Exercise and Sirtuins: A Way to Mitochondrial Health in Skeletal Muscle


Abstract
:1. Introduction
2. Skeletal Muscle, Sirtuins and Exercise
3. Effects of Different Type of Exercise on the Sirtuins in Human Skeletal Muscle
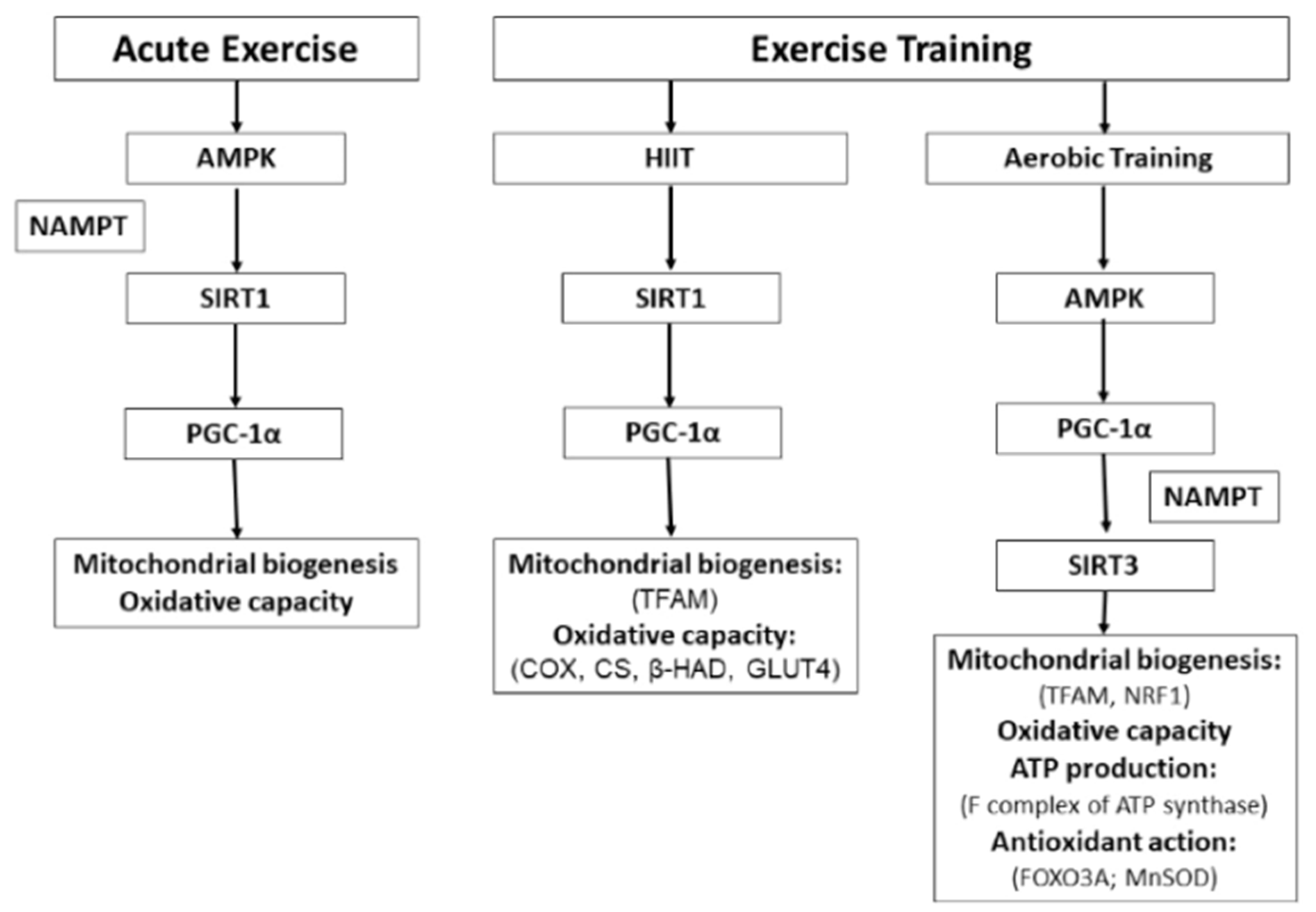
4. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| β-HAD | β-hydroxyacyl-CoA dehydrogenase |
| COX | Cytochrome C oxidase |
| CS | Citrate synthase |
| CREB | cAMP response element-binding |
| FOXO1A | Forkhead box O1 |
| FOXO3A | Forkhead box O3 |
| GLUT4 | Glucose transporter 4 |
| HIIT | High-intensity interval training |
| HDF | High fat diet |
| LPR130 | Leucine rich pentatricopeptide repeat containing |
| MnSOD | Manganese superoxide dismutase |
| NAD+ | Nicotinamide adenine dinucleotide |
| NAMPT | Nicotinamide phosphoribosyltransferase |
| NRF1 | Nuclear respiratory factor 1 |
| PGC-1α | Peroxisome proliferator-activated receptor gamma coactivator-1α |
| ROS | Reactive oxygen species |
| SIRT | Sirtuin |
| SIT | Sprint-interval training |
| TCA | Tricarboxylic acids |
| TFAM | Mitochondrial transcription factor A. |
References
- Anastasiou, D.; Krek, W. Sirt1: Linking adaptive cellular responses to aging-associated changes in organismal physiology. Physiology 2006, 21, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Dali-Youcef, N.; Lagouge, M.; Froelich, S.; Koehl, C.; Schoonjans, K.; Auwerx, J. Sirtuins: The ‘magnificent seven’, function, metabolism and longevity. Ann. Med. 2007, 39, 335–345. [Google Scholar] [CrossRef]
- Guarente, L.; Franklin, H. Epstein lecture: Sirtuins, aging, and medicine. N. Engl. J. Med. 2011, 364, 2235–2244. [Google Scholar] [CrossRef] [PubMed]
- Haigis, M.C.; Sinclair, D.A. Mammalian sirtuins: Biological insights and disease relevance. Annu. Rev. Pathol. 2010, 5, 253–295. [Google Scholar] [CrossRef] [PubMed]
- Blander, G.; Guarente, L. The sir2 family of protein deacetylases. Annu. Rev. Biochem. 2004, 73, 417–435. [Google Scholar] [CrossRef] [PubMed]
- Tanny, J.C.; Dowd, G.J.; Huang, J.; Hilz, H.; Moazed, D. An enzymatic activity in the yeast sir2 protein that is essential for gene silencing. Cell 1999, 99, 735–745. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef]
- Smith, A.G.; Muscat, G.E. Skeletal muscle and nuclear hormone receptors: Implications for cardiovascular and metabolic disease. Int. J. Biochem. Cell Biol. 2005, 37, 2047–2063. [Google Scholar] [CrossRef]
- Jing, E.; O’Neill, B.T.; Rardin, M.J.; Kleinridders, A.; Ilkeyeva, O.R.; Ussar, S.; Bain, J.R.; Lee, K.Y.; Verdin, E.M.; Newgard, C.B.; et al. Sirt3 regulates metabolic flexibility of skeletal muscle through reversible enzymatic deacetylation. Diabetes 2013, 62, 3404–3417. [Google Scholar] [CrossRef]
