Abstract
Abiotic stress affects the growth and development of crops tremendously, worldwide. To avoid adverse environmental effects, plants have evolved various efficient mechanisms to respond and adapt to harsh environmental factors. Stress conditions are associated with coordinated changes in gene expressions at a transcriptional level. Dehydrins have been extensively studied as protectors in plant cells, owing to their vital roles in sustaining the integrity of membranes and lactate dehydrogenase (LDH). Dehydrins are highly hydrophilic and thermostable intrinsically disordered proteins (IDPs), with at least one Lys-rich K-segment. Many dehydrins are induced by multiple stress factors, such as drought, salt, extreme temperatures, etc. This article reviews the role of dehydrins under abiotic stress, regulatory networks of dehydrin genes, and the physiological functions of dehydrins. Advances in our understanding of dehydrin structures, gene regulation and their close relationships with abiotic stresses demonstrates their remarkable ability to enhance stress tolerance in plants.
1. Introduction
Environmental stress such as drought, high salinity, and extreme temperatures can negatively affect plant growth and development. To confront various environmental constraints, plants have evolved systematic defense mechanisms to decrease the loss of intracellular water in order to maintain the integrity of cell structures.
Late embryogenesis abundant (LEA) proteins, initially discovered in cotton [1], have been found to accumulate in many plant species during environmental stress and seed development stage. Subsequent studies of LEA proteins indicate their involvement in improving stress resistance in plants. LEA proteins are highly hydrophilic, and their structural analysis implies that they are intrinsically disordered proteins (IDPs), which provide a flexible property to interact with metal ions and biomolecules [2,3]. There are 7 groups of LEA proteins based on sequence similarity and structural property [4]. Although variations have been observed between different LEA groups, they share qualities of high hydrophilicity and Gly richness [5,6,7].
The most commonly described LEA proteins are known as dehydrins (or group II LEA). Like typical LEA proteins, dehydrins are highly hydrophilic and thermostable. Dehydrins mainly accumulate in the later stages of embryogenesis in response to various environmental stresses [4]. According to recent studies, dehydrins have been found widely distributed in angiosperm [5,6,7,8,9,10], gymnosperm [11,12,13] and bryophyte [14,15] plants.
The expression of dehydrins can be induced by various abiotic factors and phyto-hormones [6,7,16,17,18]. Since some dehydrins can respond to abscisic acid (ABA), they are referred to as Response to ABA (RAB) proteins. For instance, the expression of a wheat dehydrin WZY2 can be stimulated by drought, salinity and ABA [9,17]. The main functions of dehydrins have been discovered to participate significantly in stabilizing membranes, enzymes, and nucleotides in cells under abiotic stresses. Studies on transgenic plants have also proved that overexpression of dehydrins could enhance the tolerance to various abiotic stresses. Two dehydrins from the moss Physcomitrella patens, PpDHNA and PpDHNC, have been shown to improve the stress resistance to salinity and drought in transgenic Arabidopsis. The transgenic lines also exhibited strong scavenging ability for reactive oxygen species (ROS) with enhanced content of superoxide dismutase (SOD) and catalase (CAT) [14]. On the other hand, knockout of PpDHNA in Physcomitrella patens mutant lines severely impaired their ability to recover from salt and osmotic-stress treatments [19]. Similar to PpDHNA, overexpression and RNAi assays in transgenic Arabidopsis imply that cold-regulated (COR) proteins, dehydrins (COR15A and COR15B) are necessary for full cold acclimatization as well [20].
Several studies on differential expression of dehydrins in frost-tolerant and frost-susceptible wheat genotypes have revealed that transcripts of many dehydrins are higher in frost-tolerant wheat, such as WCS120, Wdhn13 [21,22]. Similar to WCS120, a dehydrin from barley (DHN5) was also accumulated in higher amounts in winter cultivars than spring ones [23,24]. These results suggest a significant correlation between dehydrins and acquired frost tolerance (FT) of plants, hence these dehydrins can also be considered promising FT markers. We also draw a phylogenetic analysis tree of well-studied dehydrins in this article (Figure 1).

Figure 1.
Phylogenetic analysis of well-studied dehydrins. Phylogenetic relationship of well-studied dehydrins. The bootstrapped tree with 1000 replicates was constructed using MEGA 7 tools.
Although the detailed and overall mechanisms in dehydrin functioning are still ambiguous, some protective models have been proposed to elucidate their role in different aspects, and will be discussed in this review. In addition, advances in dehydrin structures, gene regulations and their close relationships with abiotic stresses will be elucidated in accordance with recent studies.
2. Molecular Structure of Dehydrins
Most dehydrins are low molecular weight proteins, and their molecular weights range from 9 kD to 200 kD [25]. Dehydrin protein families contain three conserved motifs, named K-, Y- and S-segments. Among the conserved motifs, a Lys-rich residue called K-segment is prevalent in all dehydrins. In angiosperms, the common sequence of K-segment is EKKGIMDKIKEKLPG, while in gymnosperms, dehydrins share a more variable sequence: (Q/E)K(P/A)G(M/L)LDKIK(A/Q)(K/M)(I/L)PG [26,27]. K-segment is usually located near the C-terminal and participates in forming a class of A2 amphipathic α-helix [28]. Some dehydrins possess more than one K-segment, and they share distinct amino acid sequences. Although the amino acid position in K-segment varies slightly, conserved residues still exist. The conserved residues are Lys-Ile-Lys-Glu in the core, Glu-Lys-Lys in the N-terminal and Lys-Leu-Pro-Gly in the C-terminal of this segment [25]. Y-segment (DEYGNP) is an N-terminal conserved sequence, and it is homologous to the nucleotide-binding of chaperones of bacteria and plants [29]. S-segment is a serial of the serine cascade sequence (SSSSSSSD), and this segment often has one copy in the dehydrins. Based on their conserved segments, dehydrins are usually classified into five subfamilies: KnS, YnSKn, Kn, YnKn, and SKn [30].
In addition, some untypical segments also appear in many dehydrins, and these segments exhibit various properties and functions. One segment that has been found is a polyhistidine- or histidine-rich motif, which contributes to binding ability of dehydrin with ligands. OpsDHN1 from Opuntia streptacantha with a polyhistidine motif exhibited self-dimerizing via a split-ubiquitin yeast two-hybrid system; subsequent deletion of this motif results in loss of self-interaction [31]. Three dehydrins from Arabidopsis, AtCOR47, AtERD10, and AtRAB18, also showed formation of homodimeric complexes, as well as heterodimeric complexes with each other [32].
In addition to dehydrin-dehydrin interaction, a histidine-rich motif also participates in dehydrin metal binding and lipid binding. A KS-type dehydrin, AtHIRD11, inhibited generation of reactive oxygen species (ROS) in a Cu-ascorbate system by binding with free Cu2+. Meanwhile, dehydrins with His-flanking K-segments are capable of binding to membrane structures [33]. In many YnSKn-type dehydrins, the K-segment and S-segment are linked by a fixed motif, GXGGRRKK (X can be any amino acid), which suggests a potential functional linkage between K- and S-segments. The GXGG motif is highly flexible and participates in the interaction of negatively charged phosphoserines with K-segment, while RRKK motif is considered to be a nuclear localization signal [34,35,36].
