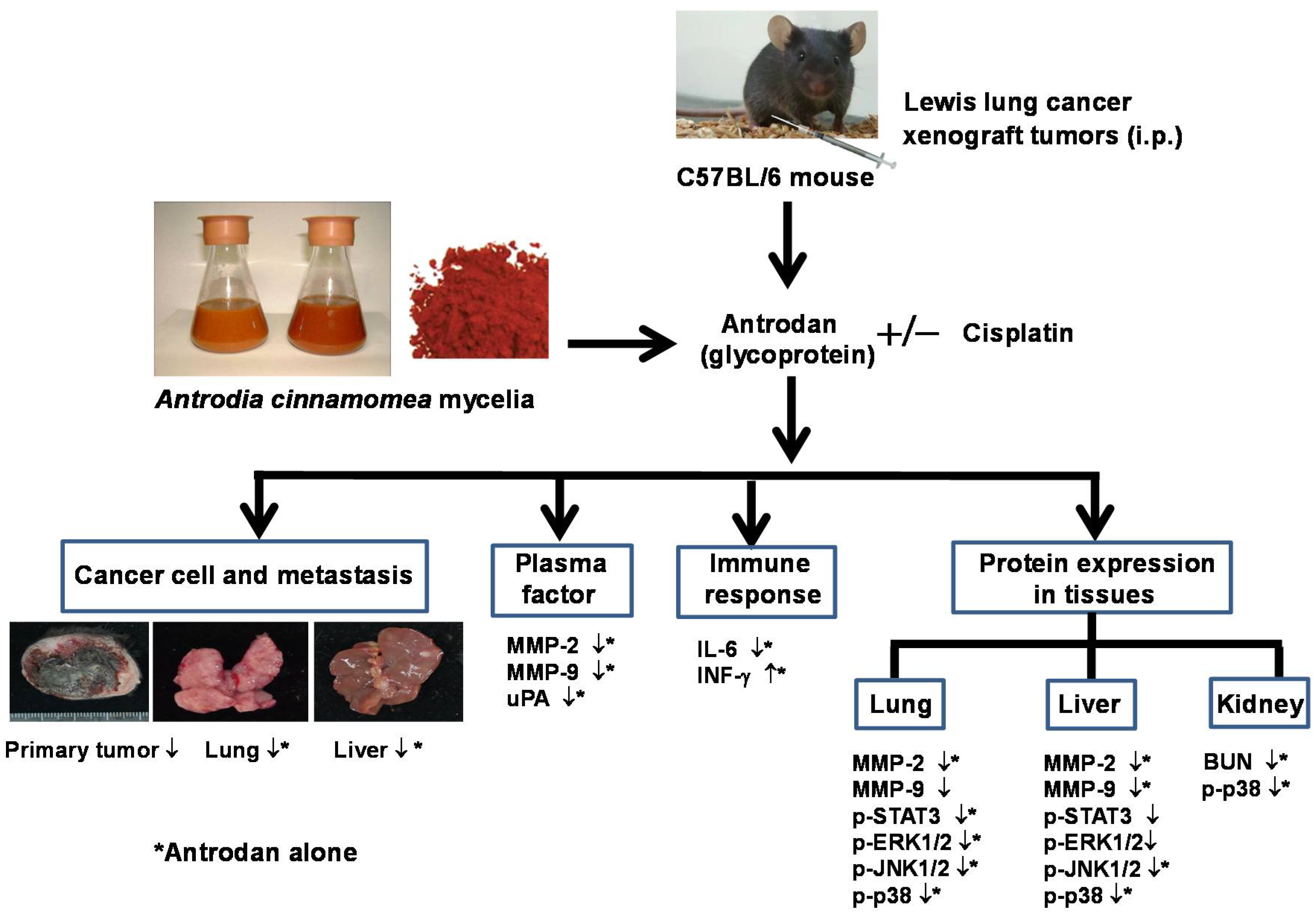
1. Introduction
Metastatic cancers are one of the leading causes of death. Statistical data indicate that tumor metastasis is responsible for approximately 90% of all cancer-related deaths [
1]. The metastasis process consists of a complex cascade of events, including angiogenesis, migration, and invasion. In the tumor microenvironment, matrix metalloproteinases (MMPs) degrade the extracellular matrix, and secrete the vascular endothelial growth factor (VEGF) responsible for providing nutrients to cancer cells, and for inducing vascular proliferation. These tumor cells may undergo migration and invasion, traveling through the blood or lymphatic vessels, eventually leaving the blood or lymphatic vessels, before adhering to and growing at a distal site [
2,
3].
Among the biomarkers of cancer metastasis, MMPs, a family of zinc-dependent enzymes, play an important role in many physiological functions, such as embryonic development, wound healing, angiogenesis, tissue remodeling, and the regulation of inflammatory responses [
4,
5]. MMP-2 and MMP-9 are highly expressed in malignant tumors with a high metastatic capacity. These proteinases are considered the key factors contributing to cancer cell invasion and cancer metastasis [
6]. The formation of active MMPs may be linked to effects caused by the urokinase plasminogen activator (uPA). uPA, a serine protease, is often overexpressed in non-small-cell lung cancers, liver cancers, colorectal cancers, and breast cancers, and is recognized as being responsible for poor prognoses and cancer metastasis [
7,
8,
9]. In the tumor microenvironment, signal transducer and activator of transcription (STAT) proteins also play a vital role in promoting or inhibiting tumor metastasis. STAT3 proteins have been reported to downregulate expression of the vascular endothelial growth factor (VEGF), thereby reducing angiogenesis and delaying progress of the tumor [
10]. As the STAT3 pathway is mainly responsible for the regulation of cell proliferation, invasion, and angiogenesis, many reports have shown that its sustained activation promotes the metastasis of many cancers, including thymic tumors, colorectal cancers, and squamous cell carcinomas of the skin. Thus, the STAT3 pathway is thought to be associated with cancer metastasis, and the suppression of this pathway can be used as one of the strategies in cancer therapy [
11,
12]. In addition to the STAT3 pathway, related transcription factors named mitogen-activated protein kinases (MAPKs) are involved in cell proliferation, apoptosis, angiogenesis, migration, and metastasis. The MAPK pathway can be divided into three subclasses: extracellular signal-regulated kinases 1 and 2 (ERK1/2), c-Jun N-terminal kinases (JNKs), and p38 pathways. The ERK1/2-MAPK pathway can be activated in a number of ways, including through growth factors, cytokines, viral infections, G-coupled protein receptors, and carcinogenic factors [
13]. JNK proteins regulate three genes—MAPK8, MAPK9, and MAPK10—which have been found to produce JNK1, JNK2, and JNK3 proteins, respectively. JNK1 and JNK2 are expressed in most cells, while JNK3 is mainly present in the brain. JNKs primarily affect the expression of the AP1 transcription factor. The oncogenic functions of JNKs are mostly based on their ability to phosphorylate Jun, and to activate AP1, thus promoting cancer cell growth [
14].