- Palacios, O.M.; Carmona, J.J.; Michan, S.; Chen, K.Y.; Manabe, Y.; Ward, J.L., 3rd; Goodyear, L.J.; Tong, Q. Diet and exercise signals regulate sirt3 and activate ampk and pgc-1alpha in skeletal muscle. Aging 2009, 1, 771–783. [Google Scholar] [CrossRef]
- Hokari, F.; Kawasaki, E.; Sakai, A.; Koshinaka, K.; Sakuma, K.; Kawanaka, K. Muscle contractile activity regulates sirt3 protein expression in rat skeletal muscles. J. Appl. Physiol. 2010, 109, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Bayod, S.; Del Valle, J.; Lalanza, J.F.; Sanchez-Roige, S.; de Luxan-Delgado, B.; Coto-Montes, A.; Canudas, A.M.; Camins, A.; Escorihuela, R.M.; Pallas, M. Long-term physical exercise induces changes in sirtuin 1 pathway and oxidative parameters in adult rat tissues. Exp. Gerontol. 2012, 47, 925–935. [Google Scholar] [CrossRef]
- White, A.T.; Schenk, S. Nad(+)/nadh and skeletal muscle mitochondrial adaptations to exercise. Am J. Physiol. Endocrinol. Metab. 2012, 303, E308–E321. [Google Scholar] [CrossRef] [PubMed]
- Gurd, B.J.; Holloway, G.P.; Yoshida, Y.; Bonen, A. In mammalian muscle, sirt3 is present in mitochondria and not in the nucleus; and sirt3 is upregulated by chronic muscle contraction in an adenosine monophosphate-activated protein kinase-independent manner. Metabolism 2012, 61, 733–741. [Google Scholar] [CrossRef]
- Nakamura, Y.; Ogura, M.; Tanaka, D.; Inagaki, N. Localization of mouse mitochondrial sirt proteins: Shift of sirt3 to nucleus by co-expression with sirt5. Biochem. Biophys. Res. Commun. 2008, 366, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Vassilopoulos, A.; Pennington, J.D.; Andresson, T.; Rees, D.M.; Bosley, A.D.; Fearnley, I.M.; Ham, A.; Flynn, C.R.; Hill, S.; Rose, K.L.; et al. Sirt3 deacetylates atp synthase f1 complex proteins in response to nutrient- and exercise-induced stress. Antioxid Redox Signal 2014, 21, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Coleman, M.C.; Pennington, J.D.; Ozden, O.; Park, S.H.; Jiang, H.; Kim, H.S.; Flynn, C.R.; Hill, S.; Hayes McDonald, W.; et al. Sirt3-mediated deacetylation of evolutionarily conserved lysine 122 regulates mnsod activity in response to stress. Mol. Cell 2010, 40, 893–904. [Google Scholar] [CrossRef]
- Shi, Z.; Li, C.; Yin, Y.; Yang, Z.; Xue, H.; Mu, N.; Wang, Y.; Liu, M.; Ma, H. Aerobic interval training regulated sirt3 attenuates high-fat-diet-associated cognitive dysfunction. Biomed. Res. Int. 2018, 2018, 2708491. [Google Scholar] [CrossRef]
- Lanza, I.R.; Short, D.K.; Short, K.R.; Raghavakaimal, S.; Basu, R.; Joyner, M.J.; McConnell, J.P.; Nair, K.S. Endurance exercise as a countermeasure for aging. Diabetes 2008, 57, 2933–2942. [Google Scholar] [CrossRef]
