zone comprising the disk and the clear region (where
C.a. growth was inhibited) was measured for the different composites after their preparation (T
0) and six days after their preparation (T
6). The results (
Figure 5 and
Supplementary Figure S1) showed an increase of the inhibition zone along with increasing UA concentration, especially for composites with ≥6% of UA. Moreover, the results were similar in-between the studied time points. These results show that our composites prevent the growth of
C.a. cells in the vicinity of the composites. Moreover, these results are in agreement with the above given results and confirm the anti-
Candida activity of the developed composites against the sessile
C.a. cells. However, the HR SEM study and the Kirby–Bauer test should be complemented by an additional quantitative test.
Therefore, to quantify the number of
C.a. cells, which were attached to the surface of the studied materials, we performed an XTT test, which measures the number of metabolically active
C.a. cells. The XTT test was performed after the composites’ preparation (after 0 days, marked as T
0) and six days after that (marked as T
6). The results given in
Figure 6a show the relative number of metabolically active
C.a. cells on the surface of the studied PMMA-UA composites (as compared to the surface of native PMMA). We found that the percentage of metabolically active
C.a. cells was less than five percent on composites with 9% and 12% of UA as compared to native PMMA for both studied timepoints, i.e., T
0 and T
6. In other words, the number of metabolically active
C.a. cells was reduced by more than 95% in composites with 9% and 12% of UA as compared to native PMMA for both timepoints. On the surface of the composites with six percent of UA, 3.7% and 29.6% of metabolically active
C.a. cells were measured for T
0 and T
6, respectively; while on composites with three percent of UA, the corresponding values were 59.4% and 68.6%, respectively (even these values were significantly different as compared to native PMMA,
p = 0.001). Overall, the obtained XTT results showed a considerable reduction of the number of metabolically active
C.a. cells on the surface of PMMA-UA composites in both T
0 and T
6. This reduction was more than 95% on the composites: with ≥6% UA in T
0 and with ≥9% UA in T
6. Therefore, the developed PMMA-UA composites showed an antifungal surface activity towards
C.a. sessile cells at an early stage of colonisation (after 24 h), which has not been previously observed in composites of acrylic-denture liners with UA, which was explained by probably low concentrations of the released UA [
31]. The low percentages of metabolically active
C.a. cells measured on the composites’ surfaces are in agreement with the above given HR SEM observation of the PMMA-UA surfaces, where very few
C.a. cells were observed, often deformed and with loss of their cell integrity (see
Figure 3g), i.e., probably metabolically non-active.
Beyond the anti-
Candida activity of the PMMA-UA surface found against the sessile
C.a. cells, it was of interest to study if UA molecules from the composites could also affect planktonic
C.a. cells. This was a challenge, because UA has a limited solubility in water (depending on the temperature and pH, for instance 38.46 mg/L at 20 °C and pH 4.27) and this is reduced in the presence of cell media and
C.a. cells. In order to study this, we performed an XTT test on the supernatant, which was above the surface of the composites during 24 h incubation with
C.a. cells. The obtained results (
Figure 6b) showed similar values for the planktonic as for the sessile
C.a. cells. This could be a consequence of the opposing effects of: (a) an expected lower drag-to-cell ratio (drug is here UA) in medium than on the surface of the corresponding composites, and (b) the previously reported lower drag resistance for the planktonic as compared to the sessile
C.a. cells at the comparable cell concentration [
32,
33]. Surprisingly, the results given in
Figure 6b showed similar results as on the surface of the composites for both studied timepoints. This suggests that our composites affected not only sessile, but also planktonic
C.a. cells, by reducing the number of metabolically active both sessile and planktonic
C.a. cells. Therefore, we plotted in
Figure 6c the percentages of the total metabolically active
C.a. cells, which include both the sessile and the planktonic
C.a. cells, as a function of the UA concentration. We found that the percentage of the total metabolically active
C.a. cells was less than five percent for composites with 9% and 12% of UA for both timepoints (i.e., percentage of the total metabolically active
C.a. cells was reduced more than 95%). On the other hand, for composites with 3% and 6% of UA, the percentage of the total metabolically active
C.a. cells was 68.6% and 5.2%, respectively, after T
0 (while 87.2% and 18.4%, respectively, after T
6). This also confirms the above given fungicidal activity of the here developed PMMA-UA composites.