Cisplatin, one of many well-known anti-cancer drugs, was approved for cancer treatment by the Food and Drug Administration (FDA) in 1979 [
15]. One of the major functions of cisplatin in cancer therapy is attributed to its ability to suppress the DNA replication of cancer cells [
15,
16]. However, most questions concerning the treatment of cancer with cisplatin arise from its toxicity, which induces serious side effects, such as gastrotoxicity, myelosuppression, nephrotoxicity, chronic renal failure, etc. [
17]. As such, there exists substantial interest in the development of nutraceuticals for combined treatment with chemotherapy, so as to reduce the aforementioned side effects.
Polysaccharides have been widely applied in cancer treatments, for example, polysaccharide-K (Krestin
®, Kureha Corporation, Tokyo, Japan) from the
Coriolus versicolor mushroom used in the treatments of colorectal and breast cancers [
18,
19], and lentinan from the shiitake mushroom used in the treatment of gastric cancer [
20]. Antrodan, an immunomodulatory protein-bound polysaccharide prepared from fungal polysaccharides in the mycelia of
Antrodia cinnamomea, was applied in various biological contexts, including as an anti-inflammatory drug and an antioxidant [
21], in the beneficial treatment of benign prostatic hyperplasia [
22], and for its protective effects against lipopolysaccharide-induced acute liver damage in rat models [
23]. Furthermore, antrodan inhibited the metastasis of Lewis lung carcinomas in vitro, through upregulation of the protein expression of tissue inhibitor of matrix metalloproteinase (TIMP)-1, TIMP-2, and nm23-H1, leading to a decrease in the activities and protein expression of MMP-2 and MMP-9 [