- Villanova, L.; Vernucci, E.; Pucci, B.; Pellegrini, L.; Nebbioso, M.; Mauri, C.; Marfe, G.; Spataro, A.; Fini, M.; Banfi, G.; et al. Influence of age and physical exercise on sirtuin activity in humans. J. Biol. Regul. Homeost. Agents 2013, 27, 497–507. [Google Scholar]
- Koltai, E.; Bori, Z.; Osvath, P.; Ihasz, F.; Peter, S.; Toth, G.; Degens, H.; Rittweger, J.; Boldogh, I.; Radak, Z. Master athletes have higher mir-7, sirt3 and sod2 expression in skeletal muscle than age-matched sedentary controls. Redox Biol. 2018, 19, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Midgley, A.C.; Morris, G.; Phillips, A.O.; Steadman, R. 17beta-estradiol ameliorates age-associated loss of fibroblast function by attenuating ifn-gamma/stat1-dependent mir-7 upregulation. Aging Cell 2016, 15, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Bharathi, S.S.; Zhang, Y.; Mohsen, A.W.; Uppala, R.; Balasubramani, M.; Schreiber, E.; Uechi, G.; Beck, M.E.; Rardin, M.J.; Vockley, J.; et al. Sirtuin 3 (sirt3) protein regulates long-chain acyl-coa dehydrogenase by deacetylating conserved lysines near the active site. J. Biol. Chem. 2013, 288, 33837–33847. [Google Scholar] [CrossRef]
- Zhang, B.; Cui, S.; Bai, X.; Zhuo, L.; Sun, X.; Hong, Q.; Fu, B.; Wang, J.; Chen, X.; Cai, G. Sirt3 overexpression antagonizes high glucose accelerated cellular senescence in human diploid fibroblasts via the sirt3-foxo1 signaling pathway. Age 2013, 35, 2237–2253. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Brown, K.; Hirschey, M.D.; Verdin, E.; Chen, D. Calorie restriction reduces oxidative stress by sirt3-mediated sod2 activation. Cell Metab. 2010, 12, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Guadalupe-Grau, A.; Fuentes, T.; Ponce-Gonzalez, J.G.; Morales-Alamo, D.; Olmedillas, H.; Guillen-Salgado, J.; Santana, A.; Calbet, J.A. Sirt1, amp-activated protein kinase phosphorylation and downstream kinases in response to a single bout of sprint exercise: Influence of glucose ingestion. Eur. J. Appl. Physiol. 2010, 109, 731–743. [Google Scholar] [CrossRef]
- Edgett, B.A.; Bonafiglia, J.T.; Baechler, B.L.; Quadrilatero, J.; Gurd, B.J. The effect of acute and chronic sprint-interval training on lrp130, sirt3, and pgc-1alpha expression in human skeletal muscle. Physiol. Rep. 2016, 4. [Google Scholar] [CrossRef]
- Edgett, B.A.; Hughes, M.C.; Matusiak, J.B.; Perry, C.G.; Simpson, C.A.; Gurd, B.J. Sirt3 gene expression but not sirt3 subcellular localization is altered in response to fasting and exercise in human skeletal muscle. Exp. Physiol. 2016, 101, 1101–1113. [Google Scholar] [CrossRef]