Based on circular dichroism (CD), nuclear magnetic resonance (NMR) and Fourier Transform Infrared Spectroscopy (FTIR) analysis, dehydrins are characterized as intrinsically disordered proteins (IDPs) [37,38]. These do not adopt a fixed structure, but contain flexible mobile polypeptides [39,40]. The NMR spectrum provides a general view of IDP structures with respect to the chemical shift values in their secondary structures [41]. The mutual authentication from CD, FTIR, and NMR studies has helped to clearly uncover the transformation of dehydrin secondary structures.
Interaction of dehydrins and their ligands (metals, biomembranes, and proteins/enzymes) could strongly induce the formation of the ordered protein structure of dehydrins. According to Hara et al., (2013), a KS-type dehydrin AtHIRD11 showed an obvious signal minimum near 200 nm under CD determination, which represents a random coil. However, disordered conformation was attenuated by Cu2+ [42]. A similar conformational change was also revealed in TsDHN-1 and TsDHN-2 with Zn2+ via CD and FTIR assay. In addition, this phenomenon is also associated with variation in temperature [43]. Lactate dehydrogenase (LDH) is widely used in dehydrin cryoprotective assays [7,9,16,44,45]. It has been reported that K2 and YSK2 dehydrin were able to protect LDH activity better than BSA. Compared to K2, YSK2-type dehydrin provided a 50% higher LDH activity at a 2-fold lower protein concentration, suggesting that larger proteins could provide more efficient protection [45].
In another study, a small model dehydrin Vitis Riparia K2 was selected to examine protection of membranes under freeze-thaw stress with CD and NMR spectrum. The results showed the ability of this protein to bind with liposomes which contain phosphatidic acid, leading to protection of liposomes under freeze-thaw stress. Further analysis implies that K2 dehydrin did not significantly affect liposome surface accessibility or lipid mobility; however, presence of K2 could decrease membrane transition temperature by up to 3 °C. Experiments involving NMR showed that conserved lysine-rich segments participated in the binding of dehydrin to membranes and poorly conserved φ-segments, and did not show any relevance in dehydrin binding and protection [46].
The results are consistent with previous studies that revealed structure of dehydrins could become ordered or partially ordered through binding with ligands [15,44,47]. The lack of defined secondary structures may contribute to flexible binding properties of dehydrin with other ligands other than Cu2+ and Zn2+, such as lipid membrane and dehydrins [31,33].
3. The Regulation of Dehydrin Genes under Abiotic Stresses
The expression of dehydrins is strongly induced by adverse environmental factors. Some dehydrins share intricate response mechanisms, and their expressions can be triggered by multiple stresses. Abiotic stresses can trigger rapid accumulation of dehydrins in plants through multiple signaling pathways (Figure 2).
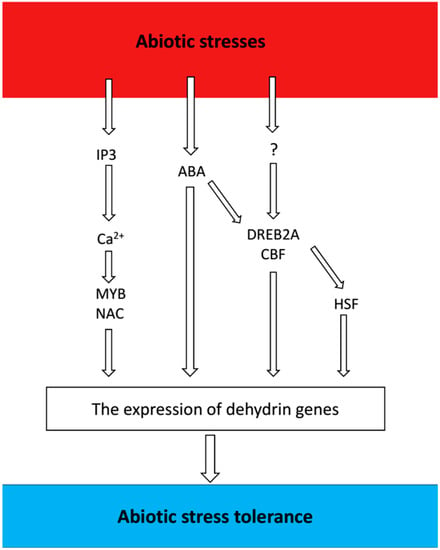
Figure 2.
Overview of signaling pathways and factors involved in signaling regulation of dehydrin genes. The figure shows the signaling components involved in the abiotic stress responses that are described in the text. Question marks represent as-yet-unidentified factors in the corresponding signal transduction pathways.
3.1. Drought Stress and Dehydrins
Drought is one of the most prevalent constraints in plant development due to severe restriction of cell extension, reduction of cell water content, and effects on photosynthetic activity. In addition, drought stress can induce secondary stresses, such as osmotic and oxidative stress [48]. ShDHN is a gene whose expression reaches its maximum level (almost 12-fold-changes) in 6 h under drought conditions. Similarly, SiDhn2 expression increases 12-fold after 3 h of dehydration stress [49,50]. However, dehydrin WZY2 gene shows lower response to dehydration for its maximum expression level at 24 h after stress [17]. These findings suggest that different dehydrins may respond at different time intervals in order to mediate distinct stages of drought stress.
Abscisic Acid (ABA) acts as a vital mediator in plant abiotic stress, regulates expression of drought stress-related genes, and protects cells against damage due to abiotic stress leading to increased stress tolerance. Presence of ABA-responsive cis-elements (ABRE) in promoters of dehydrin genes enable them to participate in the ABA-dependent pathway. So far, several ABRE motifs are found in dehydrin gene promoters in wheat (CCACGTGTC/GACACGTGG) [51], maize (CCGACGCG/CGCGTCGG) [52], rice (CCGACGCG/CGCGTCGG) [53] and barley (CCAACGCG/CGCGTTGG) [54]. Many dehydrin genes are induced by exogenous ABA treatment, as reported in various studies [17,49]. In ABA-dependent signaling, ABRE-binding protein/ABRE-binding factors (AREB/ABFs) play a pivotal role in regulating the expression of ABA-dependent genes. Moreover, the AREB/ABF family belongs to group A of the basic leucine zipper (bZIP) transcription factor (TF) family, and studies have revealed that overexpression of AREB in the transgenic plant could significantly activate the expression of dehydrins [55].
Some dehydrins possess dehydration-responsive elements (DRE, A/GCCGAC motifs) with or without ABRE motifs. The DRE motif is one of the core domains for dehydrin genes in responding to drought stress in the ABA-independent pathway. Promoters of peach dehydrins PpDhn1 and PpDhn3 share a typical CRT element (known as C-repeats, RCCGAC) upstream of the TATA box [56]. Additionally, some SKn- and YnSKn-type dehydrins in plants also possess de novo DRE/CRT motifs [57]. Previous studies have provided evidence that transcription factors DREB1 and DREB2 are capable of binding to the DRE element of Arabidopsis rd29A in order to mitigate drought stress [58,59]. Gupta et al. found that in transgenic tobacco, a Salicornia brachiate DREB factor could significantly up-regulate the expressions of NtERD10B and NtERD10D [60]. Furthermore, overexpression of Vigna radiata DREB gene VrDREB2A in transgenic Arabidopsis also enhances the expression of AtCOR15A, AtCOR15B, AtKIN1, AtRD17, AtRD29A, and AtRD29B, indicating an important role of the DREB family in the dehydrin regulation pathway under drought stress [61]. Kim et al. discovered binding activities of three AREB members (AREB1, AREB2, and ABF) to ABRE cis-elements in DREB2A promoter, which is also necessary for DREB2A expression under osmotic stress. Furthermore, in plant vegetative tissues, AREB was also shown to interact with DREB in ABA-related gene expression under dehydration conditions [62,63]. VvDHN1 from V. vinifera leaves is hypersensitive to both drought and ABA; after four days of drought stress and eight hours of ABA treatment, the expression levels of the VvDHN1 gene reach about a 350-fold change and a 170-fold change, respectively [18]. These results imply that dehydrins are participate broadly in complicated crosstalk signaling networks under the regulation of drought-related TFs.