24]. We also found that antrodan acted through indirect immunomodulatory effects, by increasing interleukin (IL)-12 and IL-1β levels, and by decreasing tumor necrosis factor alpha (TNF-α), IL-6, and IL-8 levels in experiments using mononuclear cells [
24]. However, it was unclear whether antrodan was effective in the inhibition of tumor metastasis in vivo. In this study, Lewis lung carcinoma (LLC) cells, from a highly invasive murine lung cancer cell line, were used to investigate the anti-metastatic activities of antrodan, as well as those in combination with cisplatin, to explore whether antrodan had the potential to act as an adjuvant of cisplatin in anti-cancer therapy.
3. Discussion
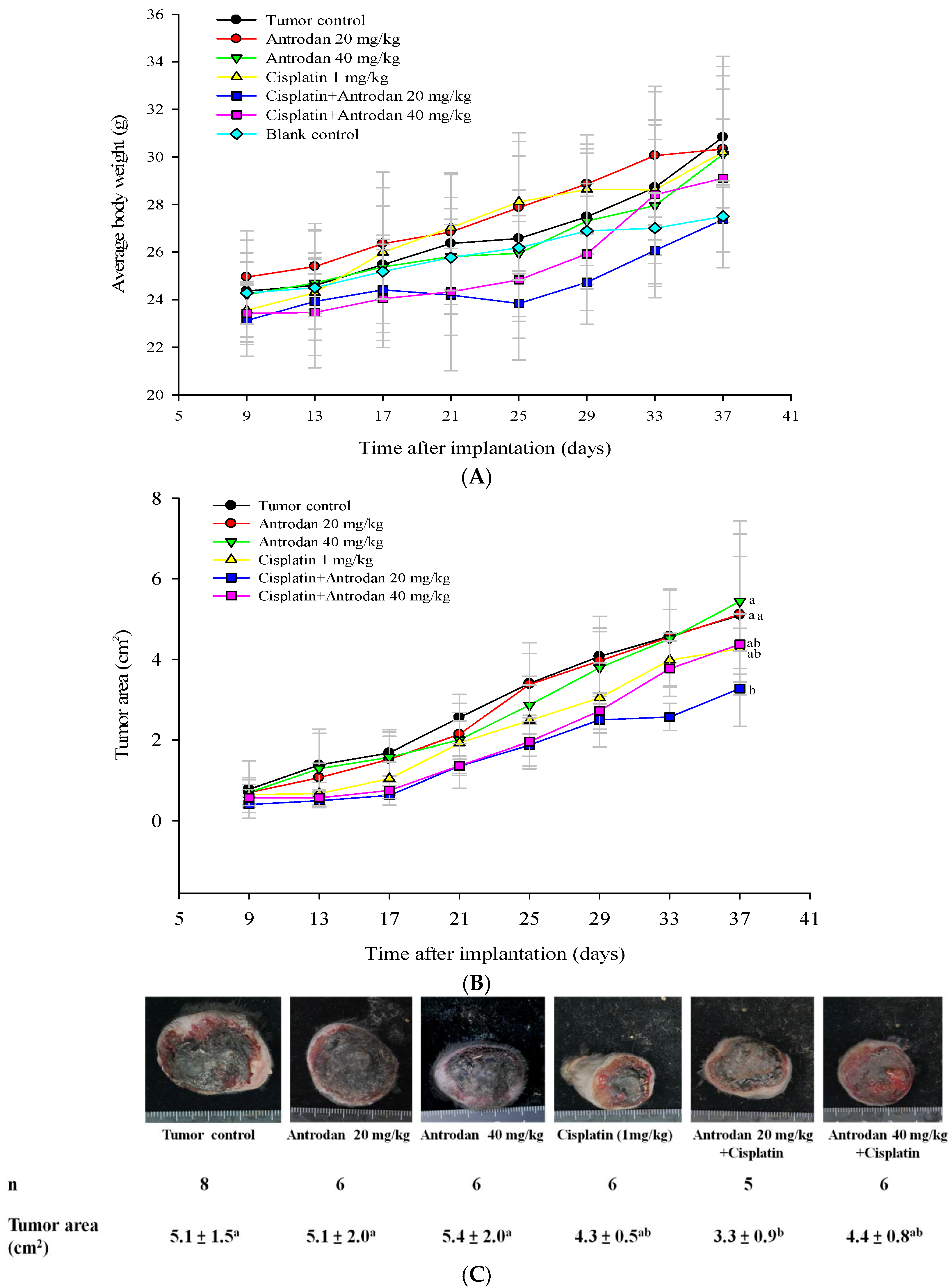
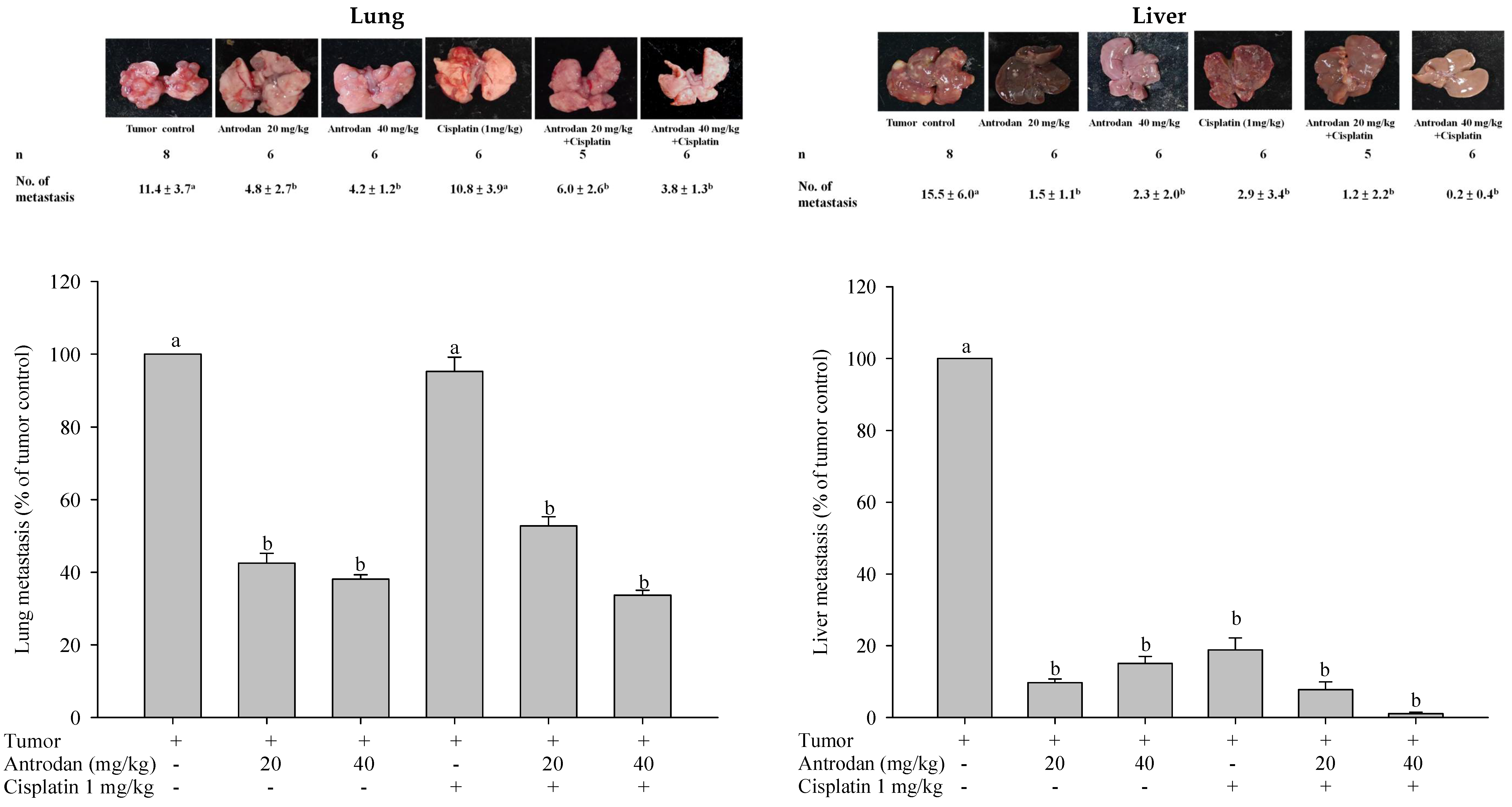
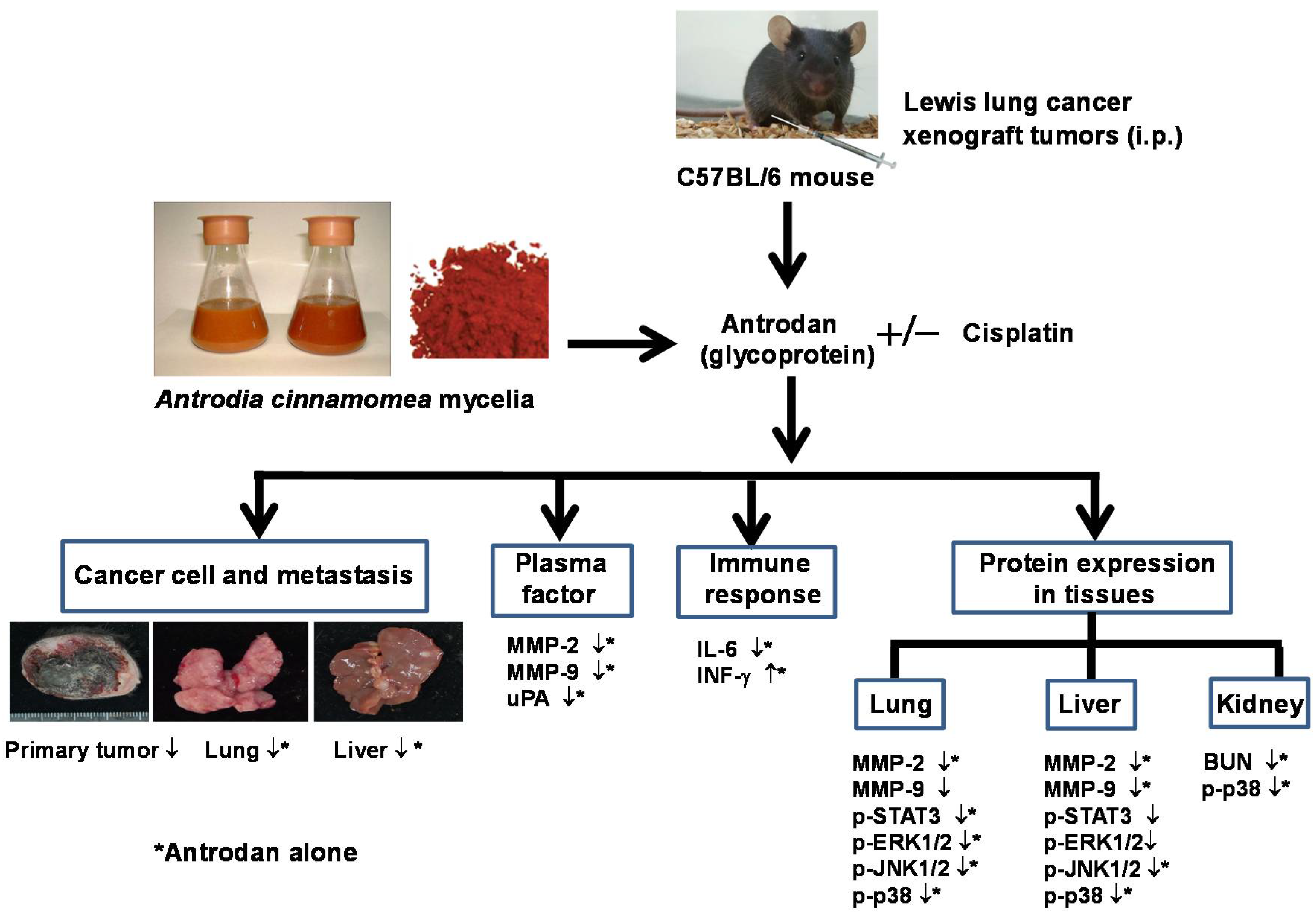
In this study, we examined the potential of antrodan to both enhance the anti-cancer effects of cisplatin and reduce its side effects in LLC-bearing mice. This study reports, for the first time, that treatment with antrodan only effectively inhibited both lung and liver metastasis, and enhanced the effects of cisplatin on lung metastasis when both agents were combined. Antrodan itself did not inhibit primary tumor growth, nor did it disturb the effect of tumor growth inhibition by cisplatin. Unexpectedly, the effects of treatment with cisplatin only, and the combined treatment of antrodan (40 mg/kg) and cisplatin were less, but not significantly less, than the effect of the combined treatment of antrodan (20 mg/kg) and cisplatin on reduction of the primary tumor area. This non-dose-dependent effect could be attributed to the relatively large variation in tumor area, and the relatively small number (5 to 6) of rats per group. In addition, we found that antrodan effectively prevented cisplatin-induced kidney dysfunction in this animal model.
