Updates on Aptamer Research


Abstract
1. Introduction
2. Antibodies and Aptamers
3. Super-Resolution Imaging
4. Aptamers for Super-Resolution Imaging
Funding
Conflicts of Interest
Abbreviations
| NAs | nucleic acids |
| SELEX | systematic evolution of ligands by exponential enrichment |
| STED | stimulated emission depletion microscopy |
| SMLM | single-molecule localization techniques |
| PET | positron-emission tomography |
| Cryo-EM | cryo-electron microscopy |
| STORM | stochastic optical reconstruction microscopy |
References
- Lakhin, A.; Tarantul, V.; Gening, L. Aptamers: Problems, solutions and prospects. Acta Nat. 2013, 5, 34–43. [Google Scholar]
- Saenger, W. Principles of Nucleic acid Structure; Springer Science & Business Media: Des Moines, IA, USA, 2013; pp. 1–8. [Google Scholar]
- Yarus, M. Getting past the rna world: The initial darwinian ancestor. Cold Spring Harb. Perspect. Biol. 2011, 3, a003590. [Google Scholar] [CrossRef] [PubMed]
- Robertson, M.P.; Joyce, G.F. The origins of the RNA world. Cold Spring Harb. Perspect. Biol. 2012, 4, a003608. [Google Scholar] [CrossRef]
- Higgs, P.G.; Lehman, N. The RNA world: Molecular cooperation at the origins of life. Nat. Rev. Genet. 2015, 16, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Blumenstiel, J.P. Evolutionary dynamics of transposable elements in a small RNA world. Trends Genet. 2011, 27, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, H.; Wrenger, C. Disease-specific biomarker discovery by aptamers. Cytom. Part A 2009, 75, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Tucker, W.O. Towards Specific DNA Aptamers which Bind and Inhibit wwp1 Hect Ubiquitin Ligase in the Osteoblast. Ph.D. Thesis, The University of Hong Kong, Hong Kong, China, June 2013. [Google Scholar]
- Klussmann, S. The Aptamer Handbook: Functional Oligonucleotides and Their Applications; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Veedu, R.N. Aptamers: Tools for Nanotherapy and Molecular Imaging; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Kaur, H.; Bruno, J.G.; Kumar, A.; Sharma, T.K. Aptamers in the therapeutics and diagnostics pipelines. Theranostics 2018, 8, 4016–4032. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Yung, L.Y. Probing high affinity sequences of DNA aptamer against vegf165. PLoS ONE 2012, 7, e31196. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage t4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Janjic, N.; Jarvis, T.; Schneider, D.; Walker, J.J.; Wilcox, S.K.; Zichi, D. Aptamers and the RNA world, past and present. Cold Spring Harb. Perspect. Biol. 2012, 4, a003582. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818. [Google Scholar] [CrossRef]
- Guo, K.-T.; Ziemer, G.; Paul, A.; Wendel, H.P. Cell-selex: Novel perspectives of aptamer-based therapeutics. Int. J. Mol. Sci. 2008, 9, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Qian, J.; Yang, Z.; Pang, Z.; Xi, Z.; Cao, S.; Wang, Y.; Pan, S.; Zhang, S.; Wang, W. Whole-cell selex aptamer-functionalised poly (ethyleneglycol)-poly (ε-caprolactone) nanoparticles for enhanced targeted glioblastoma therapy. Biomaterials 2012, 33, 6264–6272. [Google Scholar] [CrossRef]
- Chen, J.; Tong, T.; Wang, H. Super-resolution imaging in glycoscience: New developments and challenges. J. Innov. Opt. Health Sci. 2016, 09, 1630007. [Google Scholar] [CrossRef]
- Sefah, K.; Shangguan, D.; Xiong, X.; O’donoghue, M.B.; Tan, W. Development of DNA aptamers using cell-selex. Nat. Protoc. 2010, 5, 1169–1185. [Google Scholar] [CrossRef]
- Kaur, H. Recent developments in cell-selex technology for aptamer selection. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2323–2329. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. Selection in vitro of single-stranded DNA molecules that fold into specific ligand-binding structures. Nature 1992, 355, 850. [Google Scholar] [CrossRef] [PubMed]
- Berkhout, B.; Klaver, B. In vivo selection of randomly mutated retroviral genomes. Nucleic Acids Res. 1993, 21, 5020–5024. [Google Scholar] [CrossRef] [PubMed]
- Coulter, L.R.; Landree, M.A.; Cooper, T.A. Identification of a new class of exonic splicing enhancers by in vivo selection. Mol. Cell. Biol. 1997, 17, 2143–2150. [Google Scholar] [CrossRef] [PubMed]
- Jenison, R.D.; Gill, S.C.; Pardi, A.; Polisky, B. High-resolution molecular discrimination by RNA. Science 1994, 263, 1425–1429. [Google Scholar] [CrossRef] [PubMed]
- Dobbelstein, M.; Shenk, T. In vitro selection of RNA ligands for the ribosomal l22 protein associated with epstein-barr virus-expressed RNA by using randomized and cdna-derived RNA libraries. J. Virol. 1995, 69, 8027–8034. [Google Scholar]
- Smith, D.; Kirschenheuter, G.P.; Charlton, J.; Guidot, D.M.; Repine, J.E. In vitro selection of RNA-based irreversible inhibitors of human neutrophil elastase. Chem. Biol. 1995, 2, 741–750. [Google Scholar] [CrossRef]
- Jensen, K.B.; Atkinson, B.L.; Willis, M.C.; Koch, T.H.; Gold, L. Using in vitro selection to direct the covalent attachment of human immunodeficiency virus type 1 rev protein to high-affinity RNA ligands. Proc. Natl. Acad. Sci. USA 1995, 92, 12220–12224. [Google Scholar] [CrossRef]
- Golden, M.C.; Collins, B.D.; Willis, M.C.; Koch, T.H. Diagnostic potential of photoselex-evolved ssdna aptamers. J. Biotechnol. 2000, 81, 167–178. [Google Scholar] [CrossRef]
- Klußmann, S.; Nolte, A.; Bald, R.; Erdmann, V.A.; Fürste, J.P. Mirror-image RNA that binds d-adenosine. Nat. Biotechnol. 1996, 14, 1112. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.G. In vitroselection of DNA to chloroaromatics using magnetic microbead-based affinity separation and fluorescence detection. Biochem. Biophys. Res. Commun. 1997, 234, 117–120. [Google Scholar] [CrossRef]
- Tsai, R.Y.; Reed, R.R. Identification of DNA recognition sequences and protein interaction domains of the multiple-zn-finger protein roaz. Mol. Cell. Biol. 1998, 18, 6447–6456. [Google Scholar] [CrossRef]
- Vant-Hull, B.; Payano-Baez, A.; Davis, R.H.; Gold, L. The mathematics of selex against complex targets1. J. Mol. Biol. 1998, 278, 579–597. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.N.; Jensen, K.B.; Julin, C.M.; Weil, M.; Gold, L. High affinity ligands from in vitro selection: Complex targets. Proc. Natl. Acad. Sci. USA 1998, 95, 2902–2907. [Google Scholar] [CrossRef] [PubMed]
- Homann, M.; Göringer, H.U. Combinatorial selection of high affinity RNA ligands to live african trypanosomes. Nucleic Acids Res. 1999, 27, 2006–2014. [Google Scholar] [CrossRef]
- Burke, D.H.; Willis, J.H. Recombination, RNA evolution, and bifunctional RNA molecules isolated through chimeric selex. RNA 1998, 4, 1165–1175. [Google Scholar] [CrossRef]
- Wu, L.; Curran, J.F. An allosteric synthetic DNA. Nucleic Acids Res. 1999, 27, 1512–1516. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, J.; Imanaka, H.; Yokota, Y.; Sugimoto, N. In vitro selection of aptamers that act with Zn2+. J. Inorg. Biochem. 2000, 82, 197–206. [Google Scholar] [CrossRef]