3.2. Cold Stress and Dehydrins
Cold stress can directly influence the fluidity of plant cell membrane and enzyme activity, resulting in metabolic disturbance, photosynthesis inhibition, material transportation disorder and, finally, damage to plants [64]. Cold stress can induce ShDHN and WZY2 dehydrin, with more than 40-fold and 80-fold changes in gene expression levels, respectively. Moreover, promoters of these dehydrin genes contain low temperature-responsive elements (LTREs), which play a crucial role in their response to cold stress. For example, ShDHN promoter contains two LTRE elements [49]. However, the regulation of LTREs still needs to be clarified. In addition to drought stress response, DREs also participate in cold stress signaling. In Arabidopsis, overexpression of AtDREB1A or AtDREB2A induces cold stress-related genes such as rd29A and COR47 under cold stress [63].
C-repeat response element binding factors (CBFs), belonging to the DREB family, contain AP2 DNA binding domains. AtCBF1, AtCBF2, and AtCBF3 can be up-regulated more rapidly and significantly by cold stress than ABA and dehydration [65,66]. Previous studies have shown that some CBFs are specifically induced by cold stress. Dehydrin gene promoters from numerous species contain the low-temperature response element CRT. Study on CBFs has revealed its functions in binding to DRE or CRT motifs [67]. In addition, several DREB promoters have been characterized that contain ABRE motifs, including MACGYGB and CGTGG, in Sorghum, and over-representation of motifs related to other phytohormone signalings in DREB promoters has also been reported [68]. Based on microarray analyses, several dehydrins induced by cold and dehydration have been characterized in Arabidopsis, rice, and soybean [69].
Interestingly, some evidence showed that dehydrin gene promoters containing DREs did not respond to drought and/or cold stress. PpdhnA promoter contains one DRE and several ABRE motifs, but qRT-PCR results showed that this gene could not be induced by cold stress [15]. This reveals a complicated mechanism of cold/drought regulation in dehydrin genes which needs further elucidation to understand the related networks involved.
3.3. Heat Stress and Dehydrins
Heat stress is an intricate process that often directly causes protein denaturation and aggregation, and increases membrane lipids fluidity [70]. Evidence from many studies has revealed a strong association between dehydrins and plant tolerance to heat stress. For instance, dehydrin CsLEA11 and WZY2 proteins were able to protect both recombinant E. coli and activity of the LDH enzyme under heat stress [9,16]. Dehydrin DHN-5 from wheat was found to play a relevant role in protecting enzyme β-glucosidase (bglG) against heat stress. Truncation assay of DHN-5 indicated that K-segments were vital to the thermal protection of dehydrins [71,72].
In wheat, promoters of dehydrin TaDHN1 and TaDHN3 genes possess heat stress elements (HSEs), that are involved in heat stress response [73]. HSEs can be specifically recognized by heat stress transcription factors (HSFs) due to its conserved N-terminal DNA binding domain (DBD). However, there is little evidence showing that HSEs is also present in other known promoters of dehydrins.
Other than HSEs, promoters of dehydrins often possess DREs with intricate regulation mechanisms under heat stress. In Arabidopsis, the DRE-binding protein 2A (DREB2A) could directly regulate expression of AtHSFA3 through binding two DRE elements in the AtHSFA3 promoter [74]. As mentioned above, the DREB protein family also participates in the regulation of dehydrins under drought and cold stresses, implying that they may share similar strategies with plant tolerance to drought, cold and heat stresses.
3.4. Salt Stress and Dehydrins
The plant tolerance mechanism to salt stress has been widely studied and extensively characterized in many plants in which both ABA-dependent and ABA-independent signaling pathways are involved [75,76,77]. High salinity concentration causes cell osmotic change and water loss, and eventually damages the cell. Salt tolerance mechanisms involve maintaining the homeostasis of cellular ions and osmotic adjustment and ROS scavenging [78]. Overexpression of a salt stress-induced dehydrin DHN-5 from Durum wheat was found to enhance salt tolerance in transgenic Arabidopsis. Furthermore, numerous dehydrin genes have been identified to respond to salt stress, revealing the relationship between salt stress and dehydrin on a larger scale, as reported by omics studies [79,80,81]. Further overexpression and interference with dehydrin genes in transgenic plants validated association of dehydrin with salt tolerance in plants [10,19,49,81,82,83,84].
The osmotic pressure induced by salt stress leads to an increase in the concentration of Ca2+ and inositol 1,4,5-triphosphate (IP3) in the cytosol [85]. As secondary messengers, Ca2+ and IP3 then activate the mitogen-activated protein kinase (MAPK) cascades to regulate phosphorylation of various transcription factors including CBF/DREB, ABF, bZIP, Myc/MYB, and NAC (NAM, ATAF, CUC) factors [86]. Analyses of known dehydrin promoters have also shown many vital cis-elements [17,57]. One putative DRE element (ACCGAC) and one putative MYB transcription factor binding element (YAACKG/CANNTG), corresponding to MYB-binding sequences in Arabidopsis, were predicted in an isolated dehydrin AmDHN1 promoter [87]. However, there is no clear evidence regarding the interaction between these transcription factors and dehydrin promoters.
These results reflect the intricacy of the expression pattern of dehydrins under drought, cold and salt stresses. These motifs greatly contribute to the regulation of dehydrin genes under various abiotic stresses. Additionally, some other phytohormone responsive elements are also found in dehydrin promoters. However, explicit regulation of dehydrin genes requires further studies to elucidate precise mechanisms involved.
3.5. Heavy Metal Toxicity and Dehydrins
Studies on the functional roles of dehydrin in heavy metal toxicity are limited in plants. However, there is evidence that implies important role of dehydrins in plant against heavy metal stress. An isolated SK2-type dehydrin gene PvSR3 was highly induced by heavy metal stress [88]. Similarly, two Brassica juncea dehydrin genes, BjDHN2 and BjDHN3, were both induced under heavy metal stress. Moreover, overexpression of BjDHN2 and BjDHN3 in transgenic tobacco plants significantly enhanced their tolerance to heavy metal [89]. Dehydrins exhibited metal binding behavior in in vitro experiments, which uncovers potential mechanisms of plant tolerance to heavy metal. A binding property test of a dehydrin CuCOR15 implemented using immobilized metal ion affinity chromatography (IMAC) demonstrated that CuCOR15 was capable of binding to Fe3+, Co2+, Ni2+, Cu2+ and Zn2+ ions. Further truncated assay also indicated that HKGEHHSGDHH was the core sequence for binding [90]. Another in vitro study on dehydrin AtHIRD11, showed a similar metal binding behavior with CuCOR15. Subsequent amino acid mutation assay suggested the presence of histidine in AtHIRD11, which has an important role due to its metal binding property. Since both CuCOR15 and AtHIRD11 are radical-scavenging proteins, the results imply that dehydrins bind metals through a specific sequence containing histidine to reduce metal toxicity in plant.
