1. Introduction
Wheat (
Triticum aestivum L.) is a cereal grain, originating from the Near East but now being grown all over the globe, occupying the largest land area compared to other commercial food crops, which, however, makes it face multiple biotic and abiotic stresses. The fungal leaf spotting diseases complex comprising tan spot (TS, caused by
Pyrenophora tritici-repentis (Died.) Drechs.), Septoria nodorum blotch (SNB,
Parastagonospora nodorum (Berk.) Quaedvlieg, Verkley & Crous), Septoria tritici blotch (STB,
Zymoseptoria tritici (Desm.) Quaedvlieg & Crous) and spot blotch (SB,
Cochliobolus sativus (Ito & Kuribayashi) Drechs. Ex Dastur) cause major damage to wheat production globally. Among these diseases, TS and SNB in recent decades have observed significant increases globally [
1]. On average, TS and SNB cause generally 5 to 10% yield losses; however, under conditions favourable for disease development, yield losses of up to 50% have been reported [
2,
3]. Murray and Brennan [
4] reported that in Australia the three most devastating pathogens were
P. tritici-repentis,
Puccinia striiformis and
P. nodorum, with an average annual losses caused by the diseases induced by these pathogen amounting to
$212 million,
$127 million and
$108 million, respectively. Serious yield losses due to TS and SNB occur due to a reduction in the photosynthetic area of leaves resulting in reduced grain fill, lower test weight, kernel shriveling, and lower number of kernel per head [
3,
5]. TS and SNB also render red or dark smudges and black points to the infected kernels, significantly downgrading wheat quality [
5,
6].
Isolates of the fungus
P. tritici-repentis (Died.) Drechs. [anamorph
Drechslera tritici-repentis (Died.) Shoem.] differ in virulence and have been race classified. Susceptibility of wheat to
P. tritici-repentis is shown via the development of tan necrosis and/or extensive chlorosis symptoms depending on the race of the pathogen and the susceptibility in the host [
5]. Presently, eight races of
P. tritici-repentis have been identified worldwide based on their virulence pattern on a set of wheat differential cultivars involving Glenlea, 6B-365, 6B-662 and Salamouni/Erik [
7] and on the presence of genes coding for specific host-selective toxins (HSTs) [
8]. Race structure in the fungus
P. nodorum (Berk.) Quaedvlieg, Verkley & Crous (anamorph
Stagonospora nodorum (Berk.) E. Castell. & Germano) is not universally accepted; significant variation in aggressiveness among isolates is observed.
P. nodorum produces multiple HSTs that contribute to variation in aggressiveness and for infection to occur, the host must produce the target receptor protein recognized by the toxin [
9].
Genetic analysis of wheat-
P. tritici-repentis interactions have revealed that host resistance ranges from qualitative [
10,
11,
12,
13,
14] to quantitative control [
15,
16,
17,
18,
19,
20]. Some studies utilizing quantitative trait loci (QTL) analysis reported resistance to TS as polygenic, but observed that previously reported major genes frequently underline these QTLs [
18,
20,
21]. Isolates of the fungus
P. tritici-repentis are known to produce at least three HSTs; Ptr ToxA [
22], Ptr ToxB [
23] and Ptr ToxC [
24] that interact with products of specific host sensitivity genes located on chromosome arm 5BL [
10,
11], 2BS [
25] and 1AS [
24], respectively, to cause disease. However, Friesen, et al. [
26] observed that insensitivity to HST Ptr ToxA accounted for only a part of the resistance to the fungus and pointed out that Ptr ToxA was a virulence factor instead of a pathogenicity factor. Similar findings were observed by other studies [
16,
27], indicating that there may be other mechanisms than HSTs involved in disease development including non-toxin factors, host non-specific toxins, and unknown host-specific toxins [
7].