- White, R.; Rusconi, C.; Scardino, E.; Wolberg, A.; Lawson, J.; Hoffman, M.; Sullenger, B. Generation of species cross-reactive aptamers using “toggle” selex. Mol. Ther. 2001, 4, 567–573. [Google Scholar] [CrossRef]
- Pagratis, N.; Gold, L.; Shtatland, T.; Javornik, B. Truncation selex method. U.S. Patent 6,261,774, 17 July 2001. [Google Scholar]
- Wen, J.-D.; Gray, D.M. Selection of genomic sequences that bind tightly to ff gene 5 protein: Primer-free genomic selex. Nucleic Acids Res. 2004, 32, e182. [Google Scholar] [CrossRef]
- Pan, W.; Xin, P.; Clawson, G.A. Minimal primer and primer-free selex protocols for selection of aptamers from random DNA libraries. BioTechniques 2008, 44, 351–362. [Google Scholar] [CrossRef]
- Pan, W.; Clawson, G.A. Primer-free aptamer selection using a random DNA library. In RNA Therapeutics; Springer: Berlin/Heidelberg, Germany, 2010; pp. 367–383. [Google Scholar]
- Martell, R.E.; Nevins, J.R.; Sullenger, B.A. Optimizing aptamer activity for gene therapy applications using expression cassette selex. Mol. Ther. 2002, 6, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Berezovski, M.; Krylov, S.N. Nonequilibrium capillary electrophoresis of equilibrium mixtures− a single experiment reveals equilibrium and kinetic parameters of protein− DNA interactions. J. Am. Chem. Soc. 2002, 124, 13674–13675. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Yang, G.; Zhang, D.; Ding, H.; Wang, H.; Fan, M.; Shen, B.; Shao, N. Single-stranded DNA aptamers that bind differentiated but not parental cells: Subtractive systematic evolution of ligands by exponential enrichment. J. Biotechnol. 2003, 102, 15–22. [Google Scholar] [CrossRef]
- Vater, A.; Jarosch, F.; Buchner, K.; Klussmann, S. Short bioactive spiegelmers to migraine-associated calcitonin gene-related peptide rapidly identified by a novel approach: Tailored-selex. Nucleic Acids Res. 2003, 31, e130. [Google Scholar] [CrossRef] [PubMed]
- Mendonsa, S.D.; Bowser, M.T. In vitro evolution of functional DNA using capillary electrophoresis. J. Am. Chem. Soc. 2004, 126, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Okhonin, V.; Berezovski, M.; Krylov, S.N. Sweeping capillary electrophoresis: A non-stopped-flow method for measuring bimolecular rate constant of complex formation between protein and DNA. J. Am. Chem. Soc. 2004, 126, 7166–7167. [Google Scholar] [CrossRef] [PubMed]
- Asai, R.; Nishimura, S.I.; Aita, T.; Takahashi, K. In vitro selection of DNA aptamers on chips using a method for generating point mutations. Anal. Lett. 2004, 37, 645–656. [Google Scholar] [CrossRef]
- Drabovich, A.; Berezovski, M.; Krylov, S.N. Selection of smart aptamers by equilibrium capillary electrophoresis of equilibrium mixtures (eceem). J. Am. Chem. Soc. 2005, 127, 11224–11225. [Google Scholar] [CrossRef] [PubMed]
- Stoltenburg, R.; Reinemann, C.; Strehlitz, B. Flumag-selex as an advantageous method for DNA aptamer selection. Anal. Bioanal. Chem. 2005, 383, 83–91. [Google Scholar] [CrossRef]
- Berezovski, M.V.; Musheev, M.U.; Drabovich, A.P.; Jitkova, J.V.; Krylov, S.N. Non-selex: Selection of aptamers without intermediate amplification of candidate oligonucleotides. Nat. Protoc. 2006, 1, 1359. [Google Scholar] [CrossRef] [PubMed]
- Win, M.N.; Klein, J.S.; Smolke, C.D. Codeine-binding RNA aptamers and rapid determination of their binding constants using a direct coupling surface plasmon resonance assay. Nucleic Acids Res 2006, 34, 5670–5682. [Google Scholar] [CrossRef]
- Khati, M.; Schüman, M.; Ibrahim, J.; Sattentau, Q.; Gordon, S.; James, W. Neutralization of infectivity of diverse r5 clinical isolates of human immunodeficiency virus type 1 by gp120-binding 2′ f-RNA aptamers. J. Virol. 2003, 77, 12692–12698. [Google Scholar] [CrossRef] [PubMed]
- Ohuchi, S.P.; Ohtsu, T.; Nakamura, Y. Selection of RNA aptamers against recombinant transforming growth factor-β type iii receptor displayed on cell surface. Biochimie 2006, 88, 897–904. [Google Scholar] [CrossRef]
- Lorenz, C.; Gesell, T.; Zimmermann, B.; Schoeberl, U.; Bilusic, I.; Rajkowitsch, L.; Waldsich, C.; Von Haeseler, A.; Schroeder, R. Genomic selex for hfq-binding RNAs identifies genomic aptamers predominantly in antisense transcripts. Nucleic Acids Res. 2010, 38, 3794–3808. [Google Scholar] [CrossRef]
- Fujimoto, Y.; Nakamura, Y.; Ohuchi, S. Hexim1-binding elements on mRNAs identified through transcriptomic selex and computational screening. Biochimie 2012, 94, 1900–1909. [Google Scholar] [CrossRef] [PubMed]
- Layzer, J.M.; Sullenger, B.A. Simultaneous generation of aptamers to multiple gamma-carboxyglutamic acid proteins from a focused aptamer library using deselex and convergent selection. Oligonucleotides 2007, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, A.; Kurth, A.; Dunkhorst, A.; Pänke, O.; Sielaff, H.; Junge, W.; Muth, D.; Scheller, F.; Stöcklein, W.; Dahmen, C. One-step selection of vaccinia virus-binding DNA aptamers by monolex. BMC Biotechnol. 2007, 7, 48. [Google Scholar] [CrossRef]
- Peng, L.; Stephens, B.J.; Bonin, K.; Cubicciotti, R.; Guthold, M. A combined atomic force/fluorescence microscopy technique to select aptamers in a single cycle from a small pool of random oligonucleotides. Microsc. Res. Tech. 2007, 70, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; McBurnett, S.R.; Andrews, C.J.; Allman, A.M.; Bruno, J.G.; Kiel, J.L. Aptamer selection express: A novel method for rapid single-step selection and sensing of aptamers. J. Biomol. Tech. JBT 2008, 19, 311. [Google Scholar]
- Raddatz, M.S.L.; Dolf, A.; Endl, E.; Knolle, P.; Famulok, M.; Mayer, G. Enrichment of cell-targeting and population-specific aptamers by fluorescence-activated cell sorting. Angew. Chem. 2008, 120, 5268–5271. [Google Scholar] [CrossRef]
- Mayer, G.; Ahmed, M.-S.L.; Dolf, A.; Endl, E.; Knolle, P.A.; Famulok, M. Fluorescence-activated cell sorting for aptamer selex with cell mixtures. Nat. Protoc. 2010, 5, 1993. [Google Scholar] [CrossRef]
- Tok, J.B.-H.; Fischer, N.O. Single microbead selex for efficient ssdna aptamer generation against botulinum neurotoxin. Chem. Commun. 2008, 1883–1885. [Google Scholar] [CrossRef]
- Knight, C.G.; Platt, M.; Rowe, W.; Wedge, D.C.; Khan, F.; Day, P.J.; McShea, A.; Knowles, J.; Kell, D.B. Array-based evolution of DNA aptamers allows modelling of an explicit sequence-fitness landscape. Nucleic Acids Res. 2008, 37, e6. [Google Scholar] [CrossRef]
- Chushak, Y.; Stone, M.O. In silico selection of RNA aptamers. Nucleic Acids Res. 2009, 37, e87. [Google Scholar] [CrossRef]
- Li, S.; Xu, H.; Ding, H.; Huang, Y.; Cao, X.; Yang, G.; Li, J.; Xie, Z.; Meng, Y.; Li, X. Identification of an aptamer targeting hnrnp a1 by tissue slide-based selex. J. Pathol. 2009, 218, 327–336. [Google Scholar] [CrossRef]
- Park, S.-m.; Ahn, J.-Y.; Jo, M.; Lee, D.-k.; Lis, J.T.; Craighead, H.G.; Kim, S. Selection and elution of aptamers using nanoporous sol-gel arrays with integrated microheaters. Lab Chip 2009, 9, 1206–1212. [Google Scholar] [CrossRef]