An understanding of regulatory networks is essential for elucidating a full vision of associated links between heavy metal tolerance and dehydrins. DREB transcription factor family can play important roles in several abiotic stresses, including drought, freezing and heavy metal. Overexpression of DREB in transgenic plants showed activated expression of dehydrins such as COR15a, RD29A, RD29B, leading to improved tolerance to drought, heat and heavy metals [91,92,93,94,95]. Several pieces of evidence have unveiled close relationships between dehydrins and plant tolerance to heavy metals. However, further studies about interaction of dehydrin promoters and transcription factors are extremely necessary for deep understanding of mechanism of action.
Here we provide an overview of the dehydrins involved in abiotic stress signaling, as shown in Table 1. Although possible regulatory pathways under various abiotic stresses in plants have not been specifically unified, they still provide credible directions for studies on improving the plant tolerance against stresses.

Table 1.
An overview of dehydrins involved in abiotic stress signaling.
4. Localizations and Functions of Dehydrins
4.1. The Localization of Dehydrins
The localization and functional versatility of dehydrins has been widely studied with respect to plant tolerance against multiple abiotic stresses. Dehydrins are ubiquitously localized in different parts of cells, such as cytoplasm, nucleus, chloroplasts, vacuole, endoplasmic reticulum, mitochondria, cytoplasm and cell membranes [9,96,97,98,99,100,101,102]. Some dehydrins are also found to be localized in several parts of cells under abiotic stresses. GFP-fused dehydrin WZY2 is observed in both the nucleus and cytoplasm under cold stress [9]. Correspondingly, DHN24 is detected to be localized in cytosol and close to organelle membranes through immuno-electron microscopy [103].
As mentioned above, histidine-rich motifs in dehydrins may control its nuclear localization. OpsDHN1 is a nuclear-localized protein, whereas deletion of its histidine-rich segment results in significantly lower signal detection in the nucleus of OpsDHN1. Further studies on OpsDHN1 revealed another vital segment of dehydrin, S-segment, which is probably relevant for OpsDHN1 nuclear localization. In silico analysis also predicted a putative nuclear localization signal (NLS) containing partial of OpsDHN1 S-segment using cNLS mapper [97,104]. A report on CuCOR15 suggests that the phosphorylation state of serine in dehydrin S-segment is involved in its translocation to the nucleus. In addition, S-segment is activated by phosphorylation of casein kinase 2 (CK2) in Arabidopsis [105]. Meanwhile, phosphorylation of RAB17 was significant in binding nuclear localization signal (NLS) peptides in vitro [106]. As they are localized throughout the cells, dehydrins may provide more extensive support in protecting cell membranes and biomolecules under abiotic stresses.
4.2. The Multifunction of Dehydrins
The ubiquitous localization of dehydrins suggests their versatile functions in plant tolerance to abiotic stresses. Transgenic studies on dehydrin function imply a wide consensus that dehydrins are positively associated with tolerance of the plants against abiotic stress. It is reported that PpDHNB can enhance the cold tolerance in transgenic tobacco [15]. Overexpression of DHN-5 confers salt tolerance to transgenic Arabidopsis with higher expression of P5CS, proline, CAT, SOD, and POD [10]. Similar to PpDHNB and DHN-5, ShDHN significantly improves the tolerance of transgenic tomatoes under drought and cold stresses, leading to retention of higher relative water content and less ROS accumulation in leaves. The MDA contents in transgenic lines showed a lower level than that of wild-type tomatoes [49]. On the other hand, silencing of CaDHN1 resulted in lower accumulation of chlorophyll and expression of POD and Mn-SOD in Capsicum annuum under multiple abiotic stresses [81]. The above results provide a macroscopic view of dehydrin functions in enhancement of tolerance of plants at physiological level.
Protection of biomolecules is the main function of dehydrins during stress, as revealed by in vitro and in vivo studies. Overexpression of dehydrin genes usually improves the RWC in transgenic plants [49]. Amphipathic α-helix assists in bounding of dehydrin and water in cells [45,46,96,107], while on the other hand, dehydrins can stabilize cell structures and organelles to prevent water loss during stress [108,109]. In wheat dehydrin WZY2, K-segment is essential for the protecting activity of lactate dehydrogenase (LDH) and sustaining the survival rate of transformed E. coli under temperature stress [9]. A similar pattern was observed in DHN1a from Vitis vinifera and DHN5 from Rhododendron catawbiense, the lack of K-segment in which leads to a reduction in the effects of enzyme and bacterial protection [110]. The histidine-rich motifs in some dehydrins have been reported to participate in the binding function. Dehydrin CuCOR15 has been reported to bind DNA, and the presence of Zn2+ could strongly promote the ultimate binding [111]. Using an electrophoretic mobility shift assay, dehydrin DHN1a with a histidine-rich motif revealed weak interaction with double-strand DNA fragments. However, none of the divalent metals were capable of enhancing interaction [110].
In addition, some dehydrins with metal-binding capacity are regarded as ROS scavengers under abiotic stresses. In a common ROS-generation system, Cu2+ acts as an electron donor, promoting the variation from O2 to O2−, H2O2 and eventual ·OH in the presence of ascorbate. The addition of dehydrin AtHIRD11 could limit free Cu2+ to inhibit ROS generation. Meanwhile, CD spectrum implied a conformational change in AtHIRD11 participating in the inhibition of ROS synthesis [112]. The structural studies within AtHIRD11 suggested a strong relation between conformational changes and metal-chelating in ROS scavenging.
K-segment and histidine-rich motifs play vital roles in interaction, not with only enzymes and DNA, but also with phospholipids, which provide a potential structural foundation in membrane binding under various stresses [38]. A recent study has elucidated the pattern of dehydrin binding with lipid membrane. Dehydrin LTI30 contains K-segment with flanking His residues. NMR spectrum implies that LTI30 are membrane vesicles binding protein and require positively charged residues. Further study also suggested that LTI30 could be modulated by phosphorylation at predicted sites within K-segments. These results imply that protonation of His residues and phosphorylation of K-segments have key roles in lipid vesicle binding of dehydrins [33]. Although not all dehydrins contain histidine-rich motifs, above studies imply an important feature of dehydrin function.
Some possible mechanistic models in dehydrin functioning have been proposed. Dehydrins/LEA proteins may bind to liposomes [20,46,113] or sequester metal ions to form dehydrin-metal ion complex in order to interact with other biomolecules [42,43,47,112]. Recent studies have uncovered that several dehydrins form homo- and heterodimeric complexes, or even multimers, to bind and protect biomolecules for purpose of maintaining the cell and organelle structures and adjusting regular cell processes under adverse conditions of stress.
The capability of dehydrins to bind to multiple ligands provides protection to plants against abiotic stresses. Although the function of K-segment has been widely studied, the physiology of the other two conserved motifs, S- and Y-segments, is still ambiguous. Further studies on the functions of dehydrins are required to strengthen the existing evidence, and supporting data is needed to uncover the potential of dehydrins in plant physiology, especially under stress conditions.
5. Conclusions
The interest in exploring the association between dehydrins and abiotic stresses has been increasing recently, as it is very important to elucidate how plants respond to environmental stresses.