For SNB, although several reports have identified a single dominant gene conferring resistance [
28,
29], there are studies that testified the genetics of adult plant resistance is quantitative [
20,
30,
31,
32,
33,
34]. So far, there have been 15 HSTs found, of which, eight (SnToxA, SnTox1, SnTox2, SnTox3, SnTox4, SnTox5, SnTox6 and SnTox7) were characterized from
P. nodorum, which interacted with host genes such as
Tsn1,
Snn1,
Snn2,
Snn3,
Snn4,
Snn5,
Snn6 and
Snn7 [
35,
36,
37]. In order to understand the inheritance of SNB resistance, genetic mapping and QTL analysis have been researched [
9,
20,
32,
38,
39]. In the last decade, there were some reports on QTLs for seedling resistance and adult plant resistance [
39,
40]. The identified QTLs of seedling resistance were located on 1B, 2B, 2D, 3A, 4A, 4B, 5A, 5B, 5D, 6A, and 7A chromosomes [
35,
41,
42]. Among them, some genomic regions contributed to adult plant resistance. For example, the QTL on 5BL at the
Tsn1 locus for seedling resistance was in the same marker interval as the QTL for flag leaf resistance [
35], similar to those on 2D, 4B and 5A chromosomes [
35,
40]. Some QTLs for flag leaf resistance have been found to be associated with the host genes, such as on 1B with
Snn1, on 2D with
Snn2, and on 5B with
Tsn1.
One of the major objectives of the CIMMYT’s wheat breeding program is to develop superior germplasm with durable resistance to multiple diseases including TS and SNB. Identification, characterization and transfer of new resistance genes from different resistant sources are always underway to enhance the level and diversity of resistance. These diverse genetic resources potentially possessing multiple resistance genes are then incorporated into breeding programs to develop durable multiple disease resistant germplasm. The mega variety PBW343 is a high yielding, widely adapted and very popular cultivar in India, which is resistant to TS and SNB but susceptible to stem rust Ug99. Kenya Nyangumi (KN) shows adult plant resistance to Ug99 but is susceptible to TS and SNB. Primarily, the population PBW343/KN was developed to decipher the adult plant resistance to Ug99; however, since this population segregates for TS and SNB as well, this study was initiated to determine and map the resistance to TS and SNB.
3. Discussion
Since its inception, CIMMYT’s primary goal has been to develop superior wheat germplasm incorporating broad-spectrum disease resistance, homogeneity in agronomic traits that impart high yields, superior quality, adaptability and stability. One of the strategies of developing disease resistant germplasm involves breeding for adult plant resistance (APR), which is associated with non-hypersensitive reaction, race-nonspecific resistance, quantitatively inherited and possibly effective against multiple diseases. The focus on APR was based on the fact that stacking multiple genes of small effects leads to high levels of resistance, which is highly durable due to its quantitative resistance nature [
43]. Previous reports have indicated that the race-nonspecific resistance genes
Lr34 and
Lr46 confer broad-spectrum resistance to biotrophic diseases like rusts and powdery mildew and hemi-biotrophic disease like spot blotch [
44].
TS and SNB are two major components of the destructive leaf-spotting diseases complex of wheat. Resistant varieties are regarded as the most efficient and effective strategies of disease management, but the information on resistance genes or QTLs is a prerequisite for development of broad genetic based durable resistant cultivars. Additionally, to hasten the process and progress of marker-assisted selection (MAS) it is paramount that new resistance genes are precisely tagged. Many studies on QTL mapping for TS and SNB have been reported [
9,
19,
35,
45,
46,
47], but most of them were aimed at single diseases. The RIL population used in this study was not only characterized for TS and SNB, but also for stem rust [
48]. It is interesting to note that some important QTLs found on 5BL were associated with all three diseases, which implied multi-disease resistance QTLs.