- Zykovich, A.; Korf, I.; Segal, D.J. Bind-n-seq: High-throughput analysis of in vitro protein–DNA interactions using massively parallel sequencing. Nucleic Acids Res. 2009, 37, e151. [Google Scholar] [CrossRef]
- Jolma, A.; Kivioja, T.; Toivonen, J.; Cheng, L.; Wei, G.; Enge, M.; Taipale, M.; Vaquerizas, J.M.; Yan, J.; Sillanpää, M.J. Multiplexed massively parallel selex for characterization of human transcription factor binding specificities. Genome Res. 2010, 20, 861–873. [Google Scholar] [CrossRef]
- Slattery, M.; Riley, T.; Liu, P.; Abe, N.; Gomez-Alcala, P.; Dror, I.; Zhou, T.; Rohs, R.; Honig, B.; Bussemaker, H.J. Cofactor binding evokes latent differences in DNA binding specificity between hox proteins. Cell 2011, 147, 1270–1282. [Google Scholar] [CrossRef]
- Lou, X.; Qian, J.; Xiao, Y.; Viel, L.; Gerdon, A.E.; Lagally, E.T.; Atzberger, P.; Tarasow, T.M.; Heeger, A.J.; Soh, H.T. Micromagnetic selection of aptamers in microfluidic channels. Proc. Natl. Acad. Sci. USA 2009, 106, 2989–2994. [Google Scholar] [CrossRef]
- Qian, J.; Lou, X.; Zhang, Y.; Xiao, Y.; Soh, H.T. Generation of highly specific aptamers via micromagnetic selection. Anal. Chem. 2009, 81, 5490–5495. [Google Scholar] [CrossRef]
- Oh, S.S.; Qian, J.; Lou, X.; Zhang, Y.; Xiao, Y.; Soh, H.T. Generation of highly specific aptamers via micromagnetic selection. Anal. Chem. 2011, 83, 1866. [Google Scholar] [CrossRef]
- Huang, C.-J.; Lin, H.-I.; Shiesh, S.-C.; Lee, G.-B. Integrated microfluidic system for rapid screening of crp aptamers utilizing systematic evolution of ligands by exponential enrichment (selex). Biosens. Bioelectron. 2010, 25, 1761–1766. [Google Scholar] [CrossRef]
- Cho, M.; Xiao, Y.; Nie, J.; Stewart, R.; Csordas, A.T.; Oh, S.S.; Thomson, J.A.; Soh, H.T. Quantitative selection of DNA aptamers through microfluidic selection and high-throughput sequencing. Proc. Natl. Acad. Sci. USA 2010, 107, 15373–15378. [Google Scholar] [CrossRef]
- Mi, J.; Liu, Y.; Rabbani, Z.N.; Yang, Z.; Urban, J.H.; Sullenger, B.A.; Clary, B.M. In vivo selection of tumor-targeting RNA motifs. Nat. Chem. Biol. 2010, 6, 22. [Google Scholar] [CrossRef]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 5, e15004. [Google Scholar] [CrossRef] [PubMed]
- Savory, N.; Abe, K.; Sode, K.; Ikebukuro, K. Selection of DNA aptamer against prostate specific antigen using a genetic algorithm and application to sensing. Biosens. Bioelectron. 2010, 26, 1386–1391. [Google Scholar] [CrossRef]
- Nonaka, Y.; Yoshida, W.; Abe, K.; Ferri, S.; Schulze, H.; Bachmann, T.T.; Ikebukuro, K. Affinity improvement of a vegf aptamer by in silico maturation for a sensitive vegf-detection system. Anal. Chem. 2012, 85, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Savory, N.; Lednor, D.; Tsukakoshi, K.; Abe, K.; Yoshida, W.; Ferri, S.; Jones, B.V.; Ikebukuro, K. In silico maturation of binding-specificity of DNA aptamers against proteus mirabilis. Biotechnol. Bioeng. 2013, 110, 2573–2580. [Google Scholar] [CrossRef]
- Savory, N.; Takahashi, Y.; Tsukakoshi, K.; Hasegawa, H.; Takase, M.; Abe, K.; Yoshida, W.; Ferri, S.; Kumazawa, S.; Sode, K. Simultaneous improvement of specificity and affinity of aptamers against streptococcus mutans by in silico maturation for biosensor development. Biotechnol. Bioeng. 2014, 111, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Jing, M.; Bowser, M.T. Isolation of DNA aptamers using micro free flow electrophoresis. Lab Chip 2011, 11, 3703–3709. [Google Scholar] [CrossRef]
- Hoon, S.; Zhou, B.; Janda, K.D.; Brenner, S.; Scolnick, J. Aptamer selection by high-throughput sequencing and informatic analysis. Biotechniques 2011, 51, 413–416. [Google Scholar] [CrossRef]
- Lauridsen, L.H.; Shamaileh, H.A.; Edwards, S.L.; Taran, E.; Veedu, R.N. Rapid one-step selection method for generating nucleic acid aptamers: Development of a DNA aptamer against α-bungarotoxin. PLoS ONE 2012, 7, e41702. [Google Scholar] [CrossRef]
- Thiel, W.H.; Bair, T.; Peek, A.S.; Liu, X.; Dassie, J.; Stockdale, K.R.; Behlke, M.A.; Miller Jr, F.J.; Giangrande, P.H. Rapid identification of cell-specific, internalizing RNA aptamers with bioinformatics analyses of a cell-based aptamer selection. PLoS ONE 2012, 7, e43836. [Google Scholar] [CrossRef]
- Stoltenburg, R.; Nikolaus, N.; Strehlitz, B. Capture-selex: Selection of DNA aptamers for aminoglycoside antibiotics. J. Anal. Methods Chem. 2012, 2012, 415697. [Google Scholar] [CrossRef] [PubMed]
- Waybrant, B.; Pearce, T.R.; Wang, P.; Sreevatsan, S.; Kokkoli, E. Development and characterization of an aptamer binding ligand of fractalkine using domain targeted selex. Chem. Commun. 2012, 48, 10043–10045. [Google Scholar] [CrossRef] [PubMed]
- WooáKim, D.; BockáGu, M. Immobilization-free screening of aptamers assisted by graphene oxide. Chem. Commun. 2012, 48, 2071–2073. [Google Scholar]
- Gong, Q.; Wang, J.; Ahmad, K.M.; Csordas, A.T.; Zhou, J.; Nie, J.; Stewart, R.; Thomson, J.A.; Rossi, J.J.; Soh, H.T. Selection strategy to generate aptamer pairs that bind to distinct sites on protein targets. Anal. Chem. 2012, 84, 5365–5371. [Google Scholar] [CrossRef]
- Szeto, K.; Latulippe, D.R.; Ozer, A.; Pagano, J.M.; White, B.S.; Shalloway, D.; Lis, J.T.; Craighead, H.G. Rapid-selex for RNA aptamers. PLoS ONE 2013, 8, e82667. [Google Scholar] [CrossRef]
- Kimoto, M.; Yamashige, R.; Matsunaga, K.-i.; Yokoyama, S.; Hirao, I. Generation of high-affinity DNA aptamers using an expanded genetic alphabet. Nat. Biotechnol. 2013, 31, 453. [Google Scholar] [CrossRef] [PubMed]
- Sefah, K.; Yang, Z.; Bradley, K.M.; Hoshika, S.; Jiménez, E.; Zhang, L.; Zhu, G.; Shanker, S.; Yu, F.; Turek, D. In vitro selection with artificial expanded genetic information systems. Proc. Natl. Acad. Sci. USA 2014, 111, 1449–1454. [Google Scholar] [CrossRef]
- Lao, Y.-H.; Chiang, H.-Y.; Yang, D.-K.; Peck, K.; Chen, L.-C. Selection of aptamers targeting the sialic acid receptor of hemagglutinin by epitope-specific selex. Chem. Commun. 2014, 50, 8719–8722. [Google Scholar] [CrossRef]
- Lai, J.-C.; Hong, C.-Y. Magnetic-assisted rapid aptamer selection (maras) for generating high-affinity DNA aptamer using rotating magnetic fields. ACS Comb. Sci. 2014, 16, 321–327. [Google Scholar] [CrossRef]
- Wang, J.; Gong, Q.; Maheshwari, N.; Eisenstein, M.; Arcila, M.L.; Kosik, K.S.; Soh, H.T. Particle display: A quantitative screening method for generating high-affinity aptamers. Angew. Chem. Int. Ed. 2014, 53, 4796–4801. [Google Scholar] [CrossRef]
- Zhu, Z.; Song, Y.; Li, C.; Zou, Y.; Zhu, L.; An, Y.; Yang, C.J. Monoclonal surface display selex for simple, rapid, efficient, and cost-effective aptamer enrichment and identification. Anal. Chem. 2014, 86, 5881–5888. [Google Scholar] [CrossRef]
- Jacoby, K.; Scharenberg, A.M. Homing endonuclease target determination using selex adapted for yeast surface display. In Homing Endonucleases; Springer: Berlin/Heidelberg, Germany, 2014; pp. 165–190. [Google Scholar]
- Ouellet, E.; Foley, J.H.; Conway, E.M.; Haynes, C. Hi-fi selex: A high-fidelity digital-PCR based therapeutic aptamer discovery platform. Biotechnol. Bioeng. 2015, 112, 1506–1522. [Google Scholar] [CrossRef] [PubMed]