The effect of UA on the growth of
C.a. planktonic cells was also studied on agar containing different concentrations of UA (from 0.0125% to 0.4%, see the Materials and Methods for details). Firstly, after the incubation (for 24 h at 37 °C) of
C.a. cells at a concentration of 10
6 C.a. cells/mL, we measured the absorbance of the supernatant containing the planktonic
C.a. cells at 620 nm, A
620. The results expressed as average percentages of A
620 readings compared to control (0% UA) are given versus UA concentration in agar in
Figure 7a. Interestingly, even the lowest UA concentration (0.0125%,
w/
w) in agar was sufficient to reduce more than 90% of the
C.a. planktonic cell growth, i.e., 0.0125% was already the concentration above the one necessary to reduce
C.a. growth of 90% with respect to the control (called minimal inhibitory concentration (MIC) for 90% growth reduction, i.e., MIC90). Similar results were obtained with the XTT test done on the same suspensions (
Supplementary Figure S2). Since we could not measure the dose of UA to which
C.a. cells are exposed, we calculated the maximal theoretical amount of UA to which a
C.a. cell could be exposed to in such an experiment (if all UA from the agar would come into the suspension of the incubated
C.a. cells) by dividing the total mass of UA in the agar substrate by the number of incubated
C.a. cells. The so-obtained results (
Figure 7b) showed that the maximal theoretical concentration could be from 0.25 to 8 ng
UA/
C.a. cell for 0.0125% to 0.4% UA in agar, respectively. Since these values seemed to be rather high (the lowest one is above MIC90), we performed the same experiment with ten times higher concentration of
C.a. cells (10
7 C.a. cells/mL). The results (
Figure 7c,d) showed that MIC90 for 10
7 C.a. cells/mL was below the lowest studied concentrations of 0.0125% and 0.025% of UA in agar (corresponding to values between 0.025 and 0.050 ng
UA/
C.a. cell). However, even the lowest UA concentration in agar of 0.0125% (i.e., 0.025 ng
UA/
C.a. cell) was higher than MIC for 50% growth reduction (MIC50). Thus, only a limit to what MIC90 could be was observed. However, the value for a limit below which is MIC90 found here (0.025%,
w/
w, corresponding to 0.025 μg
UA/mL) is much lower than the 256 μg
UA/mL, which was previously reported for planktonic
C.a. cells incubated at 10
7 C.a. cells/mL [
31]. We have to note that percentages of the
C.a. growth in-between samples with the similar drag-to-cell ratios (e.g., 3.1%