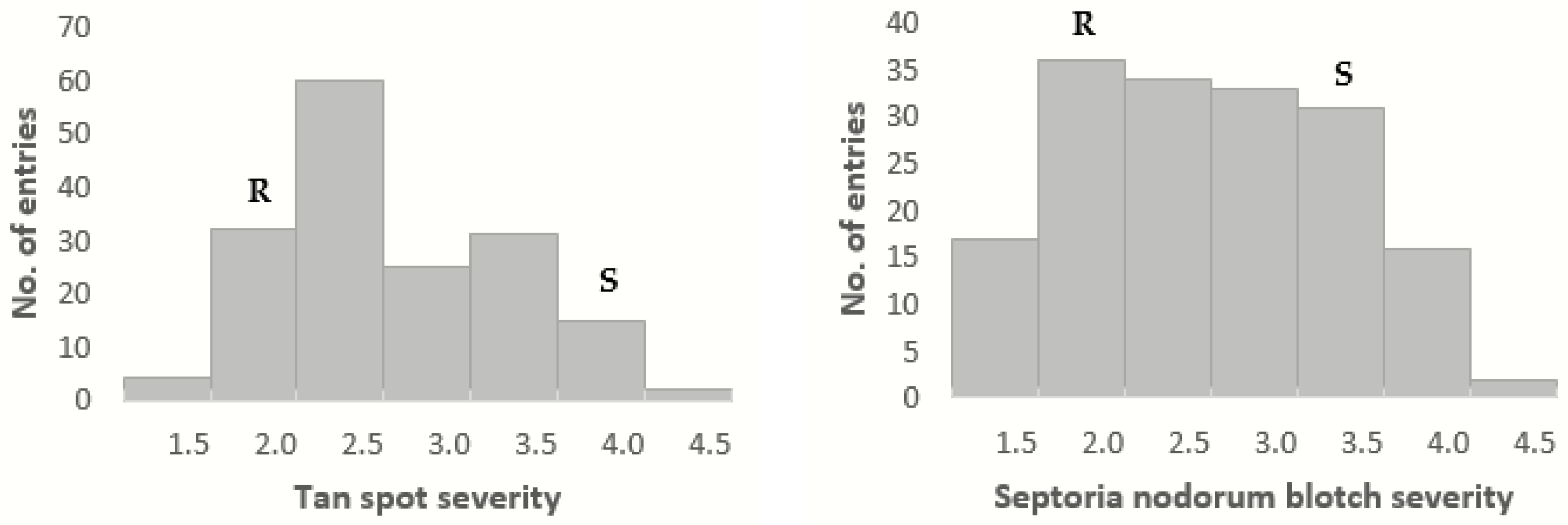
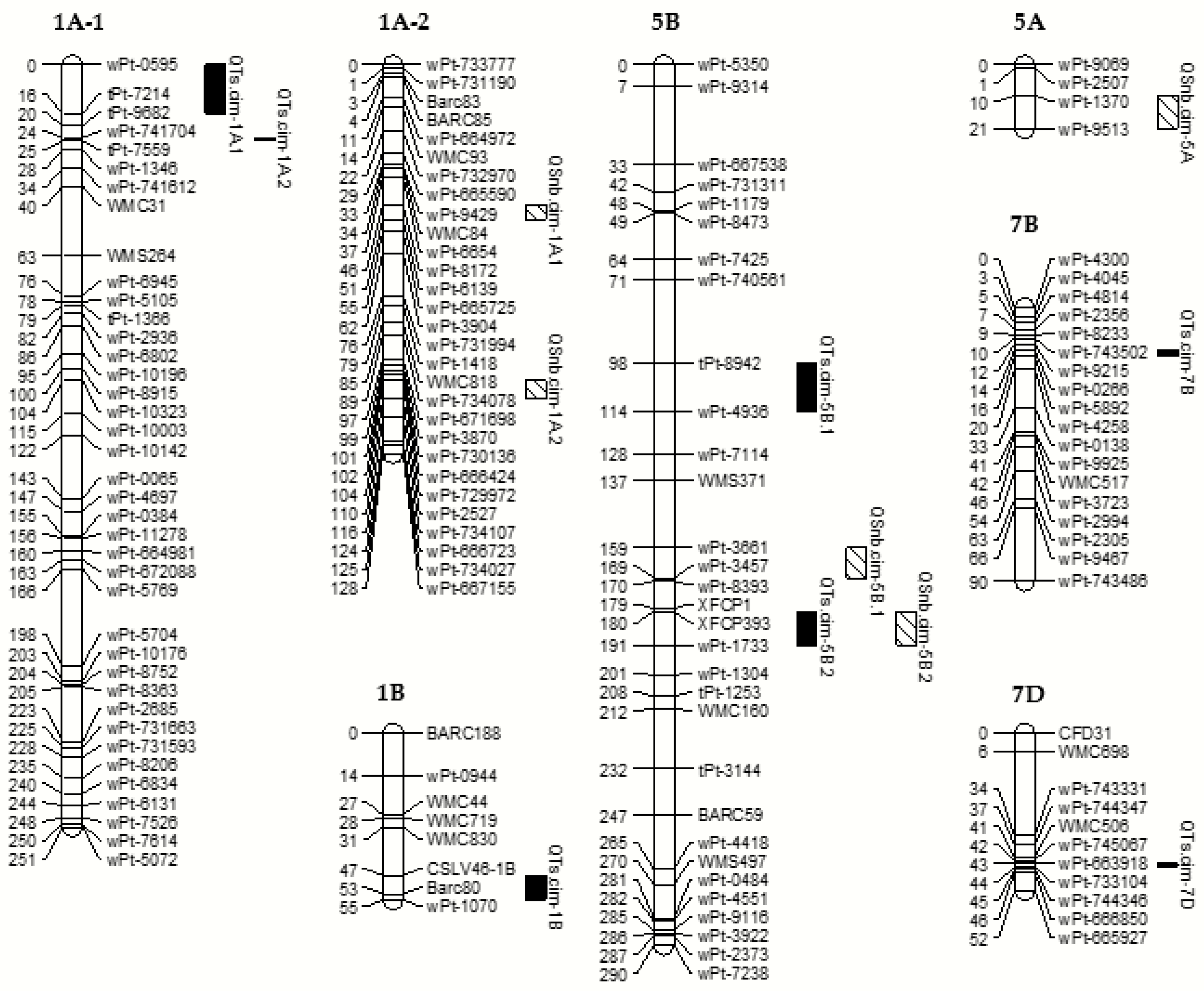
In this research, we identified seven TS QTLs, contributed mostly by PBW343, of which
QTs.cim-5B.2 on chromosome 5BL showed the biggest phenotypic effects. This QTL was reported in previous research and corresponds to
Tsn1 [
14,
20,
46,
49]. Cheong et al. [
15] and Singh et al. [
18] also found QTLs corresponding or close to the location of
Tsn1 (
Tsr1) with PVE of 39% and 18%, respectively. Molecular markers closely linked to this locus have been identified and are available for MAS [
50]. Another major QTL was detected on 1AS, where
QTsc.ndsu-1A was initially reported by Faris et al. [
27] and validated by Effertz et al. [
21,
24] to be associated with chlorosis in both adult plants and seedlings. Sun et al. [
51] identified
QTs.ksu-1AS in this region, whose position coincided with
Tsc1, and its effect was attributed to the Tsc1-Ptr ToxC interaction. Faris et al. [
46] identified molecular markers linked to
Tsc1, of which several STS makers could be useful in MAS. QTLs on 1BS, 7BS and 7DS chromosomes with lower PVE were detected in the current study, some of which might be the same as reported previously. On 1BS, Faris and Friesen [
17] identified a race-nonspecific QTL, explaining from 13 to 29% of the phenotypic variation. On 7BS, Faris, et al. [
52] identified a race-nonspecific QTL with PVE ranging from 5 to 8%. On 7DS, Faris and Friesen [
17] detected a minor QTL, but it was specific for race 3, suggesting that the 7DS QTL found in our study might be a new one.
There were also many reports on QTLs for SNB resistance in both seedling and adult plant. Most of them were located on 1BS, 2AS, 2BS, 2D, 3BS, 4BL, 5AL, 5BL, 5DL, 6AL and 7BL chromosomes [