- Mayer, G.; Tolle, F. Method of identifying or producing an aptamer. Google Patents WO2015049356A1, 19 October 2017. [Google Scholar]
- Tolle, F.; Brändle, G.M.; Matzner, D.; Mayer, G. A versatile approach towards nucleobase-modified aptamers. Angew. Chem. Int. Ed. 2015, 54, 10971–10974. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Sakota, E.; Nakamura, Y. The efficient cell-selex strategy, icell-selex, using isogenic cell lines for selection and counter-selection to generate RNA aptamers to cell surface proteins. Biochimie 2016, 131, 77–84. [Google Scholar] [CrossRef]
- Zumrut, H.E.; Ara, M.N.; Fraile, M.; Maio, G.; Mallikaratchy, P. Ligand-guided selection of target-specific aptamers: A screening technology for identifying specific aptamers against cell-surface proteins. Nucleic Acid Ther. 2016, 26, 190–198. [Google Scholar] [CrossRef]
- Yu, H.; Yang, W.; Alkhamis, O.; Canoura, J.; Yang, K.; Xiao, Y. In vitro isolation of small-molecule-binding aptamers with intrinsic dye-displacement functionality. Nucleic Acids Res. 2018. [Google Scholar] [CrossRef]
- Damase, T.R.; Miura, T.A.; Parent, C.E.; Allen, P.B. Application of the open qPCR instrument for the in vitro selection of DNA aptamers against epidermal growth factor receptor and drosophila c virus. ACS Comb. Sci. 2018. [Google Scholar] [CrossRef]
- Kulbachinskiy, A. Methods for selection of aptamers to protein targets. Biochemistry 2007, 72, 1505–1518. [Google Scholar] [CrossRef]
- Greenleaf, W.J.; Frieda, K.L.; Foster, D.A.; Woodside, M.T.; Block, S.M. Direct observation of hierarchical folding in single riboswitch aptamers. Science 2008, 319, 630–633. [Google Scholar] [CrossRef]
- Gu, M.B.; Kim, H.-S. Biosensors Based on Aptamers and Enzymes; Springer: Berlin/Heidelberg, Germany, 2014; Volume 140. [Google Scholar]
- Huang, B.; Wang, W.; Bates, M.; Zhuang, X. Three-dimensional super-resolution imaging by stochastic optical reconstruction microscopy. Science 2008, 319, 810–813. [Google Scholar] [CrossRef]
- Lippincott-Schwartz, J.; Patterson, G.H. Photoactivatable fluorescent proteins for diffraction-limited and super-resolution imaging. Trends Cell Biol. 2009, 19, 555–565. [Google Scholar] [CrossRef]
- Senoner, M.; Unger, W.E. Sims imaging of the nanoworld: Applications in science and technology. J. Anal. At. Spectrom. 2012, 27, 1050–1068. [Google Scholar] [CrossRef]
- Newman, C.F.; Havelund, R.; Passarelli, M.K.; Marshall, P.S.; Francis, I.; West, A.; Alexander, M.R.; Gilmore, I.S.; Dollery, C.T. Intracellular drug uptake a comparison of single cell measurements using tof-sims imaging and quantification from cell populations with lc/ms/ms. Anal. Chem. 2017, 89, 11944–11953. [Google Scholar] [CrossRef]
- Huang, B.; Babcock, H.; Zhuang, X. Breaking the diffraction barrier: Super-resolution imaging of cells. Cell 2010, 143, 1047–1058. [Google Scholar] [CrossRef]
- Mairal, T.; Özalp, V.C.; Sánchez, P.L.; Mir, M.; Katakis, I.; O’Sullivan, C.K. Aptamers: Molecular tools for analytical applications. Anal. Bioanal. Chem. 2008, 390, 989–1007. [Google Scholar] [CrossRef] [PubMed]
- Toh, S.Y.; Citartan, M.; Gopinath, S.C.; Tang, T.-H. Aptamers as a replacement for antibodies in enzyme-linked immunosorbent assay. Biosens. Bioelectron. 2015, 64, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Yang, S. Replacing antibodies with aptamers in lateral flow immunoassay. Biosens. Bioelectron. 2015, 71, 230–242. [Google Scholar] [CrossRef]
- Tan, W.; Wang, H.; Chen, Y.; Zhang, X.; Zhu, H.; Yang, C.; Yang, R.; Liu, C. Molecular aptamers for drug delivery. Trends Biotechnol. 2011, 29, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, J.; Nilsen-Hamilton, M. Aptamers: Multifunctional molecules for biomedical research. J. Mol. Med. 2013, 91, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Stoltenburg, R.; Reinemann, C.; Strehlitz, B. Selex—a (r) evolutionary method to generate high-affinity nucleic acid ligands. Biomol. Eng. 2007, 24, 381–403. [Google Scholar] [CrossRef]
- Fang, X.; Tan, W. Aptamers generated from cell-selex for molecular medicine: A chemical biology approach. Acc. Chem. Res. 2009, 43, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, V.J.; Levisson, M.; Eppink, M.H.; Smidt, H.; Van Der Oost, J. Alternative affinity tools: More attractive than antibodies? Biochem. J. 2011, 436, 1–13. [Google Scholar] [CrossRef]
- Lee, J.-O.; So, H.-M.; Jeon, E.-K.; Chang, H.; Won, K.; Kim, Y.H. Aptamers as molecular recognition elements for electrical nanobiosensors. Anal. Bioanal. Chem. 2008, 390, 1023–1032. [Google Scholar] [CrossRef]
- Kedzierski, S.; Khoshnejad, M.; Caltagirone, G.T. Synthetic antibodies: The emerging field of aptamers. Bioprocess. J. 2012, 11, 46–49. [Google Scholar] [CrossRef]
- McKeague, M.; DeRosa, M.C. Challenges and opportunities for small molecule aptamer development. J. Nucleic Acids 2012, 2012. [Google Scholar] [CrossRef]
- Que, E.L.; Domaille, D.W.; Chang, C.J. Metals in neurobiology: Probing their chemistry and biology with molecular imaging. Chem. Rev. 2008, 108, 1517–1549. [Google Scholar] [CrossRef] [PubMed]
- Keiser, M.J.; Setola, V.; Irwin, J.J.; Laggner, C.; Abbas, A.I.; Hufeisen, S.J.; Jensen, N.H.; Kuijer, M.B.; Matos, R.C.; Tran, T.B. Predicting new molecular targets for known drugs. Nature 2009, 462, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Berezovski, M.V.; Lechmann, M.; Musheev, M.U.; Mak, T.W.; Krylov, S.N. Aptamer-facilitated biomarker discovery (aptabid). J. Am. Chem. Soc. 2008, 130, 9137–9143. [Google Scholar] [CrossRef]
- Kalani, M.Y.S.; Theodore, N. Personalized medicine and neurosurgery. Front. Surg. 2017, 4, 19. [Google Scholar] [CrossRef]
- De Castro, M.A.G.; Höbartner, C.; Opazo, F. Aptamers provide superior stainings of cellular receptors studied under super-resolution microscopy. PLoS ONE 2017, 12, e0173050. [Google Scholar] [CrossRef]
- de Castro, M.A.; Rammner, B.; Opazo, F. Aptamer stainings for super-resolution microscopy. Methods Mol. Biol. 2016, 1380, 197–210. [Google Scholar]
- Smolyaninov, I.I.; Hung, Y.-J.; Davis, C.C. Magnifying superlens in the visible frequency range. Science 2007, 315, 1699–1701. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Babcock, H.P.; Zhuang, X. Dual-objective storm reveals three-dimensional filament organization in the actin cytoskeleton. Nat. Methods 2012, 9, 185–188. [Google Scholar] [CrossRef]
- Uhlén, M. Affinity as a tool in life science. Biotechniques 2008, 44, 649. [Google Scholar] [CrossRef] [PubMed]
- Ries, J.; Kaplan, C.; Platonova, E.; Eghlidi, H.; Ewers, H. A simple, versatile method for gfp-based super-resolution microscopy via nanobodies. Nat. Methods 2012, 9, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Mikhaylova, M.; Cloin, B.M.; Finan, K.; Van Den Berg, R.; Teeuw, J.; Kijanka, M.M.; Sokolowski, M.; Katrukha, E.A.; Maidorn, M.; Opazo, F. Resolving bundled microtubules using anti-tubulin nanobodies. Nat. Commun. 2015, 6, 7933. [Google Scholar] [CrossRef] [PubMed]