The molecular structures, functions and localizations of dehydrins have been widely explored, indicating their potential role in stress tolerance in plants. Potential regulatory mechanisms of dehydrins associated with various abiotic stresses are discussed in this article. However, description of stress-induced regulatory networks of dehydrins is scarce. Further studies on dehydrins should mainly focus on upstream mechanisms, especially with regard to identification of transcriptional cascades and regulatory mechanism of dehydrin genes. This will definitely be helpful in the breeding of stress-tolerant plants in the future to increase global crop production.
Author Contributions
Z.Y. was responsible for the selection, organization, and summary of published literature to which reference is made in this review. X.W. was responsible for the selection, organization, and summary of published literature to which reference is made in this review. L.Z. was responsible for the selection, organization, and summary of published literature to which reference is made in this review.
Funding
This work was supported by projects from National Natural Science Foundation of China (Grant No. 31671608).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Galau, G.A.; Hughes, D.W.; Dure, L., 3rd. Abscisic acid induction of cloned cotton late embryogenesis-abundant (Lea) mRNAs. Plant Mol. Biol. 1986, 7, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Velazquez, C.L.; Reyes, J.L.; Covarrubias, A.A. Group 4 late embryogenesis abundant proteins as a model to study intrinsically disordered proteins in plants. Plant Signal. Behav. 2017, 12, e1343777. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, L.; Xing, X.; Sun, L.; Pan, J.; Kong, X.; Zhang, M.; Li, D. ZmLEA3, a multifunctional group 3 LEA protein from maize (Zea mays L.), is involved in biotic and abiotic stresses. Plant Cell Physiol. 2013, 54, 944–959. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic LEA proteins and other hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Lv, A.; Fan, N.; Xie, J.; Yuan, S.; An, Y.; Zhou, P. Expression of CdDHN4, a Novel YSK2-Type Dehydrin Gene from Bermudagrass, Responses to Drought Stress through the ABA-Dependent Signal Pathway. Front. Plant Sci. 2017, 8, 748. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, L.; Zhang, T.; Yang, X.; Li, D. Functional characterization of KS-type dehydrin ZmDHN13 and its related conserved domains under oxidative stress. Sci. Rep. 2017, 7, 7361. [Google Scholar] [CrossRef] [PubMed]
- Halder, T.; Upadhyaya, G.; Ray, S. YSK2 Type Dehydrin (SbDhn1) from Sorghum bicolor Showed Improved Protection under High Temperature and Osmotic Stress Condition. Front. Plant Sci. 2017, 8, 918. [Google Scholar] [CrossRef] [PubMed]
- Bao, F.; Du, D.; An, Y.; Yang, W.; Wang, J.; Cheng, T.; Zhang, Q. Overexpression of Prunus mume Dehydrin Genes in Tobacco Enhances Tolerance to Cold and Drought. Front. Plant Sci. 2017, 8, 151. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Zhang, L.; Lv, H.; Li, H.; Zhang, Y.; Xu, Y.; Yu, J. The K-segments of wheat dehydrin WZY2 are essential for its protective functions under temperature stress. Front. Plant Sci. 2015, 6, 406. [Google Scholar] [CrossRef] [PubMed]
- Saibi, W.; Feki, K.; Ben Mahmoud, R.; Brini, F. Durum wheat dehydrin (DHN-5) confers salinity tolerance to transgenic Arabidopsis plants through the regulation of proline metabolism and ROS scavenging system. Planta 2015, 242, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Perdiguero, P.; Collada, C.; Soto, A. Novel dehydrins lacking complete K-segments in Pinaceae. The exception rather than the rule. Front. Plant Sci. 2014, 5, 682. [Google Scholar] [CrossRef] [PubMed]
- Perdiguero, P.; Barbero, M.C.; Cervera, M.T.; Soto, A.; Collada, C. Novel conserved segments are associated with differential expression patterns for Pinaceae dehydrins. Planta 2012, 236, 1863–1874. [Google Scholar] [CrossRef] [PubMed]
- Richard, S.; Morency, M.J.; Drevet, C.; Jouanin, L.; Seguin, A. Isolation and characterization of a dehydrin gene from white spruce induced upon wounding, drought and cold stresses. Plant Mol. Biol. 2000, 43, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, X.; Lv, Q.; Zhu, D.; Qiu, T.; Xu, Y.; Bao, F.; He, Y.; Hu, Y. Physcomitrella Patens Dehydrins (PpDHNA and PpDHNC) Confer Salinity and Drought Tolerance to Transgenic Arabidopsis Plants. Front. Plant Sci. 2017, 8, 1316. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, T.; Upadhyaya, G.; Halder, T.; Mukherjee, A.; Majumder, A.L.; Ray, S. Different dehydrins perform separate functions in Physcomitrella patens. Planta 2017, 245, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; He, P.; Xu, Y.; Liu, Q.; Yang, Y.; Liu, S. Overexpression of CsLEA11, a Y3SK2-type dehydrin gene from cucumber (Cucumis sativus), enhances tolerance to heat and cold in Escherichia coli. AMB Express 2017, 7, 182. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Zhang, L.; Lv, H.; Zhang, H.; Zhang, D.; Wang, X.; Chen, J. The dehydrin wzy2 promoter from wheat defines its contribution to stress tolerance. Funct. Integr. Genom. 2014, 14, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; He, M.; Zhu, Z.; Li, S.; Xu, Y.; Zhang, C.; Singer, S.D.; Wang, Y. Identification of the dehydrin gene family from grapevine species and analysis of their responsiveness to various forms of abiotic and biotic stress. BMC Plant. Biol. 2012, 12, 140. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, L.; Svensson, J.; Carballo, V.; Izmendi, D.; Welin, B.; Vidal, S. A dehydrin gene in Physcomitrella patens is required for salt and osmotic stress tolerance. Plant J. 2006, 45, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Thalhammer, A.; Bryant, G.; Sulpice, R.; Hincha, D.K. Disordered cold regulated15 proteins protect chloroplast membranes during freezing through binding and folding, but do not stabilize chloroplast enzymes in vivo. Plant Physiol. 2014, 166, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Danyluk, J.; Kane, N.A.; Breton, G.; Limin, A.E.; Fowler, D.B.; Sarhan, F. TaVRT-1, a putative transcription factor associated with vegetative to reproductive transition in cereals. Plant Physiol. 2003, 132, 1849–1860. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, F.; Takumi, S.; Nakata, M.; Ohno, R.; Nakamura, T.; Nakamura, C. Comparative study of the expression profiles of the Cor/Lea gene family in two wheat cultivars with contrasting levels of freezing tolerance. Physiol. Plant. 2004, 120, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Kosova, K.; Holkova, L.; Prasil, I.T.; Prasilova, P.; Bradacova, M.; Vitamvas, P.; Capkova, V. Expression of dehydrin 5 during the development of frost tolerance in barley (Hordeum vulgare). J. Plant Physiol. 2008, 165, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Van Zee, K.; Chen, F.Q.; Hayes, P.M.; Close, T.J.; Chen, T. Cold-Specific Induction of a Dehydrin Gene Family Member in Barley. Plant Physiol. 1995, 108, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Graether, S.P.; Boddington, K.F. Disorder and function: A review of the dehydrin protein family. Front. Plant Sci. 2014, 5, 576. [Google Scholar] [CrossRef] [PubMed]