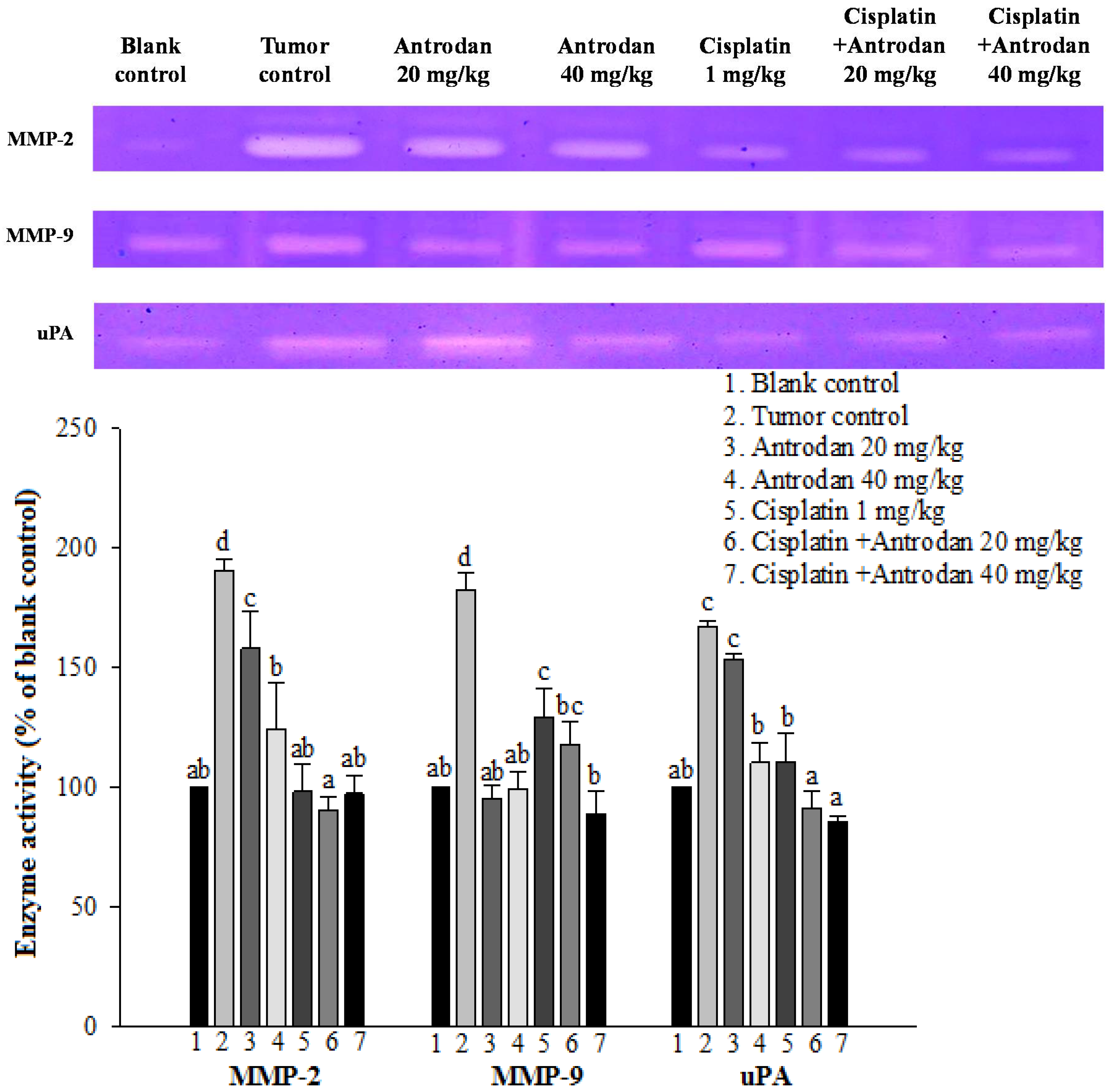
Possible molecular mechanisms underlying the anti-metastatic effects of antrodan on signal transduction pathways in LLC-bearing mice were elucidated in this study. Firstly, the effects of antrodan were observed on tumor invasion, and metastasis-involved matrix metalloproteinases (MMPs) and serine-proteinases, such as the urokinase plasminogen activator (uPA) [
25]. Among the MMPs, MMP-2 and MMP-9 specifically degrade the main structural component of basement membranes [
26]. When bound to its receptor, uPA initiates the activation of MMPs. Once activated, MMPs digest the extracellular matrix enzymes (ECMs), therefore facilitating the movement of cancer cells. In addition, overexpression of uPA has been demonstrated to be essential in the maintenance of invasion and metastatic phenotypes [
27]. Therefore, inhibition of uPA expression may contribute to a reduction in cancer metastasis. The data presented in
Figure 4 appeared to support the findings that the anti-cancer drug—cisplatin—attenuated uPA activity, resulting in a reduction in the expression of MMPs. Unexpectedly, the low-dose treatment with antrodan (20 mg/kg) only induced a higher level of uPA expression, than with the high-dose treatment (40 mg/kg). We supposed that the low-dose treatment with the polysaccharide might have stimulated macrophage activity, and increased the secretion of uPA. Clearly, more work is needed to resolve the molecular requirements and mechanisms for the inhibition of uPA with antrodan. However, the combined treatment of antrodan and cisplatin seemed to better improve the attenuation of uPA activity and the formation of MMPs than treatment with cisplatin only (
Figure 4).