- Radak, Z.; Bori, Z.; Koltai, E.; Fatouros, I.G.; Jamurtas, A.Z.; Douroudos, I.I.; Terzis, G.; Nikolaidis, M.G.; Chatzinikolaou, A.; Sovatzidis, A.; et al. Age-dependent changes in 8-oxoguanine-DNA glycosylase activity are modulated by adaptive responses to physical exercise in human skeletal muscle. Free Radic. Biol. Med. 2011, 51, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Dumke, C.L.; Mark Davis, J.; Angela Murphy, E.; Nieman, D.C.; Carmichael, M.D.; Quindry, J.C.; Travis Triplett, N.; Utter, A.C.; Gross Gowin, S.J.; Henson, D.A.; et al. Successive bouts of cycling stimulates genes associated with mitochondrial biogenesis. Eur. J. Appl. Physiol. 2009, 107, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Little, J.P.; Safdar, A.; Wilkin, G.P.; Tarnopolsky, M.A.; Gibala, M.J. A practical model of low-volume high-intensity interval training induces mitochondrial biogenesis in human skeletal muscle: Potential mechanisms. J. Physiol. 2010, 588, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Gurd, B.J.; Yoshida, Y.; McFarlan, J.T.; Holloway, G.P.; Moyes, C.D.; Heigenhauser, G.J.; Spriet, L.; Bonen, A. Nuclear sirt1 activity, but not protein content, regulates mitochondrial biogenesis in rat and human skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R67–R75. [Google Scholar] [CrossRef]
- Gurd, B.J.; Perry, C.G.; Heigenhauser, G.J.; Spriet, L.L.; Bonen, A. High-intensity interval training increases sirt1 activity in human skeletal muscle. Appl. Physiol. Nutr. Metab. 2010, 35, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.L.; Irving, B.A.; Lanza, I.R.; Vendelbo, M.H.; Konopka, A.R.; Robinson, M.M.; Henderson, G.C.; Klaus, K.A.; Morse, D.M.; Heppelmann, C.; et al. Differential effect of endurance training on mitochondrial protein damage, degradation, and acetylation in the context of aging. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 1386–1393. [Google Scholar] [CrossRef] [PubMed]
- Brandauer, J.; Andersen, M.A.; Kellezi, H.; Risis, S.; Frosig, C.; Vienberg, S.G.; Treebak, J.T. Amp-activated protein kinase controls exercise training- and aicar-induced increases in sirt3 and mnsod. Front. Physiol. 2015, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Ortiz, K.; Perez-Vazquez, V.; Diaz-Cisneros, F.J.; Figueroa, A.; Jimenez-Flores, L.M.; Rodriguez-DelaRosa, G.; Macias, M.H. Aerobic training increases expression levels of sirt3 and pgc-1alpha in skeletal muscle of overweight adolescents without change in caloric intake. Pediatr. Exerc. Sci. 2015, 27, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Ortiz, K.; Perez-Vazquez, V.; Figueroa, A.; Diaz, F.J.; Montano-Ascencio, P.G.; Macias-Cervantes, M.H. Aerobic training but no resistance training increases sirt3 in skeletal muscle of sedentary obese male adolescents. Eur. J. Sport Sci. 2018, 18, 226–234. [Google Scholar] [CrossRef]
- Canto, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. Ampk regulates energy expenditure by modulating nad+ metabolism and sirt1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef]