- Close, T.J. Dehydrins: Emergence of a biochemical role of a family of plant dehydration proteins. Physiol. Plant. 1996, 97, 795–803. [Google Scholar] [CrossRef]
- Jarvis, S.B.; Taylor, M.A.; Macleod, M.R.; Davies, H.V. Cloning and Characterisation of the cDNA Clones of three Genes that are differentially Expressed during Dormancy-Breakage in the Seeds of Douglas Fir (Pseudotsuga menziesii). J. Plant Physiol. 1996, 147, 559–566. [Google Scholar] [CrossRef]
- Baker, J.; Van Dennsteele, C.; Dure, L., 3rd. Sequence and characterization of 6 Lea proteins and their genes from cotton. Plant Mol. Biol. 1988, 11, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Geromanos, S.; Tempst, P.; Hartl, F.U. Identification of nucleotide-binding regions in the chaperonin proteins GroEL and GroES. Nature 1993, 366, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Mundy, J.; Chua, N.H. Abscisic acid and water-stress induce the expression of a novel rice gene. EMBO J. 1988, 7, 2279–2286. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Sanchez, I.E.; Martynowicz, D.M.; Rodriguez-Hernandez, A.A.; Perez-Morales, M.B.; Graether, S.P.; Jimenez-Bremont, J.F. A dehydrin-dehydrin interaction: The case of SK3 from Opuntia streptacantha. Front. Plant Sci. 2014, 5, 520. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Sanchez, I.E.; Maruri-Lopez, I.; Graether, S.P.; Jimenez-Bremont, J.F. In vivo evidence for homo- and heterodimeric interactions of Arabidopsis thaliana dehydrins AtCOR47, AtERD10, and AtRAB18. Sci. Rep. 2017, 7, 17036. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.K.; Kutzer, M.; Procek, J.; Grobner, G.; Harryson, P. Tunable membrane binding of the intrinsically disordered dehydrin Lti30, a cold-induced plant stress protein. Plant Cell 2011, 23, 2391–2404. [Google Scholar] [CrossRef] [PubMed]
- Kalderon, D.; Richardson, W.D.; Markham, A.F.; Smith, A.E. Sequence requirements for nuclear location of simian virus 40 large-T antigen. Nature 1984, 311, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.B.; Goday, A.; Figueras, M.; Jessop, A.C.; Pages, M. Phosphorylation mediates the nuclear targeting of the maize Rab17 protein. Plant J. 1998, 13, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.A.; Veltri, M.; Boddington, K.F.; Singh, K.K.; Graether, S.P. Genome Analysis of Conserved Dehydrin Motifs in Vascular Plants. Front. Plant Sci. 2017, 8, 709. [Google Scholar] [CrossRef] [PubMed]
- Rahman, L.N.; Chen, L.; Nazim, S.; Bamm, V.V.; Yaish, M.W.; Moffatt, B.A.; Dutcher, J.R.; Harauz, G. Interactions of intrinsically disordered Thellungiella salsuginea dehydrins TsDHN-1 and TsDHN-2 with membranes—Synergistic effects of lipid composition and temperature on secondary structure. Biochem. Cell Biol. 2010, 88, 791–807. [Google Scholar] [CrossRef] [PubMed]
- Szalaine Agoston, B.; Kovacs, D.; Tompa, P.; Perczel, A. Full backbone assignment and dynamics of the intrinsically disordered dehydrin ERD14. Biomol. NMR Assign. 2011, 5, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N. A decade and a half of protein intrinsic disorder: Biology still waits for physics. Protein Sci. 2013, 22, 693–724. [Google Scholar] [CrossRef] [PubMed]
- Tompa, P. Intrinsically unstructured proteins. Trends Biochem. Sci. 2002, 27, 527–533. [Google Scholar] [CrossRef]
- Mielke, S.P.; Krishnan, V.V. Characterization of protein secondary structure from NMR chemical shifts. Prog. Nucl. Magn. Reson. Spectrosc. 2009, 54, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Monna, S.; Murata, T.; Nakano, T.; Amano, S.; Nachbar, M.; Watzig, H. The Arabidopsis KS-type dehydrin recovers lactate dehydrogenase activity inhibited by copper with the contribution of His residues. Plant Sci. 2016, 245, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Rahman, L.N.; Bamm, V.V.; Voyer, J.A.; Smith, G.S.; Chen, L.; Yaish, M.W.; Moffatt, B.A.; Dutcher, J.R.; Harauz, G. Zinc induces disorder-to-order transitions in free and membrane-associated Thellungiella salsuginea dehydrins TsDHN-1 and TsDHN-2: A solution CD and solid-state ATR-FTIR study. Amino Acids 2011, 40, 1485–1502. [Google Scholar] [CrossRef] [PubMed]
- Halder, T.; Agarwal, T.; Ray, S. Isolation, cloning, and characterization of a novel Sorghum dehydrin (SbDhn2) protein. Protoplasma 2016, 253, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.; Graether, S.P. Cryoprotective mechanism of a small intrinsically disordered dehydrin protein. Protein Sci. 2011, 20, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.W.; Boddington, K.F.; Warnica, J.M.; Atkinson, J.; McKenna, S.; Madge, J.; Barker, C.H.; Graether, S.P. Structural and Functional Insights into the Cryoprotection of Membranes by the Intrinsically Disordered Dehydrins. J. Biol. Chem. 2015, 290, 26900–26913. [Google Scholar] [CrossRef] [PubMed]
- Mu, P.; Feng, D.; Su, J.; Zhang, Y.; Dai, J.; Jin, H.; Liu, B.; He, Y.; Qi, K.; Wang, H.; et al. Cu2+ triggers reversible aggregation of a disordered His-rich dehydrin MpDhn12 from Musa paradisiaca. J. Biochem. 2011, 150, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yu, C.; Li, H.; Ouyang, B.; Wang, T.; Zhang, J.; Wang, X.; Ye, Z. Overexpression of ShDHN, a dehydrin gene from Solanum habrochaites enhances tolerance to multiple abiotic stresses in tomato. Plant Sci. 2015, 231, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Zhang, L.; Liu, C.; He, L.; Wang, A.; Liu, H.L.; Zhu, J.B. Cloning and characterization of a novel dehydrin gene, SiDhn2, from Saussurea involucrata Kar. et Kir. Plant Mol. Biol. 2014, 84, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Nordin, K.; Vahala, T.; Palva, E.T. Differential expression of two related, low-temperature-induced genes in Arabidopsis thaliana (L.) Heynh. Plant Mol. Biol. 1993, 21, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Busk, P.K.; Pages, M. Protein binding to the abscisic acid-responsive element is independent of VIVIPAROUS1 in vivo. Plant Cell 1997, 9, 2261–2270. [Google Scholar] [CrossRef] [PubMed]
- Ono, A.; Izawa, T.; Chua, N.H.; Shimamoto, K. The rab16B promoter of rice contains two distinct abscisic acid-responsive elements. Plant Physiol. 1996, 112, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Straub, P.F.; Shen, Q.; Ho, T.D. Structure and promoter analysis of an ABA- and stress-regulated barley gene, HVA1. Plant Mol. Biol. 1994, 26, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.H.; Li, C.W.; Su, R.C.; Cheng, C.P.; Sanjaya; Tsai, Y.C.; Chan, M.T. A tomato bZIP transcription factor, SlAREB, is involved in water deficit and salt stress response. Planta 2010, 231, 1459–1473. [Google Scholar] [CrossRef] [PubMed]