IL-6 is an important cytokine that participates in the host’s inflammatory response. IL-6 activates many signaling molecules, including those in the pathways of PI3K, JAK-STAT, and MAPK, which affect the development of cancer. IL-6 promotes cancer cell proliferation and cancer cell metastasis via the PI3K and MAPK pathways. IL-6 also activates STAT-3 to promote VEGF protein expression, angiogenesis, and the growth of cancer cells. In clinical studies, the levels of IL-6 in plasma of cancer patients were positively correlated with the progression of cancer symptoms [
28]. Thus, IL-6 is one of the cytokines that has an important role in cancer pathogenesis. In this study, we found that the plasma levels of IL-6 in LLC-bearing mice were significantly reduced by treatment with antrodan only (20 mg/kg and 40 mg/kg), while the levels of IL-6 were not affected by treatment with cisplatin only. These results indicated that antrodan and cisplatin may have different mechanisms of action with respect to inhibiting tumor metastasis.
IFN-γ inhibits cancer cell growth by promoting innate immune responses, such as the macrophage-mediated killing of cancer cells, antigen presentation, and Th1-type cell activation [
29]. In the present study, low doses of antrodan (20 mg/kg) only did not induce a change in IFN-γ concentration in blood plasma of LLC-bearing mice, but higher doses (40 mg/kg) significantly increased plasma concentrations of IFN-γ, superior to those upon treatment with a combination of antrodan and cisplatin (
Table 2).
Members of the signal transducer and activator of transcription (STAT) protein family, known as latent cytoplasmic transcription factors, are involved in cell proliferation, differentiation, and survival. STAT3 is activated by many cytokines (e.g., IL-6), growth factors, and the Src and Ras oncoproteins. When STAT-3 is activated, the tumor cell micro-environment is promoted through cell proliferation, angiogenesis, metastasis, and immune evasion [
10]. Similarly, STAT-3 overexpression is associated with the metastasis of many types of cancer, including pancreatic cancer, colorectal carcinoma, gastric cancer, and lung cancer [
11]. In the present study, we found that treatments with both antrodan only and cisplatin only significantly inhibited STAT-3 phosphorylation in lung and liver tissues. Thus, the evidence suggested that the suppression of tumor metastasis by antrodan was related to the modulation of the IL-6/STAT-3 pathway.
The MAPK family can be divided into three different subclasses, namely extracellular signal-regulated kinases 1 and 2 (ERK-1/2), c-Jun N-terminal kinases (JNKs), and p38 [
30]. In cancer cells, ERK/MAPK is activated by the epidermal growth factor receptor, and Ras, leading to the promotion of proliferation, survival, and metastasis of cancer cells [
31]. Both JNK/MAPK and p38 MAPK are stress-activated protein kinases that are frequently activated by environmental and genotoxic stress, and inflammatory response factors [
30]. JNK/MAPK affects the survival, apoptosis, and metastasis of cancer cells through the regulation of transcription factor AP-1, and the related gene expression of the cell cycle, and through the secretion of MMPs and nuclear hormone receptors (e.g., the retinoic acid receptor). P38 MAPK regulates many physiological functions, including the cell cycle, inflammation, and cell differentiation. Activation of p38 MAPK usually inhibits cell proliferation, but improves proliferation, invasion, inflammation, and angiogenesis in cancer cells. This may be related to the activation of kinases, and the interactions between various pathways [
14,
32]. Studies have shown that bladder cancer invasion may be promoted by the p38 MAPK pathway through the regulation of the downstream MAPKAPK2 so as to regulate MMP-2 and MMP-9 activity [
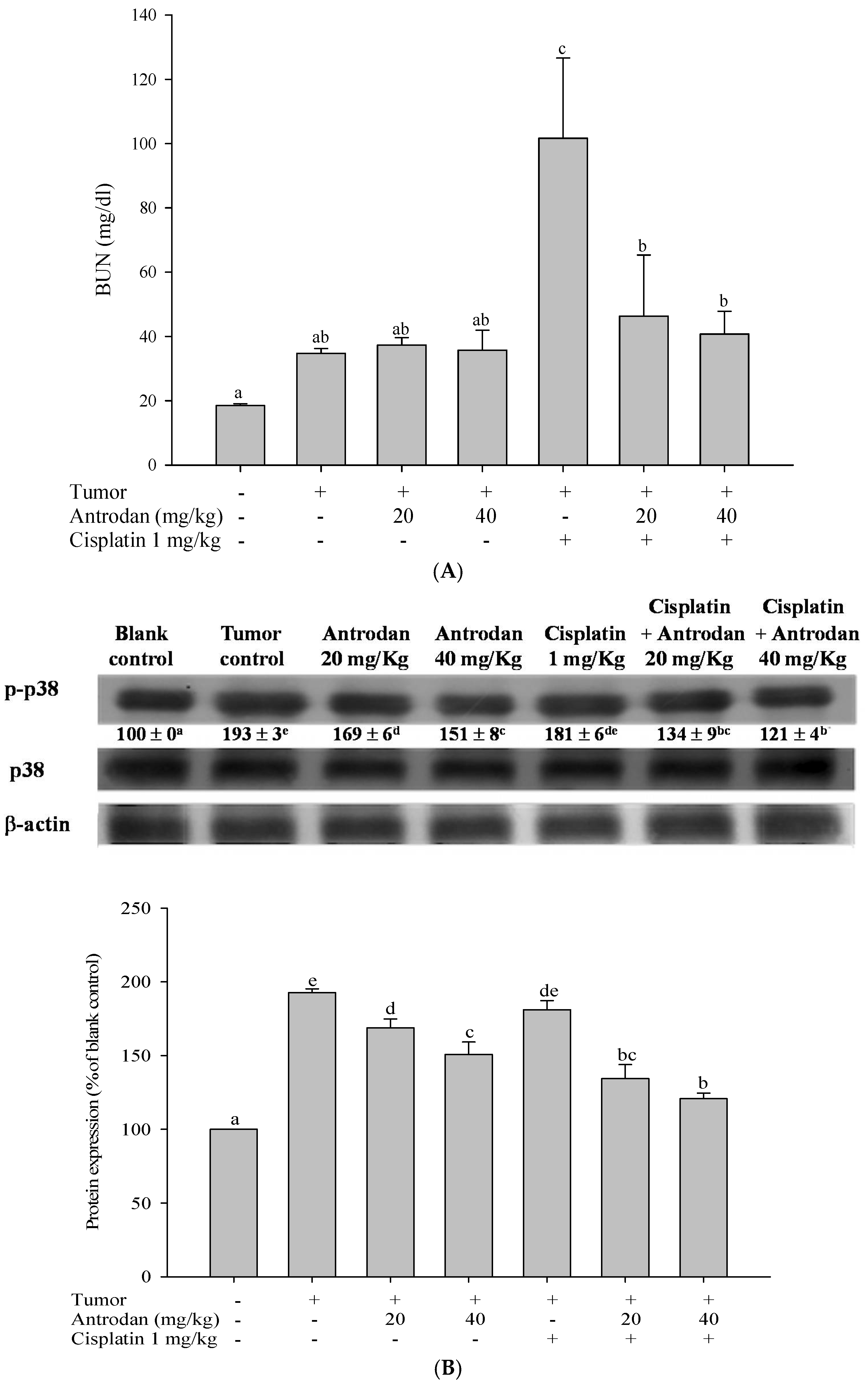
33]. Herein, we showed that both antrodan and cisplatin significantly inhibited tumor metastasis, and the phosphorylation of ERK1/2, JNK1/2, and p38 in lung and liver tissues. On the other hand, although cisplatin was found to have similar effects, this anti-cancer drug can cause pathological changes, such as renal cell apoptosis and tubular necrosis, leading to an increase in the levels of plasma BUN, and possible acute renal failure [