- Opazo, F.; Levy, M.; Byrom, M.; Schäfer, C.; Geisler, C.; Groemer, T.W.; Ellington, A.D.; Rizzoli, S.O. Aptamers as potential tools for super-resolution microscopy. Nat. Methods 2012, 9, 938. [Google Scholar] [CrossRef]
- Ylera, F.; Lurz, R.; Erdmann, V.A.; Furste, J.P. Selection of RNA aptamers to the alzheimer’s disease amyloid peptide. Biochem. Biophys. Res. Commun. 2002, 290, 1583–1588. [Google Scholar] [CrossRef]
- Rahimi, F.; Murakami, K.; Summers, J.L.; Chen, C.H.; Bitan, G. RNA aptamers generated against oligomeric abeta40 recognize common amyloid aptatopes with low specificity but high sensitivity. PLoS ONE 2009, 4, e7694. [Google Scholar] [CrossRef]
- Janas, T.; Sapon, K.; Stowell, M.H.B.; Janas, T. Selection of membrane RNA aptamers to amyloid beta peptide: Implications for exosome-based antioxidant strategies. Int. J. Mol. Sci. 2019, 20, 299. [Google Scholar] [CrossRef]
- Mahmoud, M.; Deigner, H.-P. Aptamers in diagnostics: Replacing or complementing antibodies? J. Biochips Tissue Chips 2015, 5, 1. [Google Scholar] [CrossRef]
- Kanwar, J.R.; Roy, K.; Maremanda, N.G.; Subramanian, K.; Veedu, R.N.; Bawa, R.; Kanwar, R.K. Nucleic acid-based aptamers: Applications, development and clinical trials. Curr. Med. Chem. 2015, 22, 2539–2557. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Leslie, M.; Kameyama, H.; Volk, D.E.; Tanaka, T. Aptamer therapeutics in cancer: Current and future. Cancers 2018, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.L.; Nascimento, I.C.; Santos, A.P.; Ogusuku, I.E.Y.; Lameu, C.; Mayer, G.; Ulrich, H. Aptamers: Novelty tools for cancer biology. Oncotarget 2018, 9, 26934–26953. [Google Scholar] [CrossRef] [PubMed]
- Hori, S.I.; Herrera, A.; Rossi, J.J.; Zhou, J. Current advances in aptamers for cancer diagnosis and therapy. Cancers 2018, 10, 9. [Google Scholar] [CrossRef]
- Hell, S.W. Microscopy and its focal switch. Nat. Methods 2009, 6, 24–32. [Google Scholar] [CrossRef]
- Harke, B.; Keller, J.; Ullal, C.K.; Westphal, V.; Schönle, A.; Hell, S.W. Resolution scaling in sted microscopy. Opt. Express 2008, 16, 4154–4162. [Google Scholar] [CrossRef]
- Chen, W.; Delaloye, S.; Silverman, D.H.; Geist, C.; Czernin, J.; Sayre, J.; Satyamurthy, N.; Pope, W.; Lai, A.; Phelps, M.E. Predicting treatment response of malignant gliomas to bevacizumab and irinotecan by imaging proliferation with [18f] fluorothymidine positron emission tomography: A pilot study. J. Clin. Oncol. 2007, 25, 4714–4721. [Google Scholar] [CrossRef]
- Sartori, A.; Gatz, R.; Beck, F.; Rigort, A.; Baumeister, W.; Plitzko, J.M. Correlative microscopy: Bridging the gap between fluorescence light microscopy and cryo-electron tomography. J. Struct. Biol. 2007, 160, 135–145. [Google Scholar] [CrossRef]
- Pavani, S.R.P.; Thompson, M.A.; Biteen, J.S.; Lord, S.J.; Liu, N.; Twieg, R.J.; Piestun, R.; Moerner, W. Three-dimensional, single-molecule fluorescence imaging beyond the diffraction limit by using a double-helix point spread function. Proc. Natl. Acad. Sci. USA 2009, 106, 2995–2999. [Google Scholar] [CrossRef]
- Holly, T.A.; Abbott, B.G.; Al-Mallah, M.; Calnon, D.A.; Cohen, M.C.; DiFilippo, F.P.; Ficaro, E.P.; Freeman, M.R.; Hendel, R.C.; Jain, D. Single photon-emission computed tomography. J. Nucl. Cardiol. 2010, 17, 941–973. [Google Scholar] [CrossRef] [PubMed]
- Rego, E.H.; Shao, L.; Macklin, J.J.; Winoto, L.; Johansson, G.A.; Kamps-Hughes, N.; Davidson, M.W.; Gustafsson, M.G. Nonlinear structured-illumination microscopy with a photoswitchable protein reveals cellular structures at 50-nm resolution. Proc. Natl. Acad. Sci. USA 2012, 109, E135–E143. [Google Scholar] [CrossRef] [PubMed]
- Rossy, J.; Cohen, E.; Gaus, K.; Owen, D.M. Method for co-cluster analysis in multichannel single-molecule localisation data. Histochem. Cell Biol. 2014, 141, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Vira, S.; Mekhedov, E.; Humphrey, G.; Blank, P.S. Fluorescent-labeled antibodies: Balancing functionality and degree of labeling. Anal. Biochem. 2010, 402, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Bates, M.; Huang, B.; Dempsey, G.T.; Zhuang, X. Multicolor super-resolution imaging with photo-switchable fluorescent probes. Science 2007, 317, 1749–1753. [Google Scholar] [CrossRef]
- Heilemann, M.; Van De Linde, S.; Schüttpelz, M.; Kasper, R.; Seefeldt, B.; Mukherjee, A.; Tinnefeld, P.; Sauer, M. Subdiffraction-resolution fluorescence imaging with conventional fluorescent probes. Angew. Chem. Int. Ed. 2008, 47, 6172–6176. [Google Scholar] [CrossRef] [PubMed]
- Granov, A.; Tiutin, L.; Schwarz, T. Positron Emission Tomography; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Zwan, M.D.; Bouwman, F.H.; Konijnenberg, E.; van der Flier, W.M.; Lammertsma, A.A.; Verhey, F.R.; Aalten, P.; van Berckel, B.N.; Scheltens, P. Diagnostic impact of [18F] flutemetamol pet in early-onset dementia. Alzheimers Res. 2017, 9, 2. [Google Scholar] [CrossRef]
- Marcus, C.; Mena, E.; Subramaniam, R.M. Brain pet in the diagnosis of alzheimer’s disease. Clin. Nucl. Med. 2014, 39, e413–e426. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.D.; Rabinovici, G.D.; Mathis, C.A.; Jagust, W.J.; Klunk, W.E.; Ikonomovic, M.D. Using pittsburgh compound b for in vivo pet imaging of fibrillar amyloid-beta. Adv. Pharm. 2012, 64, 27–81. [Google Scholar]
- Nickell, S.; Beck, F.; Korinek, A.; Mihalache, O.; Baumeister, W.; Plitzko, J.M. Automated cryoelectron microscopy of “single particles” applied to the 26s proteasome. Febs Lett. 2007, 581, 2751–2756. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Hirokawa, N.; Kikkawa, M. Three-dimensional structures of the flagellar dynein–microtubule complex by cryoelectron microscopy. J. Cell Biol. 2007, 177, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Hoenger, A. High-resolution cryo-electron microscopy on macromolecular complexes and cell organelles. Protoplasma 2014, 251, 417–427. [Google Scholar] [CrossRef]
- Yan, Q.; Cai, M.; Zhou, L.; Xu, H.; Shi, Y.; Sun, J.; Jiang, J.; Gao, J.; Wang, H. Using an RNA aptamer probe for super-resolution imaging of native egfr. Nanoscale Adv. 2019, 1, 291–298. [Google Scholar] [CrossRef]
- Fornasiero, E.F.; Opazo, F. Super-resolution imaging for cell biologists. Bioessays 2015, 37, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, J.Y.; Lee, T.S.; Song, I.H.; Cho, Y.L.; Chae, J.R.; Kang, H.; Lim, J.H.; Lee, J.H.; Kang, W.J. Pet imaging of her2 expression with an 18f-fluoride labeled aptamer. PLoS ONE 2019, 14, e0211047. [Google Scholar] [CrossRef]
- Hassanzadeh, L.; Chen, S.; Veedu, R.N. Radiolabeling of nucleic acid aptamers for highly sensitive disease-specific molecular imaging. Pharmaceuticals 2018, 11, 106. [Google Scholar] [CrossRef]
- Serrano-Santos, M.B.; Llobet, E.; Özalp, V.C.; Schäfer, T. Characterization of structural changes in aptamer films for controlled release nanodevices. Chem. Commun. 2012, 48, 10087–10089. [Google Scholar] [CrossRef] [PubMed]
- Mayer, G. The Chemical Biology of Nucleic Acids. Available online: https://onlinelibrary.wiley.com/doi/book/10.1002/9780470664001 (accessed on 14 May 2010).