- Gerhart-Hines, Z.; Rodgers, J.T.; Bare, O.; Lerin, C.; Kim, S.H.; Mostoslavsky, R.; Alt, F.W.; Wu, Z.; Puigserver, P. Metabolic control of muscle mitochondrial function and fatty acid oxidation through sirt1/pgc-1alpha. EMBO J. 2007, 26, 1913–1923. [Google Scholar] [CrossRef]
- Wright, D.C.; Han, D.H.; Garcia-Roves, P.M.; Geiger, P.C.; Jones, T.E.; Holloszy, J.O. Exercise-induced mitochondrial biogenesis begins before the increase in muscle pgc-1alpha expression. J. Biol. Chem. 2007, 282, 194–199. [Google Scholar] [CrossRef]
- Cooper, M.P.; Uldry, M.; Kajimura, S.; Arany, Z.; Spiegelman, B.M. Modulation of pgc-1 coactivator pathways in brown fat differentiation through lrp130. J. Biol. Chem. 2008, 283, 31960–31967. [Google Scholar] [CrossRef] [PubMed]
- Gohil, V.M.; Nilsson, R.; Belcher-Timme, C.A.; Luo, B.; Root, D.E.; Mootha, V.K. Mitochondrial and nuclear genomic responses to loss of lrpprc expression. J. Biol. Chem. 2010, 285, 13742–13747. [Google Scholar] [CrossRef]
- Liu, L.; Sanosaka, M.; Lei, S.; Bestwick, M.L.; Frey, J.H., Jr.; Surovtseva, Y.V.; Shadel, G.S.; Cooper, M.P. Lrp130 protein remodels mitochondria and stimulates fatty acid oxidation. J. Biol. Chem. 2011, 286, 41253–41264. [Google Scholar] [CrossRef] [PubMed]
- Gibala, M.J.; McGee, S.L.; Garnham, A.P.; Howlett, K.F.; Snow, R.J.; Hargreaves, M. Brief intense interval exercise activates ampk and p38 mapk signaling and increases the expression of pgc-1alpha in human skeletal muscle. J. Appl. Physiol. 2009, 106, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Hallows, W.C.; Lee, S.; Denu, J.M. Sirtuins deacetylate and activate mammalian acetyl-coa synthetases. Proc. Natl. Acad. Sci. USA 2006, 103, 10230–10235. [Google Scholar] [CrossRef]
- Someya, S.; Yu, W.; Hallows, W.C.; Xu, J.; Vann, J.M.; Leeuwenburgh, C.; Tanokura, M.; Denu, J.M.; Prolla, T.A. Sirt3 mediates reduction of oxidative damage and prevention of age-related hearing loss under caloric restriction. Cell 2010, 143, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Tseng, A.H.; Shieh, S.S.; Wang, D.L. Sirt3 deacetylates foxo3 to protect mitochondria against oxidative damage. Free Radic. Biol. Med. 2013, 63, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Schwer, B.; Bunkenborg, J.; Verdin, R.O.; Andersen, J.S.; Verdin, E. Reversible lysine acetylation controls the activity of the mitochondrial enzyme acetyl-coa synthetase 2. Proc. Natl. Acad. Sci. USA 2006, 103, 10224–10229. [Google Scholar] [CrossRef]
- Wu, Y.T.; Lee, H.C.; Liao, C.C.; Wei, Y.H. Regulation of mitochondrial f(o)f(1)atpase activity by sirt3-catalyzed deacetylation and its deficiency in human cells harboring 4977bp deletion of mitochondrial DNA. Biochim. Biophys. Acta 2013, 1832, 216–227. [Google Scholar] [CrossRef]