34]. Results of this study showed that the plasma levels of BUN in the tumor control group were significantly higher than those in the blank control group. It was speculated that the rise in levels of BUN and cachexia promotes alienation in tissue proteins [
35]. In addition, when compared with the tumor control group, treatment with cisplatin only significantly increased plasma BUN levels; however, this was not observed in treatments with antrodan only or a combination of both. The upregulated levels of BUN through treatment with cisplatin may be ameliorated through treatment with antrodan by reducing or ameliorating the levels of kidney dysfunction and cachexia. In addition, treatment with cisplatin may induce renal cell apoptosis, oxidative stress, and genotoxicity, resulting in kidney damage [
16]. The damage mechanism may be related to the promotion of cell death, caused by the activation of ERK, JNK, and p38 MAPK pathways in renal epithelial cells. Among them, p38 MAPK is involved in the production of TNF-α. TNF-α promotes renal oxidative stress, leading to nephrotoxicity. Therefore, inhibition of the phosphorylation of p38 MAPK may represent a potential strategy to alleviate renal dysfunction [
36]. When compared with the tumor control group, treatment with antrodan only (20 mg/kg and 40 mg/kg) significantly reduced p38 phosphorylation in the kidney. We hypothesized that the increase of p38 phosphorylation was due to tumor formation in the kidney, and that antrodan can ameliorate kidney damage by reducing the phosphorylation of p38 in the kidney. When compared with the tumor control group, treatment with cisplatin only had no significant effect on the phosphorylation of p38 in the kidney, while the combined treatment of antrodan and cisplatin significantly reduced cisplatin-induced expression of p38, suggesting that the combined treatment could reduce tumor-induced phosphorylation of p38. In this context, our presented data indicated that treatment with antrodan only alleviated cisplatin-induced kidney dysfunction through the p38 M APK pathway.
4. Materials and Methods
4.1. Chemicals and Antibodies
Dulbecco’s modified eagle medium (DMEM), non-essential amino acids (NEAAs), penicillin, sodium pyruvate, trypsin, and fetal bovine serum (FBS) were obtained from Gibco/BRL (Rockville, MD, USA). Antibodies of mitogen-activated protein kinases (MAPKs), matrix metalloproteinases (MMPs)-2 and -9 were obtained from Cell Signaling (Bevly, MA, USA). Antibodies of signal transducer and activator of transcription 3 (STAT-3), and secondary antibodies of anti-rabbit IgG were obtained from GeneTex (CA, USA). Antibodies of JNK1 (sc-136205), JNK2 (sc-271133), ERK1 (sc-376852), ERK2 (sc-271451), and p38 (sc-398305) were purchased from Santa Cruz (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Cisplatin (50 mg/100 mL) was obtained from Yung Shin (Yung Shin Pharm. Ind. Co., Ltd., Taichung, Taiwan).
4.2. Cell Lines
The murine Lewis lung carcinoma (LLC) cells (BCRC NO. 60050) were purchased from the Food Industry Research and Development Institute (FIRDI, Hsinchu, Taiwan), and were cultured in DMEM containing 10% (v/v) FBS, 0.37% (w/v) NaHCO3, and penicillin (100 unit/mL) in a 37 °C, 5% CO2, and 95% air incubator.
4.3. Source of Antrodan
Extraction and purification of antrodan from
Antrodia cinnamomea mycelia was performed as in our previous report [
24] with slight modifications. In brief, the freeze-dried and defatted mycelia were dissolved in water (1:10,
w/
v), and heated at 80°C for 2 h to remove the water-soluble materials. The residues were then extracted three times with a hot alkaline solution (pH 9.0, 1:10
w/
v) at 80 °C, each time for 2 h. The extracts were combined and filtered. The pH value of the filtrates was adjusted to 4.0 by using 1 M HCl solution, and they were then precipitated overnight at 4 °C. After the precipitates were collected by centrifuging at 3500
g for 30 min, they were dialyzed with deionized water (DDW) for three days to remove the free sugars and amino acids (dialysis tube Mw cut-off 12,000–16,000 Da, Wako, Tokyo, Japan), and then freeze-dried to yield base-soluble extracts. The extracts containing antrodan were then loaded onto a Sepharose CL-6B column (3.0 × 82 cm), and eluted with DDW at pH 11.0 (adjusted using 1 M NaOH) to separate polysaccharides at a flow rate of 0.5 mL/min, and to collect the target with a fraction collector. The product was obtained at about 10% yield with an average molecular weight of 442 kD by high-performance size-exclusion chromatography (HPSEC).