- Stanlis, K.K.; McIntosh, J.R. Single-strand DNA aptamers as probes for protein localization in cells. J Histochem. Cytochem. 2003, 51, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Schermelleh, L.; Heintzmann, R.; Leonhardt, H. A guide to super-resolution fluorescence microscopy. J. Cell Biol. 2010, 190, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, G.T.; Vaughan, J.C.; Chen, K.H.; Bates, M.; Zhuang, X. Evaluation of fluorophores for optimal performance in localization-based super-resolution imaging. Nat. Methods 2011, 8, 1027–1036. [Google Scholar] [CrossRef]
- Lukinavicius, G.; Mitronova, G.Y.; Schnorrenberg, S.; Butkevich, A.N.; Barthel, H.; Belov, V.N.; Hell, S.W. Fluorescent dyes and probes for super-resolution microscopy of microtubules and tracheoles in living cells and tissues. Chem. Sci. 2018, 9, 3324–3334. [Google Scholar] [CrossRef]
- Kong, H.Y.; Byun, J. Nucleic acid aptamers: New methods for selection, stabilization, and application in biomedical science. Biomol. Ther. 2013, 21, 423. [Google Scholar] [CrossRef]
- Ozer, A.; Pagano, J.M.; Lis, J.T. New technologies provide quantum changes in the scale, speed, and success of selex methods and aptamer characterization. Mol. Ther. Nucleic Acids 2014, 3, e183. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.A. In Vivo Selex Strategies. Handbook of RNA Biochemistry: Second, Completely Revised and Enlarged Edition. Available online: https://onlinelibrary.wiley.com/doi/book/10.1002/9783527647064 (accessed on 28 March 2014).
- Szeto, K.; Craighead, H.G. Devices and approaches for generating specific high-affinity nucleic acid aptamers. Appl. Phys. Rev. 2014, 1, 031103. [Google Scholar] [CrossRef]
- Tombelli, S.; Minunni, M.; Mascini, M. Aptamers-based assays for diagnostics, environmental and food analysis. Biomol. Eng. 2007, 24, 191–200. [Google Scholar] [CrossRef] [PubMed]
- White, R.J.; Phares, N.; Lubin, A.A.; Xiao, Y.; Plaxco, K.W. Optimization of electrochemical aptamer-based sensors via optimization of probe packing density and surface chemistry. Langmuir 2008, 24, 10513–10518. [Google Scholar] [CrossRef]
- Nair, P.R.; Alam, M.A. Theory of "selectivity" of label-free nanobiosensors: A geometro-physical perspective. J. Appl. Phys. 2010, 107, 64701. [Google Scholar] [CrossRef]


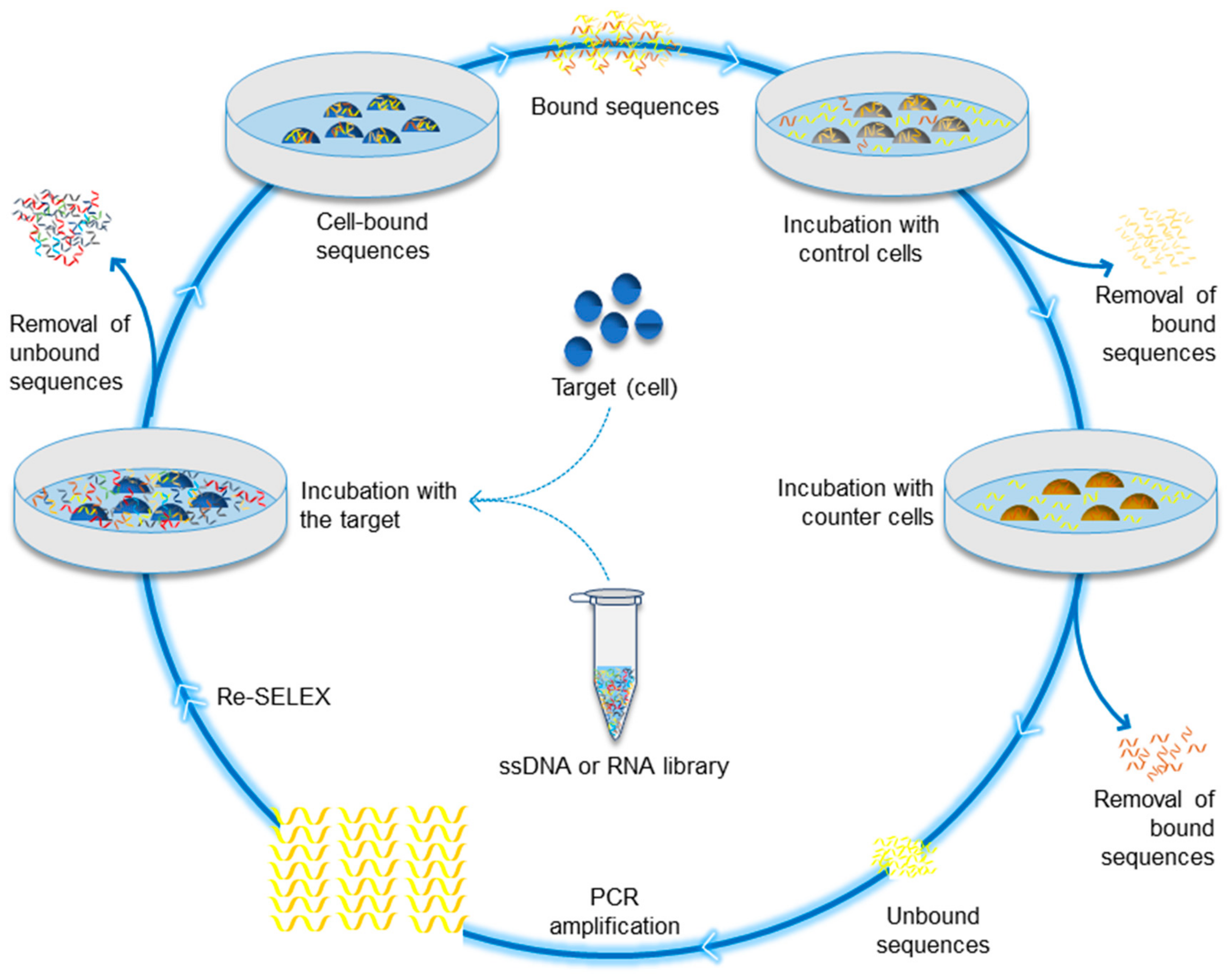{kind=link}
| SELEX Type | Principle | Aim | Application/Result | Year | Ref. |
|---|---|---|---|---|---|
| Negative SELEX | In vitro selection of single-stranded oligonucleotides against a target after removing sequences that bind to partitioning/immobilization matrices |
| Isolated ligand-specific aptamers that selectively bind to small molecules (dyes) | 1992 | [21] |
| In vivo SELEX | The selection and amplification steps occur inside the living cell using retroviral-based replication system | Produce aptamers that are functional in vivo | Provided a method to transfer aptamer selection and amplification from in vitro to in vivo | 1993 | [22,23] |
| Counter SELEX | Uses a second elution step against a molecule of similar target structure (e.g., caffeine, which differs from theophylline at the N-7 position) | Increase aptamer’s specificity and affinity towards target molecules | Isolated aptamer of high-specificity to theophylline | 1994 | [24] |
| cDNA-SELEX | Uses a preselected natural oligonucleotide pool that binds to the protein of interest rather than using a synthetic library | Identify natural library that interact with the protein of interest | Provided a new approach to study the interaction between the nucleotides and the protein of interest | 1995 | [25] |
| Blended SELEX | Forming a blended-pool through linking molecules (e.g., inhibitor) to a specific site on the library that will be subjected to SELEX |
|
| 1995 | [26] |
| Uses photoactivatable aptamers to crosslink and/or photoactive the target |
|
| 1995 2000 | [27,28] |
| Spiegelmer® | Selection of D-oligonucleotides against a mirrored target. Then, the D-oligonucleotides will be synthesized as L-oligonucleotides that can bind to unmirrored target | Increase aptamer stability against nucleases | Identified an endonuclease-stable L-RNAaptamer that binds to D-adenosine | 1996 | [29] |
| Magnetic bead-based SELEX | Uses magnetic beads for immobilization |
| Provided a new method to avoid DNA precipitation | 1997 | [30] |
| EMSA-SELEX | Uses EMSA partitioning at every cycle of the selection process | Identify a method to study oligonucleotides binding to proteins | Identified the role of specific zinc finger motifs in the Roaz protein that help in DNA‒protein binding and protein‒protein interaction | 1998 | [31] |
| Cell-SELEX | Identifies aptamers that bind specifically to different cells or targets on cell membrane |
| Identified aptamers that binds to specific cell (e.g., RBCs) or cell surface protein (e.g., variant surface glycoprotein of Trypanosoma brucei) | 1998 1999 | [32,33,34] |
| Chimeric SELEX | Fuses pre-selected aptamers of different targets together to form a combined small library followed by applying a dual selection pressure to select aptamers that can bind to multiple targets | Generate dual-function aptamers | Generated an engineered dual-function aptamer capable of testing/binding to two distinct features/targets | 1998 | [35] |
| Multi-stage SELEX | Modified chimeric-based SELEX followed by an extra selection with entirety of targets | Develop an allosteric synthetic aptamer | Provided a method to study the allosteric interactions in the DNA | 1999 | [36] |
| Indirect selection | Aptamer binds to a “primary target.” Binding to this target is required for aptamer bind ing to the main target | Develop a transition-state-bound aptamer that facilitates its binding to the main target | Generated Zn2+-dependent aptamer molecule for specific target binding to HIV-1 Tat protein | 2000 | [37] |
| Toggle-SELEX | A “polyclonal” aptamer selection strategy that generates aptamer capable of binding to multiple proteins through incubating the pre-selected aptamer with a second target until the aptamer can identify another region for binding | Select aptamers against homologous targets depending on cross-reactivity |
| 2001 | [38] |
| Truncates fixed regions present in the aptamers sequences that may limit the structure variations or reduce the ability of aptamer binding to the target |
| Provided a method to control aptamer-fixed sequences | 2001 2004 2008 2010 | [39,40,41,42] |