C.a. growth for 0.25 ng
UA/
C.a. cell at 10
6 C.a. cells/mL, and 9.9%
C.a. growth for 0.4 ng
UA/
C.a. cell at 10
7 C.a. cells/mL) was typically higher at higher
C.a. cells’ concentration (10
7 C.a. cells/mL). This is in agreement with the previous reports that the resistance of
C.a. cells to drags increases with a higher cells’ concentration [
32,
33]. These results also confirm the fungicidal effect of UA on planktonic
C.a. cells.
Besides the confirmation of the anti-
Candida effect of UA on both sessile and planktonic
C.a. cells, the only study technique that gave us information on the
C.a. cell type was HR SEM (see
Figure 3). Therefore, we performed an embedded filamentation assay for both 24 and 48 h of incubation to have an additional information on the
C.a. cells development in the presence of UA as compared to the control without UA (representative photomicrographs are given in
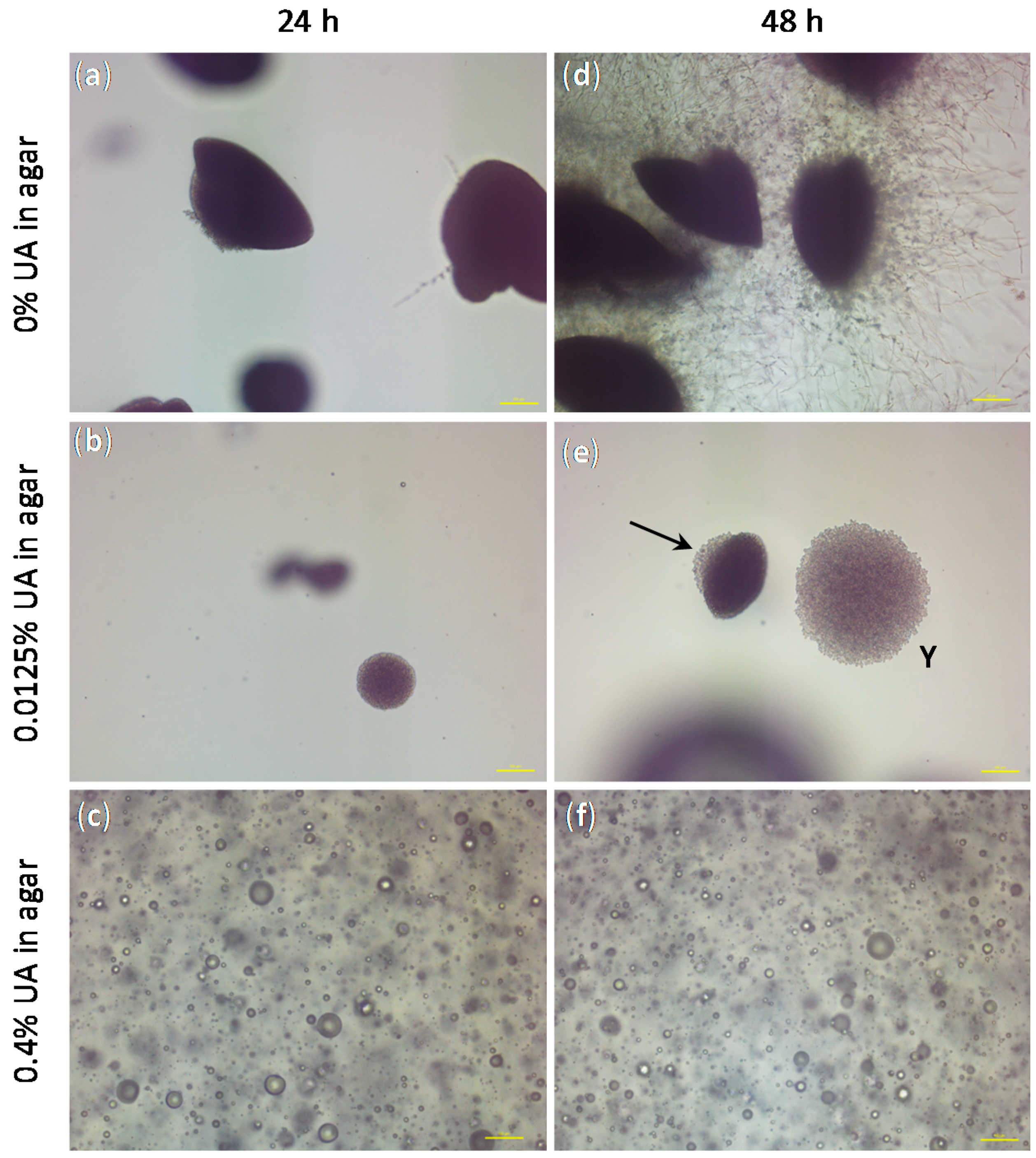
Figure 8). The concentrations of UA in agar were chosen to be the lowest and the highest values used in the previous test (i.e., 0.0125% and 0.4%). After 24 h of incubation,
C.a. cells embedded in agar without UA formed spindle-shaped colonies consisting mostly of yeast cells with rare peripheral hyphae and/or pseudohyphae, which typically had lateral yeasts (
Figure 8a), as often reported [
34,
35]. At the same time point,
C.a. cells embedded in agar with 0.0125% UA formed yeast colonies (smaller than in the control and without hyphae,
Figure 8b), while in agar containing 0.4% UA,
C.a. cells could not be observed, but only spheres probably originating from the leakage of the intracellular material after
C.a. cells’ death (
Figure 8c). After 48 h of incubation,
C.a. cells embedded in agar without UA formed numerous radially emerging peripheral hyphae, pseudohyphae and lateral yeasts from spindle-shaped yeast colonies (
Figure 8a). Moreover, all embedded colonies formed hyphae (since the incubation was at 37 °C) [
35]. On the contrary, in agar with 0.0125% of UA, spindle-shaped colonies with yeast outgrowth and yeast colonies (indicated by the arrow and Y in
Figure 8e, respectively) could be observed, both without filamentation; while the
C.a. cells in agar with 0.4% UA observed after 48 h (
Figure 8f) looked the same as after 24 h. These results showed fungicidal effect of UA at 0.4% in agar, while a UA concentration of 0.0125% was sufficient to inhibit the yeast-to-hyphae transition and to suppress the
C.a. growth.