4.4. Tumor Xenografts and Antrodan/Cisplatin Treatments
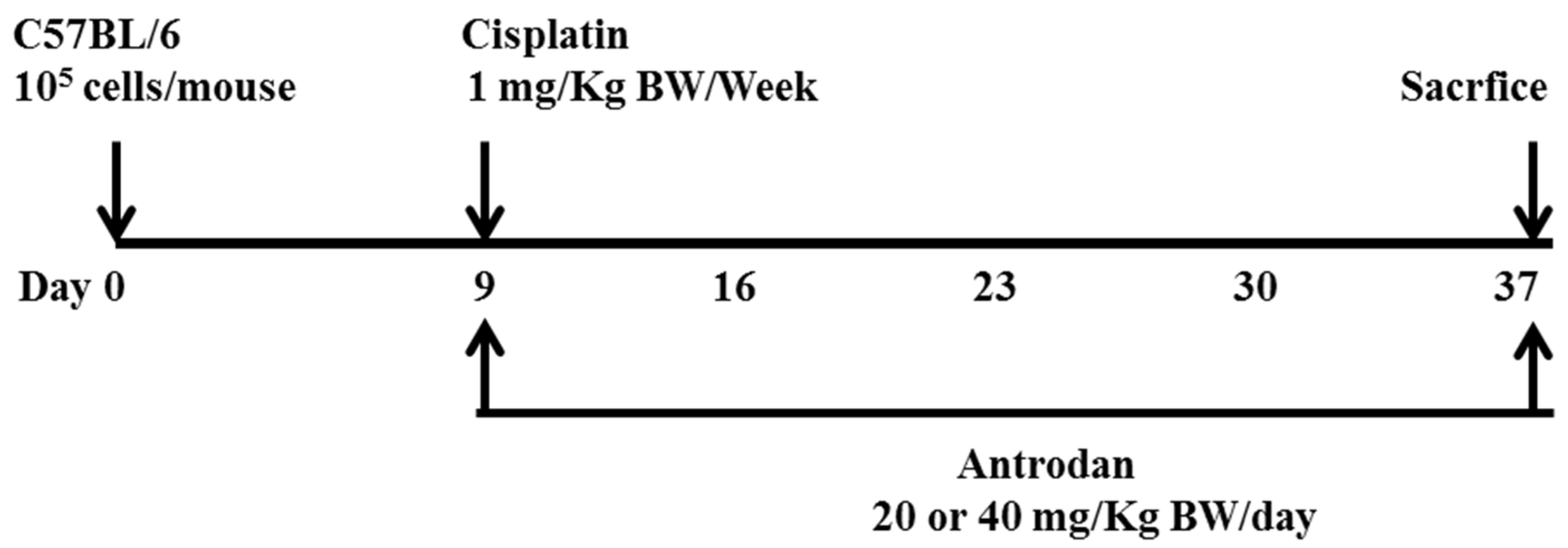
Forty-two C57BL/6 male mice (5-weeks-old) were provided by the BioLASCO Taiwan Co., Ltd. (Taipei, Taiwan). All experiments were carried out according to the Animal Research Committee of National Chung Hsing University, and this study was approved by the committee (No. 103-55). The mice were housed in a pathogen-free room at controlled temperature (25 ± 2 °C) and humidity (65 ± 5%), and alternating 12-h-light/-dark cycles. After the mice had been acclimated for one week, 100 µL of saline containing 1 × 10
5 LLC cells was subcutaneously injected into the right flanks of C57BL/6 mice. Nine days after implantation, the mice with successful tumor formations on the skin were randomly divided into seven groups (
n = 5–8 for each group) as follows: Group 1, blank control; Group 2, tumor control (implanted LLC cells); Group 3, tumor cell implantation, and oral administration of low-dose antrodan (20 mg/kg); Group 4, tumor cell implantation, and oral administration of high-dose antrodan (40 mg/kg); Group 5, tumor cell implantation, and i.p. administration of cisplatin (1 mg/kg); Group 6, tumor cell implantation, i.p. administration of cisplatin (1 mg/kg), and oral administration of antrodan (20 mg/kg); and Group 7, tumor cell implantation, i.p. administration of cisplatin (1 mg/kg), and oral administration of antrodan (40 mg/kg). The tumor areas were measured once every four days, and calculated as (π ÷ 4) × length (cm) × width (cm) using a vernier caliper [
25]. The standard diet (3.3 kcal/g) contained 58.9% carbohydrate, 28.7% protein, and 12.4% crude fat. Antrodan was dissolved in phosphate buffer saline (PBS), and was orally administered once per day, whereas cisplatin (1 mg/kg; i.p., 50 µL) was administered twice per week. For the continuous 28-day treatments, body weight was measured twice per week. The mice were then sacrificed, and the tumors were isolated. Lung, liver, spleen, and kidney samples were harvested for further examination. The number of lung and liver metastases was determined by counting the number of metastatic nodules on the lung and liver surfaces. The lungs and livers were then either stored in the freezer at −80 °C, or fixed with 10% formalin for further studies.
4.5. Determination of the Levels of Plasma Cytokines—IL-6 and IFN-γ
Cytokines—interleukin (IL)-6 and IFN-γ—were quantified using commercial kits with solid phase ELISA (R&D systems, Inc., Minneapolis, MN, USA) at a wavelength of 450 nm, performed according to the manufacturer’s protocols.
4.6. Determination of MMP-2, MMP-9, and uPA Levels in Plasma through Zymography
Our previous report was followed with respect to the collection and preparation of blood samples [
26]. The activities of MMP-2, MMP-9, and uPA in the plasma of mice were measured through gelatin-zymogram protease assays, as previously described [
24,
37]. In brief, prior to electrophoresis, the prepared plasma was diluted to 1/20 X with PBS and mixed with sodium dodecyl sulfate (SDS) sample loading buffer (5:1,
v/
v). Then, the prepared samples were electrophoresed (80 V, 2 h) using 8% sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE), containing 0.1% (
w/
v) gelatin, and 2% casein-plasminogen. After staining with Coomassie Brilliant Blue R-250, the relative activities of MMP-2, MMP-9, and uPA were quantified through densitometry using the AlphaEaseFC software (Alpha Innotech, Santa Clara, CA, USA).