| Expression Cassette SELEX | Fuses a pre-selected aptamer with a polymerase III promoter “expression cassette” in a DNA plasmid | Produce highly expressed levels of functional aptamers |
| 2002 | [43] |
| NECEEM | Uses non-equilibrium conditions to separate the constituents of protein‒DNA mixture (free and bound components) |
| Provided a novel electrophoretic method for studying protein‒DNA interaction and Kd of the DNA‒protein complex | 2002 | [44] |
| Subtractive SELEX | Similar to counter SELEX where it adopts extra SELEX rounds to control target binding | Distinguish cell subtypes of homologous origin | Distinguished between differentiated and normal PC12 cells | 2003 | [45] |
| Tailored SELEX | Isolation of short aptamer through designing of cleavable sites to remove fixed sequences |
| Provided a new method to identify short fixed nucleotides aptamers through ligation and removal of primer | 2003 | [46] |
| CE-SELEX | Uses differential electrophoretic separation at every SELEX round after aptamer binds to the target |
| Standardized the SELEX method and made it more efficient | 2004 | [47] |
| SweepCE | Uses protein non-stopped flow in the capillary electrophoresis to form DNA‒protein complex that causes sweeping of the DNA |
| Provided a method that help in studying the equilibrium and kinetic parameters during protein‒DNA complex formation | 2004 | [48] |
| On-chip selection | On-chip selection method combined with point-mutation of the sequence and usage of genetic algorithm to produce aptamers | Generate different aptamers against different target molecules | Generated aptamers against resorufin | 2004 | [49] |
| ECEEM | Maintains the equilibrium of the aptamer-target while separating the components of the equilibrium mixture by capillary electrophoresis |
| Produced “smart aptamers” with a predefined Kd value matching the theoretically predicted value | 2005 | [50] |
| FluMag-SELEX | Magnetic beads-based method using fluorescent labelling for monitoring the affinity and complex separation |
|
| 2005 | [51] |
| Non-SELEX | NECEEM-based method in which the PCR amplification step is skipped and the recovered target‒DNA complex is incubated with fresh target followed by partitioning |
| Developed a technique to reduce the selection time while producing aptamers of a 4-fold affinity improvement over the CE-SELEX | 2006 | [52] |
| SPR-SELEX | A method that couples the aptamer or the target to a chip then immobilizes the target or the aptamer and measures the change at the surface upon aptamer-target binding |
| Provided a method to measure the aptamer’s real-time binding | 2006 | [53,54] |
| TECS-SELEX | Uses modified cells expressing recombinant form of a surface protein that eliminates the need for the purification step | Produce aptamer against ectopic cell surface protein | Produced aptamer against TGF-β type III receptor that is ectopically expressed on CHO cells | 2006 | [55] |
|
| Provide a method to study in vivo oligonucleotide‒protein binding |
| 2006 2012 | [56,57] |
| DeSELEX and Convergent selection | Complex SELEX methods where many proteins targets are incubated with the library and then deselecting the dominant-protein bound aptamer and redirecting the selection toward the less abundant protein/s | Shift the selection process toward a specific protein (even a less abundant one) in mixture of different protein complexes |
| 2007 | [58] |
| MonoLEX |
|
|
| 2007 | [59] |
| NanoSelection® (nM-AFM SELEX) |
|
| Developed a method to avoid multiple selection cycles and also select aptamers from small library | 2007 | [60] |
| ASExp | Uses quenching and dequenching of aptamers | Rapid selection method | Provided a method to rapidly select aptamer for different types of targets | 2008 | [61] |
| FACS-SELEX | Combines in vitro selection with FACS to separate specific cell population |
| Produced aptamer bound to vital Burkitt lymphoma cells | 2008 | [62,63] |
| Single microbead SELEX | Reduces the amount of the target molecules and expose them to a single microbead | Produce high affinity aptamer using a single target-conjugated microbead |
| 2008 | [64] |
| CLADE | In silico aptamer selection method | Rapid aptamer designing method | Identified aptamers against allophycocyanin in silico | 2008 | [65] |
| In silico selection | In silico secondary structure-based selection followed by a 3D structure prediction and HTP screening of the selected aptamers |
| Provided a computational approach to select aptamers of high-affinity | 2009 | [66] |
| Tissue slide-based SELEX | In situ SELEX method against paraffin tissue sections | Target clinical tumour markers and provide a way to differentiate between the clinical specimens |
| 2009 | [67] |
| Sol–gel SELEX |
| Produce faster, HTP, efficient, and cheap aptamer |
| 2009 | [68] |
|
|
|
| 2009 2010 2011 | [69,70,71] |
| M-SELEX | Target‒DNA sample separation is obtained through applying a high voltage |
| Provided a fast tool for screening aptamers against any targets | 2009 | [72,73,74,75] |
| QSAS | Combines M-SELEX with HTS for efficient and rapid aptamer production | Generate aptamer with high efficiency, high affinity, and high specificity | Introduced an integrated method for rapid, high-affinity, and high-specificity generation of aptamer | 2010 | [76] |
| Cellular-uptake in vivo-variant | Intravenous injection of modified random library into cancerous animal model | Select aptamer for internal target inside the organism depending on cellular uptake | Provided a unique approach to generate aptamer that can specifically localize to tumour cells in vivo | 2010 | [77] |
| SOMAmers |
|
|
| 2010 | [78] |
| ISM | In silico post-SELEX genetic algorithm performed to identify aptamers of high-binding affinity | Improve aptamers binding-affinity and specificity | Selected aptamers of high binding affinity and specificity to prostate specific antigen, VEGF, and Proteus mirabilis | 2010 2012 2013 2014 | [79,80,81,82] |
| μFFE | Applies electric field separation to overcome the limitations of CE-SELEX to separate bound from unbound sequences |
| Improved the library size by a 300-fold over what CE-SELEX has achieved | 2011 | [83] |
| HTS-SELEX | One round of positive selection followed by HTS and informatic analysis |
| Identified aptamers that can bind to thrombin in nM range | 2011 | [84] |
| One-step selection Method | One-step selection of target-immobilized coverslip followed by library binding, extensive washing, and amplification step |