The above given study demonstrates that the developed PMMA-UA composites and UA itself are efficient anti-
Candida agents. However, the use of such composites in patients requires a low toxicity of the antifungal material to human cells. Especially the strong fungicidal effect of UA observed in
C.a. cells poses the question whether PMMA-UA composites at the here-used UA concentrations (up to 12%) would be toxic to human cells. Therefore, in order to assess the effect of PMMA-UA composites as a final material, we performed a preliminary cytotoxicity study by using the 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxypenyl)-2-(4-sulfophenyl)-2
H-tetrazolium (MTS) test according to the previously reported method [
36]. The test (which comprise of 24 h incubation with cells) was performed for two timepoints being six days apart (T
0 and T
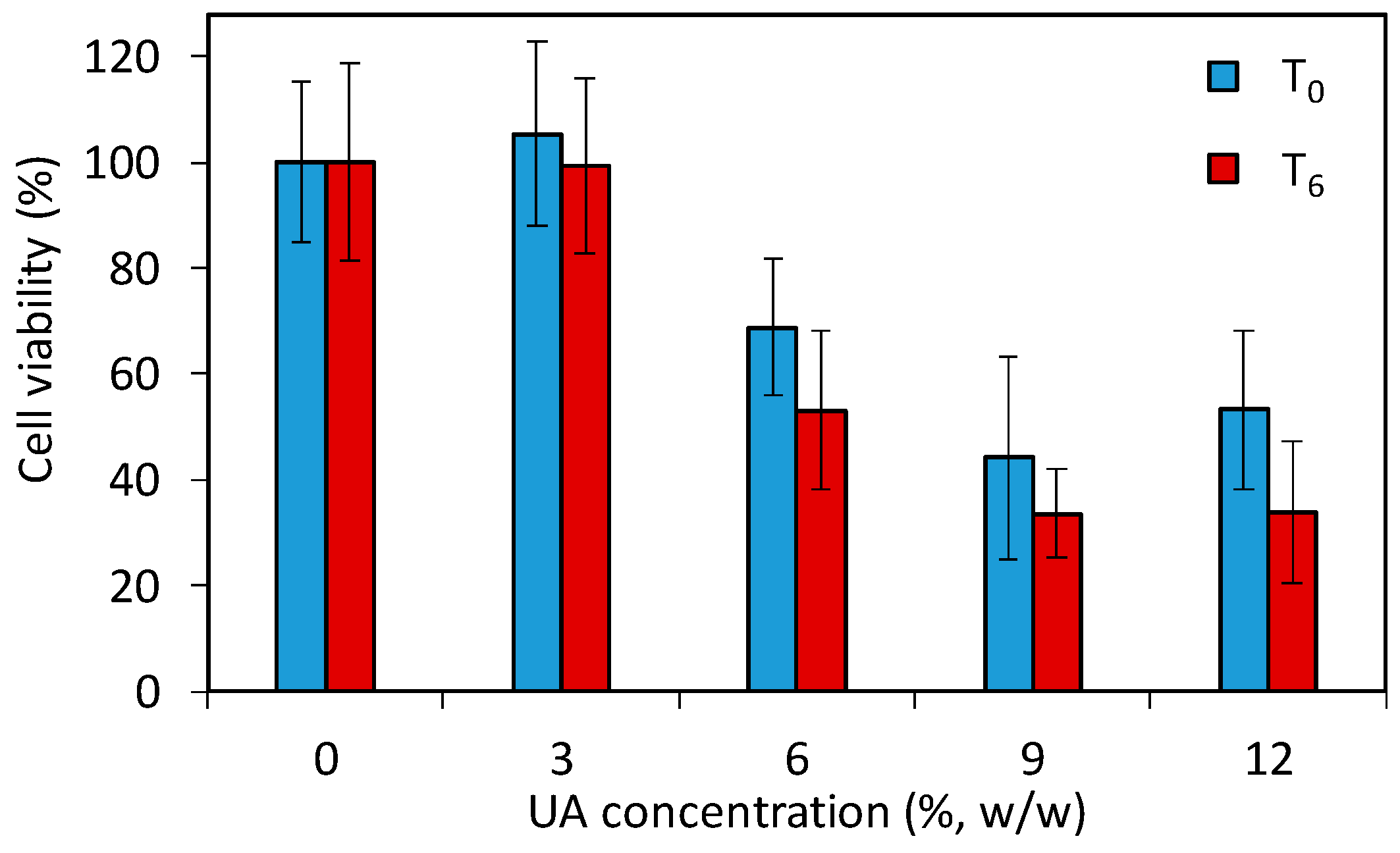
6) with respect to composites preparation. The results (
Figure 9) showed a decrease of the cell viability for composites with six percent UA (68.8% and 53.1% at T
0 and T
6, respectively), which was even more pronounced for the composite with nine percent UA (44.2% and 33.6% at T
0 and T
6, respectively). Moreover, even at T
0, meaning zero days after composite preparation and after 24 h immersion of composite-discs in the medium, there was release of UA from composites with greater than six percent, which was sufficient to affect cells. Therefore, composites with 9% and 12% of UA cannot be used as antifungal materials in or on patients due to their toxic effects on human cells (i.e., less than 50% of cells remained viable). However, based on the here-obtained values for cells viability in the preliminary toxicity test, PMMA-UA composites with UA concentrations of up to six percent look still promising for such applications.