- Jacobs, K.M.; Pennington, J.D.; Bisht, K.S.; Aykin-Burns, N.; Kim, H.S.; Mishra, M.; Sun, L.; Nguyen, P.; Ahn, B.H.; Leclerc, J.; et al. Sirt3 interacts with the daf-16 homolog foxo3a in the mitochondria, as well as increases foxo3a dependent gene expression. Int. J. Biol. Sci. 2008, 4, 291–299. [Google Scholar] [CrossRef]
- Reid, M.B. Invited review: Redox modulation of skeletal muscle contraction: What we know and what we don’t. J. Appl. Physiol. 2001, 90, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, H.C.; Fujita, S.; Cadenas, J.G.; Chinkes, D.L.; Volpi, E.; Rasmussen, B.B. Resistance exercise increases ampk activity and reduces 4e-bp1 phosphorylation and protein synthesis in human skeletal muscle. J. Physiol. 2006, 576, 613–624. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type of Exercise | Mode | SIRT1 | SIRT3 | PGC-1α |
|---|---|---|---|---|
| Acute | Wingate test [26] | ↑ (Prot) | - | = (Prot) |
| SIT [27] | - | = (mRNA) | ↑ (mRNA) | |
| Stationary bicycle [28] | - | ↓(mRNA) = (Prot) | - | |
| Treadmill [29] | ↑ (mRNA) (*S) = (mRNA) | = (mRNA) (*S) ↓(mRNA) | - | |
| Successive Bouts | 3 bouts of 3 h cycling [30] | ↑ (mRNA) | - | ↑ (mRNA) |
| HIIT | 6 sessions low-volume [31] | ↑ (Prot) | - | = Total (Prot) ↑ Nuclear (Prot) |
| 7 sessions [32] | ↑ (act) = (Prot) | - | ↑ Nuclear (Prot) ↑ Total (Prot) | |
| 18 sessions [33] | ↓(Prot) ↑ (act) | - | ↑ (Prot) | |
| 24 sessions SIT [27] | - | = (Prot) | = (Prot) | |
| Aerobic Training | 24 sessions Treadmill [34] | - | ↑ (Prot) (*S) | - |
| 15 Sessions one-legged knee extensor [35] | - | = (Prot) | - | |
| 36 sessions Cycle ergometer [36] | - | ↑ (Prot) (*S) (OW) | ↑ (Prot) (*S) (OW) | |
| Resistance Training | 36 sessions 11 exercises for the major muscle groups [37] | - | = (Prot) (*S) (OW) | = (Prot) (*S) (OW) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas-Ortiz, K.; Pérez-Vázquez, V.; Macías-Cervantes, M.H. Exercise and Sirtuins: A Way to Mitochondrial Health in Skeletal Muscle. Int. J. Mol. Sci. 2019, 20, 2717. https://doi.org/10.3390/ijms20112717
Vargas-Ortiz K, Pérez-Vázquez V, Macías-Cervantes MH. Exercise and Sirtuins: A Way to Mitochondrial Health in Skeletal Muscle. International Journal of Molecular Sciences. 2019; 20(11):2717. https://doi.org/10.3390/ijms20112717
Chicago/Turabian StyleVargas-Ortiz, Katya, Victoriano Pérez-Vázquez, and Maciste H. Macías-Cervantes. 2019. "Exercise and Sirtuins: A Way to Mitochondrial Health in Skeletal Muscle" International Journal of Molecular Sciences 20, no. 11: 2717. https://doi.org/10.3390/ijms20112717
APA StyleVargas-Ortiz, K., Pérez-Vázquez, V., & Macías-Cervantes, M. H. (2019). Exercise and Sirtuins: A Way to Mitochondrial Health in Skeletal Muscle. International Journal of Molecular Sciences, 20(11), 2717. https://doi.org/10.3390/ijms20112717