| Provided a rapid one-step generation of aptamer against α-bungarotoxin | 2012 | [85] |
| Cell-internalization SELEX | Combines cell-SELEX with HTS and bioinformatics | Produce aptamers of high intracellular internalization | Provided a new approach to design aptamers that capable of internalizations into the cytoplasm of vascular smooth muscle cells | 2012 | [86] |
| Capture-SELEX (FluMag-based) | Immobilization of specific aptamers library on magnetic beads using a docking sequence linked to the library |
| Provided a method to select aptamer against small molecule (e.g., kanamycin A) | 2012 | [87] |
| Domain targeted SELEX | Uses recombinant protein containing chemokine domain (to extend protein accessibility) immobilized onto magnetic agarose beads | Select aptamer against specific protein’s unfolded domain | Developed aptamer to Fractalkine protein | 2012 | [88] |
| Uses π-π stacking and adsorption of ssDNA on 2D GO sheets to separate unbound DNA | Generation of high-affinity, cost-effective, and target-immobilization-free SELEX | Produced aptamer to Nampt protein without target immobilization | 2012 | [89] |
| MAI-SELEX | A selection method to recognize two separate sites on the target using 2’-fluoro-modified library | Target distinct sites/subunits of the protein | Produced two aptamers that recognize the αV or β3 subunits of integrin αVβ3 | 2012 | [90] |
| RAPID-SELEX |
| Reduce selection time | Identified aptamer to CHK2 and UBLCP1 in one-third of the time required for the conventional selection | 2013 | [91] |
| Expanded genetic alphabet (Ds-base) | Uses a library containing natural nucleotides and other three unnatural nucleotides with a hydrophobic base | Increase sequence diversity and so expand SELEX selection power | Generated aptamers that bind to VEGF165 and interferon-γ with more than 100-fold improved affinity | 2013 | [92] |
| AEGIS-SELEX | Uses unnatural forms of nucleotides, nonstandard P and Z, beside the normal four bases-based library (GACTZP DNA library) |
| Produced ZAP-2012 aptamer that can bind to MDA-MB-231 with 10-fold higher affinity than achieved using the conventional aptamers | 2014 | [93] |
| ES-SELEX |
| Generate epitope-specific aptamer of a native protein structure | Produced anti-sialic acid receptor aptamers that inhibit hemagglutination at pM range | 2014 | [94] |
| MARAS | Selects aptamers with different affinities to the target molecule using magnetic beads and external rotating magnetic field | Select aptamers based on their different binding affinities | Produced aptamer with high affinity to C-reactive protein | 2014 | [95] |
| Particle Display | Measures the affinity of each aptamer sequence in the library and sorting them using HTS. Then isolating the highest-affinities candidates using FACS | Generate high-affinity, simple, rapid, and cost-effective aptamers | Measured the affinity of 100 million aptamers and obtained high-affinity aptamers for thrombin, ApoE, PAI-1, and 4-1BB proteins | 2014 | [96] |
| MSD-SELEX | Uses a library of monoclonal DNA-displaying beads | Generate high-affinity and rapid aptamers | Obtained high-affinity aptamers against EpCAM and aflatoxin B1 | 2014 | [97] |
| Yeast surface display-SELEX | Target protein is loaded on the surface of yeast | Quick and unexpansive HTS identification of DNA-binding sites of the proteins without prior knowledge of the target site | Provided a way to determine aptamer‒protein binding specificity | 2014 | [98] |
| Hi-Fi SELEX | Introduces fixed-region blocking elements to enhance the functional diversity of the library |
| Identified aptamers of Kd in nM range to α-thrombin | 2015 | [99] |
| Click-SELEX | Uses copper(I)-catalysed alkyne–azide cycloaddition modified nucleic acid libraries | Generate modified nucleobases that increase binding capabilities |
| 2015 | [100,101] |
| Icell SELEX |
| Target various membrane proteins through successful manipulation of the endogenous expression of the target proteins | Produced aptamer to integrin alpha V | 2016 | [102] |
| LIGS | Uses antibody bound to antigen for the partitioning step to compete with aptamers from partially enriched SELEX | Identify highly-specific aptamer sequences that outcompeting with specific antibody for selecting highly specific aptamers | Identified three aptamers that outcompete with mIgM antibody | 2016 | [103] |
| Colorimetric dye-displacement SELEX | Uses target-induced displacement of a small-molecule dye to isolate signal-producing aptamers triggered by binding to the target | Develop an efficient and rapid way to isolate small molecules through colorimetric dye-displacement | Provided a new way of sensitive target isolation based on Cy7-displacement colorimetric assay | 2018 | [104] |
| Open qPCR SELEX | Uses open qPCR to quantify target-aptamer binding |
| Produced aptamers against whole Drosophila C virus particles | 2018 | [105] |
| Aptamer | Antibodies | |
|---|---|---|
| Molecular weight | Small (~12–30 kDa) | Relatively big (~150–180 kDa) |
| Secondary structures | Various structures: hairpin, loop, G-quadruplex, etc | β-sheets |
| Generation time | Few hours to months | Several months (~six months) |
| Batches variations | Low | High |
| Immunogenicity | Low | High |
| Minimal target size | Targets small sizes ~60 Da | ~600 Da |
| Targets | Wide range of targets | Immunogenic molecules |
| Shelf life | Long | Short |
| Allowed chemical modifications | Various modifications | Limited modifications |
| Nuclease degradation | Sensitive | Resistant |
| In vivo half-life | Short (~20 min) | Long (~one month) |
| Stability | Very stable | Sensitive to temperature and pH changes |
| Cost | Lower | Higher |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.H.; Elsherbiny, M.E.; Emara, M. Updates on Aptamer Research. Int. J. Mol. Sci. 2019, 20, 2511. https://doi.org/10.3390/ijms20102511
Ali MH, Elsherbiny ME, Emara M. Updates on Aptamer Research. International Journal of Molecular Sciences. 2019; 20(10):2511. https://doi.org/10.3390/ijms20102511
Chicago/Turabian StyleAli, Mohamed H., Marwa E. Elsherbiny, and Marwan Emara. 2019. "Updates on Aptamer Research" International Journal of Molecular Sciences 20, no. 10: 2511. https://doi.org/10.3390/ijms20102511
APA StyleAli, M. H., Elsherbiny, M. E., & Emara, M. (2019). Updates on Aptamer Research. International Journal of Molecular Sciences, 20(10), 2511. https://doi.org/10.3390/ijms20102511