So far, there have been reported only a very few studies of one commercial acrylic denture liner (i.e., Coe Soft, GC America, Alsip, IL, USA), which according to the authors contain UA (70 mM UA [
29], or one to five percent UA [
37], or non-specified UA concentration [
31]), but according to the producer this product contains zinc undecylenate [
38]. Moreover, it is difficult to compare our results with their results due to the numerous differences in the study conditions (
C.a. strain, inoculum concentration, temperature and time of incubation, planktonic or sessile cells, etc.). McLain et al. reported a sevenfold reduction of the
C.a. germ-tube formation without an effect on the growth rate in the presence of 10 μM UA (as compared to control) (
C.a. strain ATCC 28367 at concentration 10
6 C.a. cells/mL incubated at 39 °C for two hours) [
29]. In the same study, the authors reported that on the surface of Coe Soft all adhered
C.a. cells were yeast form [
29]. Although the ability of
C.a. cells to change their form was suppressed, the adherence of
C.a. cells and their growth were not inhibited. However, one other study reported the reduction of a
C.a. biofilm formation, but a higher number of hyphae on Coe Soft as compared to the PMMA surface (
C.a. strain ATCC 90028 at concentration 10
7 C.a. cells/mL incubated at 37 °C for 24 h) [
37]. A more recent study reported that UA released from Coe Soft was appropriate to significantly affect mature
C.a. biofilms (48 and 72 h) by reducing both the cell counts and their metabolic activity, but not sufficient to influence
C.a. at an early stage of the colonization, i.e., 24 h (
C.a. strain ATCC 90028 at concentration 10
7 C.a. cells/mL incubated at 37 °C for 24–72 h) [
31]. The same study found a minimal inhibitory concentration required to inhibit
C.a. planktonic cells growth at concentration of released UA of 256 μg/mL; but at the same concentration of released UA, the fungistatic effect on
C.a. growth was shown only until eight hours [
31]. None of these reports studied the potential toxic effects of UA on human cells. However, beyond this one studied commercial acrylate containing either zinc undecylenate or UA, no new antifungal materials have been developed by incorporation of UA into some commercial acrylate for medical application (especially for dentistry).
Here, we have developed new anti-Candida materials by incorporation of UA into commercial PMMA, which allowed UA to be released in a sufficient amount to show simultaneously the inhibition of the adherence at early stage of the colonization (24 h), morphogenesis (yeast-to-hyphae transition) and growth of both sessile and planktonic C.a. cells. Therefore, we here present several pieces of evidence that novel PMMA-UA composites have strong fungistatic and fungicidal anti-Candida effect. We have also shown that beyond fungicidal effect, UA at larger concentrations (as in composites with ≥9% of UA) can be cytotoxic for human cells, and thus, care has to be taken when UA is used in or on patients.
By taking into consideration global health issues with fungal infections [
1,
2,
3], this study shows potential in combating fungal infections, especially by
C.a., thanks to the development of novel fungicidal and/or fungistatic materials in such a way that a sufficient amount of the active antifungal component can reach both sessile and planktonic fungal cells.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