- Bassett, C.L.; Wisniewski, M.E.; Artlip, T.S.; Richart, G.; Norelli, J.L.; Farrell, R.E., Jr. Comparative expression and transcript initiation of three peach dehydrin genes. Planta 2009, 230, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Zolotarov, Y.; Stromvik, M. De Novo Regulatory Motif Discovery Identifies Significant Motifs in Promoters of Five Classes of Plant Dehydrin Genes. PLoS ONE 2015, 10, e0129016. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Jha, B.; Agarwal, P.K. A dehydration-responsive element binding (DREB) transcription factor from the succulent halophyte Salicornia brachiata enhances abiotic stress tolerance in transgenic tobacco. Mar. Biotechnol. 2014, 16, 657–673. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, L.; Wang, L.; Wang, S.; Cheng, X. VrDREB2A, a DREB-binding transcription factor from Vigna radiata, increased drought and high-salt tolerance in transgenic Arabidopsis thaliana. J. Plant Res. 2016, 129, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Shukla, A.; Upadhyay, S.; Sanchita; Sharma, P.; Singh, S.; Phukan, U.J.; Meena, A.; Khan, F.; Tripathi, V.; et al. Identification, occurrence, and validation of DRE and ABRE Cis-regulatory motifs in the promoter regions of genes of Arabidopsis thaliana. J. Integr. Plant Biol. 2014, 56, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Narusaka, Y.; Nakashima, K.; Shinwari, Z.K.; Sakuma, Y.; Furihata, T.; Abe, H.; Narusaka, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Interaction between two cis-acting elements, ABRE and DRE, in ABA-dependent expression of Arabidopsis rd29A gene in response to dehydration and high-salinity stresses. Plant J. 2003, 34, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Janska, A.; Marsik, P.; Zelenkova, S.; Ovesna, J. Cold stress and acclimation—What is important for metabolic adjustment? Plant Biol. 2010, 12, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Mondini, L.; Nachit, M.M.; Pagnotta, M.A. Allelic variants in durum wheat (Triticum turgidum L. var. durum) DREB genes conferring tolerance to abiotic stresses. Mol. Genet. Genom. 2015, 290, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.G.; Cook, D.; Thomashow, M.F. Low temperature induction of Arabidopsis CBF1, 2, and 3 is gated by the circadian clock. Plant Physiol. 2005, 137, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Srivasta, A.; Mehta, S.; Lindlof, A.; Bhargava, S. Over-represented promoter motifs in abiotic stress-induced DREB genes of rice and sorghum and their probable role in regulation of gene expression. Plant Signal. Behav. 2010, 5, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Todaka, D.; Mizoi, J.; Yoshida, T.; Kidokoro, S.; Matsukura, S.; Takasaki, H.; Sakurai, T.; Yamamoto, Y.Y.; Yoshiwara, K.; et al. Identification of cis-acting promoter elements in cold- and dehydration-induced transcriptional pathways in Arabidopsis, rice, and soybean. DNA Res. 2012, 19, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Bokszczanin, K.L.; Solanaceae Pollen Thermotolerance Initial Training Network Consortium; Fragkostefanakis, S. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front. Plant Sci. 2013, 4, 315. [Google Scholar] [CrossRef] [PubMed]
- Drira, M.; Saibi, W.; Brini, F.; Gargouri, A.; Masmoudi, K.; Hanin, M. The K-segments of the wheat dehydrin DHN-5 are essential for the protection of lactate dehydrogenase and beta-glucosidase activities in vitro. Mol. Biotechnol. 2013, 54, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Brini, F.; Saibi, W.; Amara, I.; Gargouri, A.; Masmoudi, K.; Hanin, M. Wheat dehydrin DHN-5 exerts a heat-protective effect on beta-glucosidase and glucose oxidase activities. Biosci. Biotechnol. Biochem. 2010, 74, 1050–1054. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.X.; Qin, F. Dehydrins from wheat x Thinopyrum ponticum amphiploid increase salinity and drought tolerance under their own inducible promoters without growth retardation. Plant Physiol. Biochem. 2016, 99, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Sakuma, Y.; Todaka, D.; Maruyama, K.; Qin, F.; Mizoi, J.; Kidokoro, S.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of an Arabidopsis heat-shock transcription factor HsfA3 in the transcriptional cascade downstream of the DREB2A stress-regulatory system. Biochem. Biophys. Res. Commun. 2008, 368, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Lenis, J.M.; Ellersieck, M.; Blevins, D.G.; Sleper, D.A.; Nguyen, H.T.; Dunn, D.; Lee, J.D.; Shannon, J.G. Differences in Ion Accumulation and Salt Tolerance among Glycine Accessions. J. Agron. Crop Sci. 2011, 197, 302–310. [Google Scholar] [CrossRef]
- Huang, G.T.; Ma, S.L.; Bai, L.P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Phang, T.H.; Shao, G.; Lam, H.M. Salt tolerance in soybean. J. Integr. Plant Biol. 2008, 50, 1196–1212. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Nie, L.; Sun, G.; Guo, J.; Liu, Y. Cloning and characterization of dehydrin gene from Ammopiptanthus mongolicus. Mol. Biol. Rep. 2013, 40, 2281–2291. [Google Scholar] [CrossRef] [PubMed]
- Jyothi-Prakash, P.A.; Mohanty, B.; Wijaya, E.; Lim, T.M.; Lin, Q.; Loh, C.S.; Kumar, P.P. Identification of salt gland-associated genes and characterization of a dehydrin from the salt secretor mangrove Avicennia officinalis. BMC Plant Biol. 2014, 14, 291. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.G.; Jing, H.; Guo, W.L.; Wang, S.B.; Ma, F.; Pan, B.G.; Gong, Z.H. Silencing of dehydrin CaDHN1 diminishes tolerance to multiple abiotic stresses in Capsicum annuum L. Plant Cell Rep. 2015, 34, 2189–2200. [Google Scholar] [CrossRef] [PubMed]
- Saibi, W.; Zouari, N.; Masmoudi, K.; Brini, F. Role of the durum wheat dehydrin in the function of proteases conferring salinity tolerance in Arabidopsis thaliana transgenic lines. Int. J. Biol. Macromol. 2016, 85, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Shekhawat, U.K.; Srinivas, L.; Ganapathi, T.R. MusaDHN-1, a novel multiple stress-inducible SK(3)-type dehydrin gene, contributes affirmatively to drought- and salt-stress tolerance in banana. Planta 2011, 234, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Brini, F.; Hanin, M.; Lumbreras, V.; Amara, I.; Khoudi, H.; Hassairi, A.; Pages, M.; Masmoudi, K. Overexpression of wheat dehydrin DHN-5 enhances tolerance to salt and osmotic stress in Arabidopsis thaliana. Plant Cell Rep. 2007, 26, 2017–2026. [Google Scholar] [CrossRef] [PubMed]