35], of which, the 5BL QTL was the most frequently reported, for which four out of six reports were on seedling resistance. Similarly, two QTLs on 5BL were identified in the present study, with one being a major and the other a minor QTL. The major QTL was close to the
Tsn1 locus according to their linked markers. Additionally, a major QTL was found on 5AS, which has not been reported so far and thus may be a new QTL for SNB resistance. Two more possible new QTLs were detected on the 1A chromosome, one of which showed major effects.
In this population, we detected QTLs on 5BL for both TS and SNB resistance, and Singh et al. [
48] also found a QTL,
QSr.cim-5B for stem rust resistance, in a similar region. These results indicated that the 5BL QTL region could be very important for multi-disease resistance, and further studies are needed to see if it confers resistance to other diseases. PCR-based markers linked to this locus are available for MAS [
48]. Another possible multi-disease resistant locus in this population was
QTs.cim-1B for TS resistance, which was closely linked to the resistance gene
Lr46/Yr29/Pm39 for rusts and powdery mildew, suggesting that
Lr46/Yr29/Pm39 might also be able to contribute to TS resistance, but their mechanism of interaction needs to be further studied. Despite this, the functional marker for this locus is available for MAS [
53].
The findings of this study suggest that the genomic regions contributing to TS and SNB resistance may also confer resistance to other diseases, such as rusts and powdery mildew resistance [
47,
54], which perhaps indicates there are some race/pathogen-nonspecific resistance genes. Therefore, utilization of those broad-spectrum resistance genes/QTLs is recommended for breeding superior multi-disease resistant wheat cultivars, whereas the disease-specific QTLs identified in the current study are useful for breeding programs aimed at TS and SNB resistance.


 ,
, 


{kind=link}
{kind=link}