4.7. Western Blotting
Protein expression levels of MMP-2, MMP-9, pSTAT3, STAT3, pERK1, pERK2, ERK1, ERK2, pJNK1, pJNK2, JNK1, JNK2, p-p38, and p38 in lung and liver tissues were measured through western blotting. The lung and liver tissues were homogenized in radio-immunoprecipitation assay (RIPA) buffer and protease inhibitors, and centrifuged (10,000 g, 5 min). The supernatants were frozen at −80 °C until ready for use. An amount of protein (40 µg) from the supernatant was mixed with 1/5 X Laemmli sample buffer containing 60 mM Tris-HCl pH 6.8, 25% glycerol, 2% SDS, 14.4 mM β-mercaptoethanol, and 0.1% bromophenol blue, before being denatured by heating to 95 °C for 5 min. Samples were then separated on a 10% SDS-polyacrylamide gel, and electroblotted to nitrocellulose membranes. After blocking with Tris-buffered saline (TBS) buffer (20 mM Tris–HCl, 150 mM NaCl, pH 7.4) containing 5% nonfat milk, the membrane was incubated overnight at 4 °C with the various primary antibodies, followed by horseradish peroxidase-conjugated anti-mouse IgG, and then visualized using an enhanced chemiluminescence ECL detection kit (PerkinElmer, Waltham, MA, USA), and quantified using the AlphaEaseFC software.
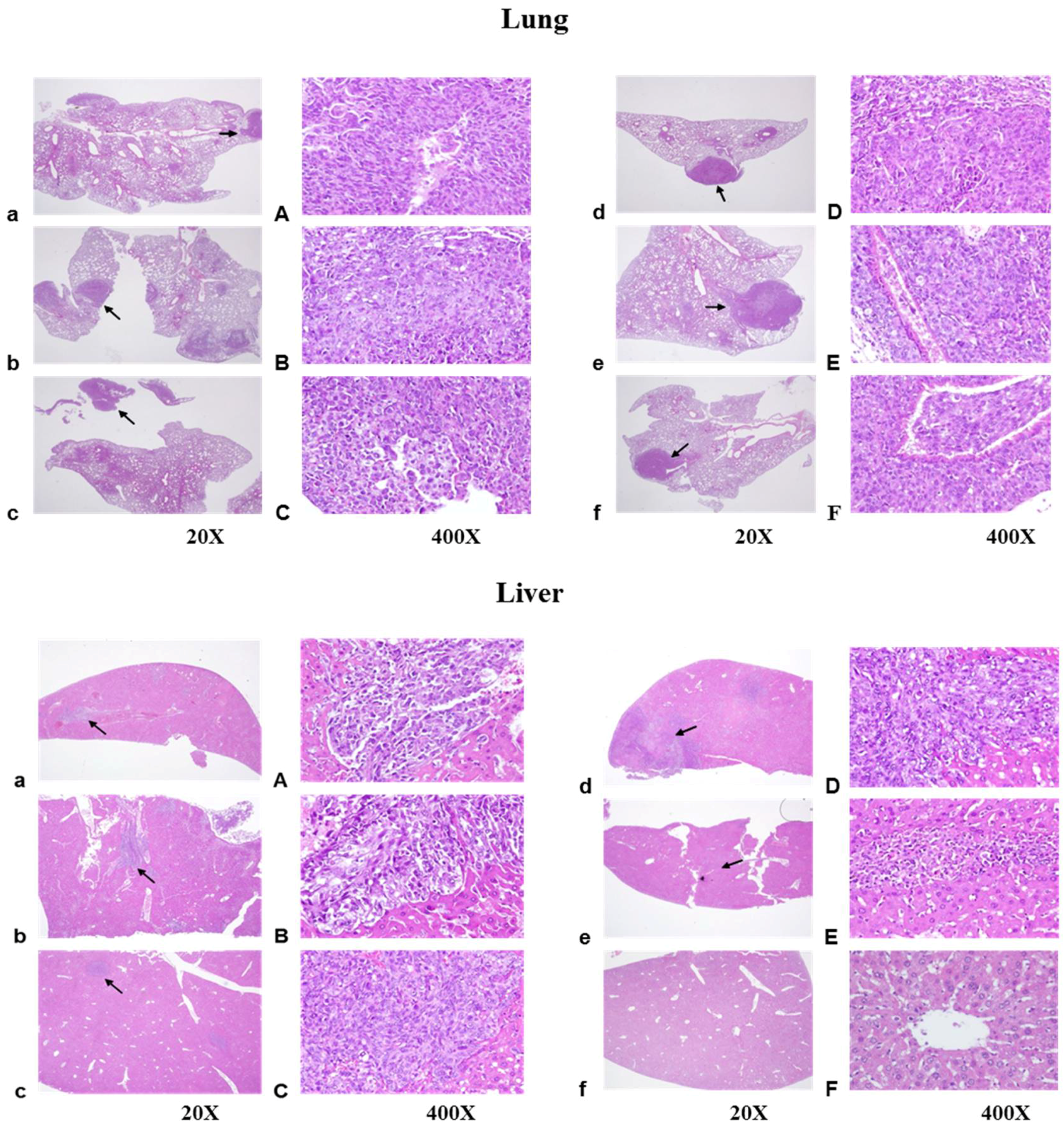
4.8. Histological Examination of Lung, Liver, and Kidney Tissues
Histological analysis of murine Lewis lung carcinomas in the lung, liver, or kidney of a C57BL/6 mouse was carried out using hematoxylin and eosin (H&E) staining. Tissues were formalin-fixed, embedded in paraffin, 2-μm-sectioned, and then subjected to H&E, and photographed.
4.9. Determination of Blood Urea Nitrogen (BUN) Levels
The blood urea nitrogen (BUN) levels were used for analysis of the renal function of mice, and were determined by FUJI DRI-CHEM 3500 (FUJI Technologies, Ebina, Japan). In brief, 20 µL of plasma was prepared as described above, then applied onto the kit, and detected at 625 nm. The BUN concentration was determined by the established calibration curve.
4.10. Statistical Analysis
Values were expressed as mean ± standard deviation (SD), and analyzed using one-way ANOVA followed by least significant difference (LSD) for the comparison of group means. All statistical analyses were performed using SPSS for Windows, version 10. Unless specified otherwise, a p-value < 0.05 was considered significant.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}