- DeWald, D.B.; Torabinejad, J.; Jones, C.A.; Shope, J.C.; Cangelosi, A.R.; Thompson, J.E.; Prestwich, G.D.; Hama, H. Rapid accumulation of phosphatidylinositol 4,5-bisphosphate and inositol 1,4,5-trisphosphate correlates with calcium mobilization in salt-stressed arabidopsis. Plant Physiol. 2001, 126, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, W.Y.; Yun, D.J. A New Insight of Salt Stress Signaling in Plant. Mol. Cells 2016, 39, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.A.; Rebala, K.C.; Venkataraman, G.; Parida, A. A diurnally regulated dehydrin from Avicennia marina that shows nucleo-cytoplasmic localization and is phosphorylated by Casein kinase II in vitro. Plant Physiol. Biochem. 2009, 47, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Yu, F.; Cong, L.; Wang, L.; Burkard, G.; Chai, T. Cloning and expression analysis of SKn-type dehydrin gene from bean in response to heavy metals. Mol. Biotechnol. 2006, 32, 205–218. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.X.; Wei, W.; Han, L.; Guan, Z.Q.; Wang, Z.; Chai, T.Y. BjDHNs confer heavy-metal tolerance in plants. Mol. Biotechnol. 2008, 38, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Fujinaga, M.; Kuboi, T. Metal binding by citrus dehydrin with histidine-rich domains. J. Exp. Bot. 2005, 56, 2695–2703. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guan, Y.; Wu, Y.; Chen, H.; Chen, F.; Chu, C. Overexpression of a rice OsDREB1F gene increases salt, drought, and low temperature tolerance in both Arabidopsis and rice. Plant Mol. Biol. 2008, 67, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.S.; Ni, Z.Y.; Li, Z.Y.; Li, L.C.; Chen, M.; Gao, D.Y.; Yu, X.D.; Liu, P.; Ma, Y.Z. Isolation and functional characterization of HvDREB1-a gene encoding a dehydration-responsive element binding protein in Hordeum vulgare. J. Plant Res. 2009, 122, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Ban, Q.; Liu, G.; Wang, Y. A DREB gene from Limonium bicolor mediates molecular and physiological responses to copper stress in transgenic tobacco. J. Plant Physiol. 2011, 168, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, D.; Li, H.; Wang, Y.; Zhang, Y.; Wood, A.J. EsDREB2B, a novel truncated DREB2-type transcription factor in the desert legume Eremosparton songoricum, enhances tolerance to multiple abiotic stresses in yeast and transgenic tobacco. BMC Plant Biol. 2014, 14, 44. [Google Scholar] [CrossRef] [PubMed]
- Hadi, F.; Gilpin, M.; Fuller, M.P. Identification and expression analysis of CBF/DREB1 and COR15 genes in mutants of Brassica oleracea var. botrytis with enhanced proline production and frost resistance. Plant Physiol. Biochem. 2011, 49, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Kalemba, E.M.; Litkowiec, M. Functional characterization of a dehydrin protein from Fagus sylvatica seeds using experimental and in silico approaches. Plant Physiol. Biochem. 2015, 97, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Sanchez, I.E.; Maruri-Lopez, I.; Ferrando, A.; Carbonell, J.; Graether, S.P.; Jimenez-Bremont, J.F. Nuclear localization of the dehydrin OpsDHN1 is determined by histidine-rich motif. Front. Plant Sci. 2015, 6, 702. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sun, X.; Yang, S.; Li, X.; Yang, Y. Molecular cloning and characterization of a novel SK3-type dehydrin gene from Stipa purpurea. Biochem. Biophys. Res. Commun. 2014, 448, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Pochon, S.; Simoneau, P.; Pigne, S.; Balidas, S.; Bataille-Simoneau, N.; Campion, C.; Jaspard, E.; Calmes, B.; Hamon, B.; Berruyer, R.; et al. Dehydrin-like proteins in the necrotrophic fungus Alternaria brassicicola have a role in plant pathogenesis and stress response. PLoS ONE 2013, 8, e75143. [Google Scholar] [CrossRef] [PubMed]
- Farias-Soares, F.L.; Burrieza, H.P.; Steiner, N.; Maldonado, S.; Guerra, M.P. Immunoanalysis of dehydrins in Araucaria angustifolia embryos. Protoplasma 2013, 250, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Ruibal, C.; Salamo, I.P.; Carballo, V.; Castro, A.; Bentancor, M.; Borsani, O.; Szabados, L.; Vidal, S. Differential contribution of individual dehydrin genes from Physcomitrella patens to salt and osmotic stress tolerance. Plant Sci. 2012, 190, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Carjuzaa, P.; Castellion, M.; Distefano, A.J.; del Vas, M.; Maldonado, S. Detection and subcellular localization of dehydrin-like proteins in quinoa (Chenopodium quinoa Willd.) embryos. Protoplasma 2008, 233, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Szabala, B.M.; Fudali, S.; Rorat, T. Accumulation of acidic SK(3) dehydrins in phloem cells of cold- and drought-stressed plants of the Solanaceae. Planta 2014, 239, 847–863. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Matsumura, N.; Takashima, H.; Miyamoto-Sato, E.; Tomita, M.; Yanagawa, H. Six classes of nuclear localization signals specific to different binding grooves of importin alpha. J. Biol. Chem. 2009, 284, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Alsheikh, M.K.; Heyen, B.J.; Randall, S.K. Ion binding properties of the dehydrin ERD14 are dependent upon phosphorylation. J. Biol. Chem. 2003, 278, 40882–40889. [Google Scholar] [CrossRef] [PubMed]
- Close, T.J.; Kortt, A.A.; Chandler, P.M. A cDNA-based comparison of dehydration-induced proteins (dehydrins) in barley and corn. Plant Mol. Biol. 1989, 13, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.; Eremina, N.; Barth, A.; Danielsson, J.; Harryson, P. Membrane-Induced Folding of the Plant Stress Dehydrin Lti30. Plant Physiol. 2016, 171, 932–943. [Google Scholar] [PubMed]
- Ismail, A.M.; Hall, A.E.; Close, T.J. Purification and partial characterization of a dehydrin involved in chilling tolerance during seedling emergence of cowpea. Plant Physiol. 1999, 120, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Koag, M.C.; Wilkens, S.; Fenton, R.D.; Resnik, J.; Vo, E.; Close, T.J. The K-segment of maize DHN1 mediates binding to anionic phospholipid vesicles and concomitant structural changes. Plant Physiol. 2009, 150, 1503–1514. [Google Scholar] [CrossRef] [PubMed]
- Rosales, R.; Romero, I.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. The crucial role of Phi- and K-segments in the in vitro functionality of Vitis vinifera dehydrin DHN1a. Phytochemistry 2014, 108, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Shinoda, Y.; Tanaka, Y.; Kuboi, T. DNA binding of citrus dehydrin promoted by zinc ion. Plant Cell Environ. 2009, 32, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Kondo, M.; Kato, T. A KS-type dehydrin and its related domains reduce Cu-promoted radical generation and the histidine residues contribute to the radical-reducing activities. J. Exp. Bot. 2013, 64, 1615–1624. [Google Scholar] [CrossRef] [PubMed]
- Bremer, A.; Wolff, M.; Thalhammer, A.; Hincha, D.K. Folding of intrinsically disordered plant LEA proteins is driven by glycerol-induced crowding and the presence of membranes. FEBS J. 2017, 284, 919–936. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).