Transforming Growth Factor Beta 3-Loaded Decellularized Equine Tendon Matrix for Orthopedic Tissue Engineering

Abstract
1. Introduction
2. Results
2.1. Scaffold Loading with TGFβ3 and Its Efficiency
2.2. Bioactivity of Scaffold-Associated TGFβ3
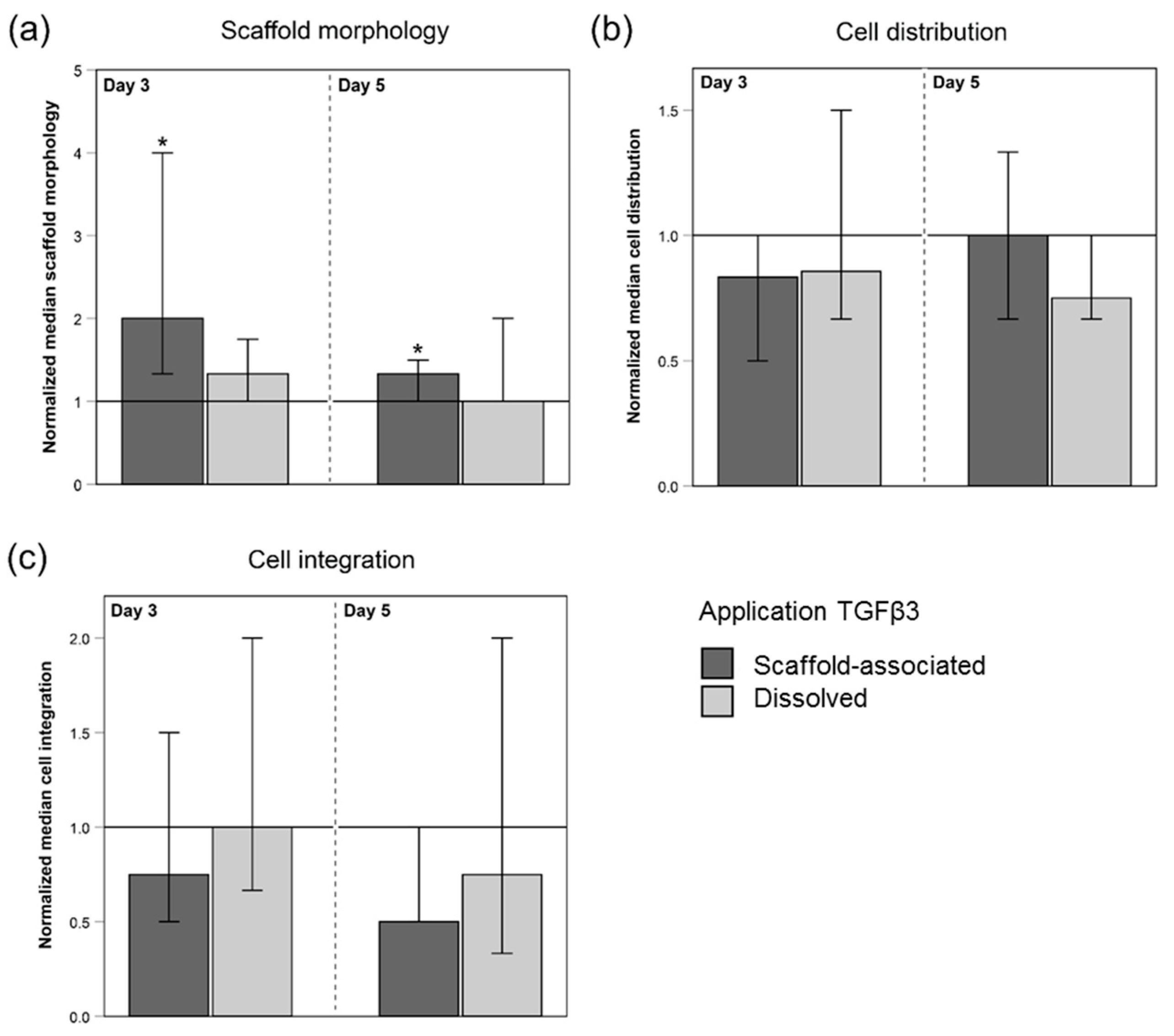
2.2.1. Macroscopic Scaffold Morphology
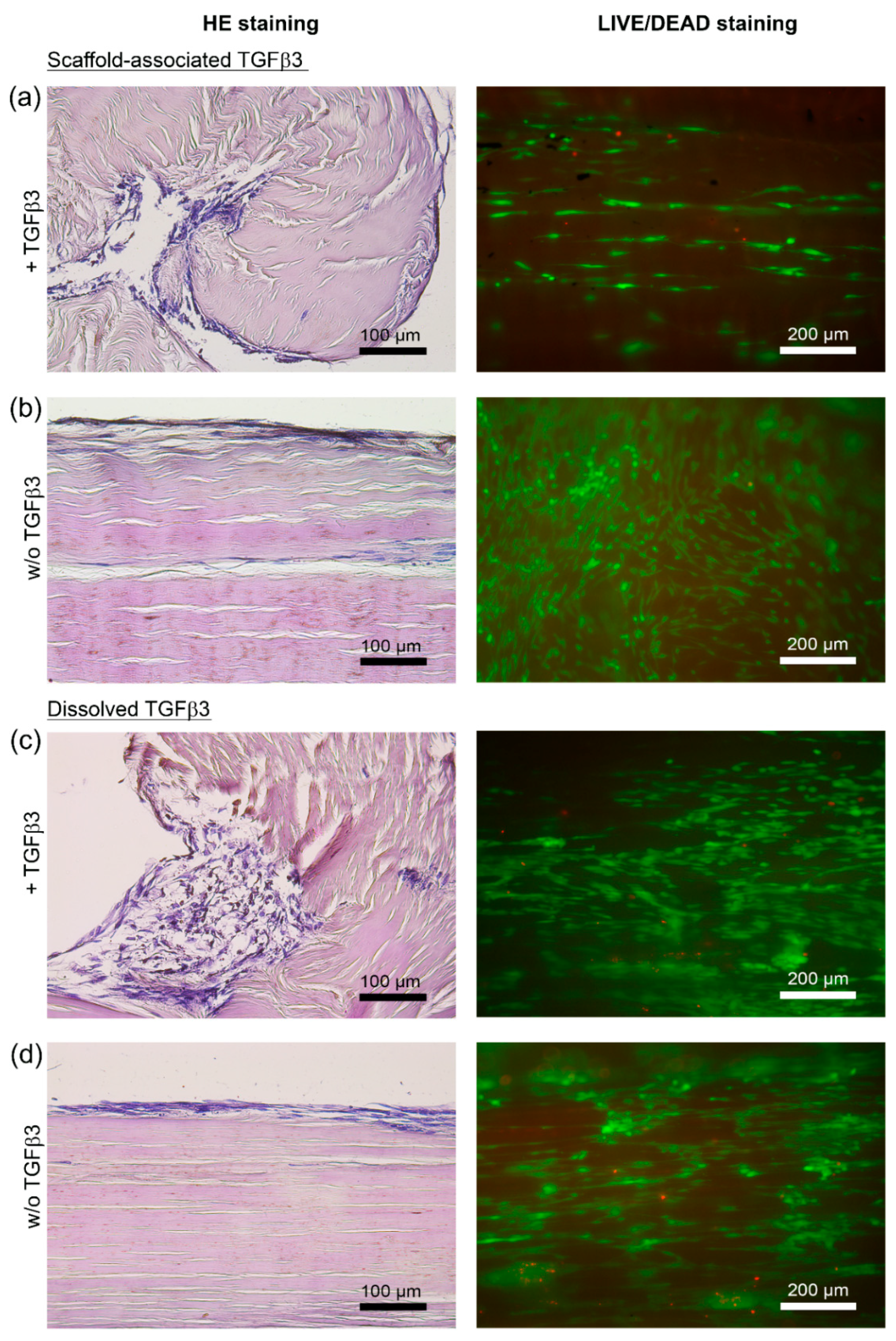
2.2.2. Cell Distribution and Cell Integration
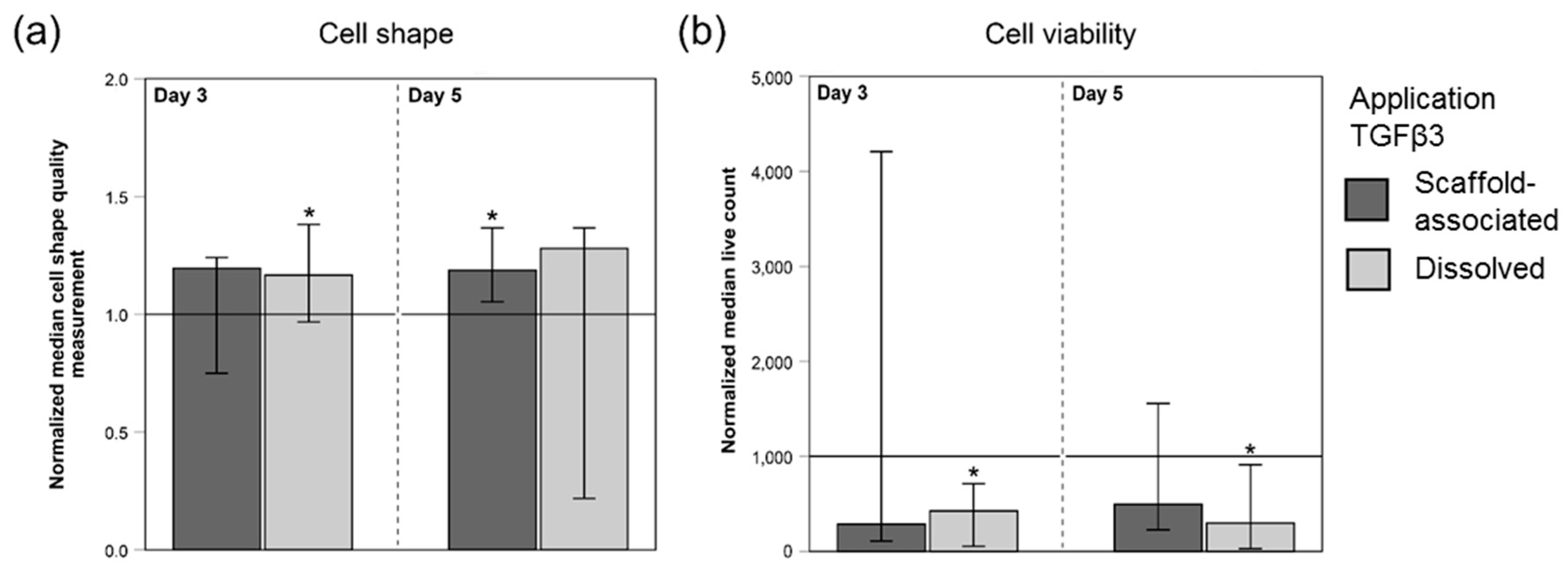
2.2.3. Cell Shape and Viability
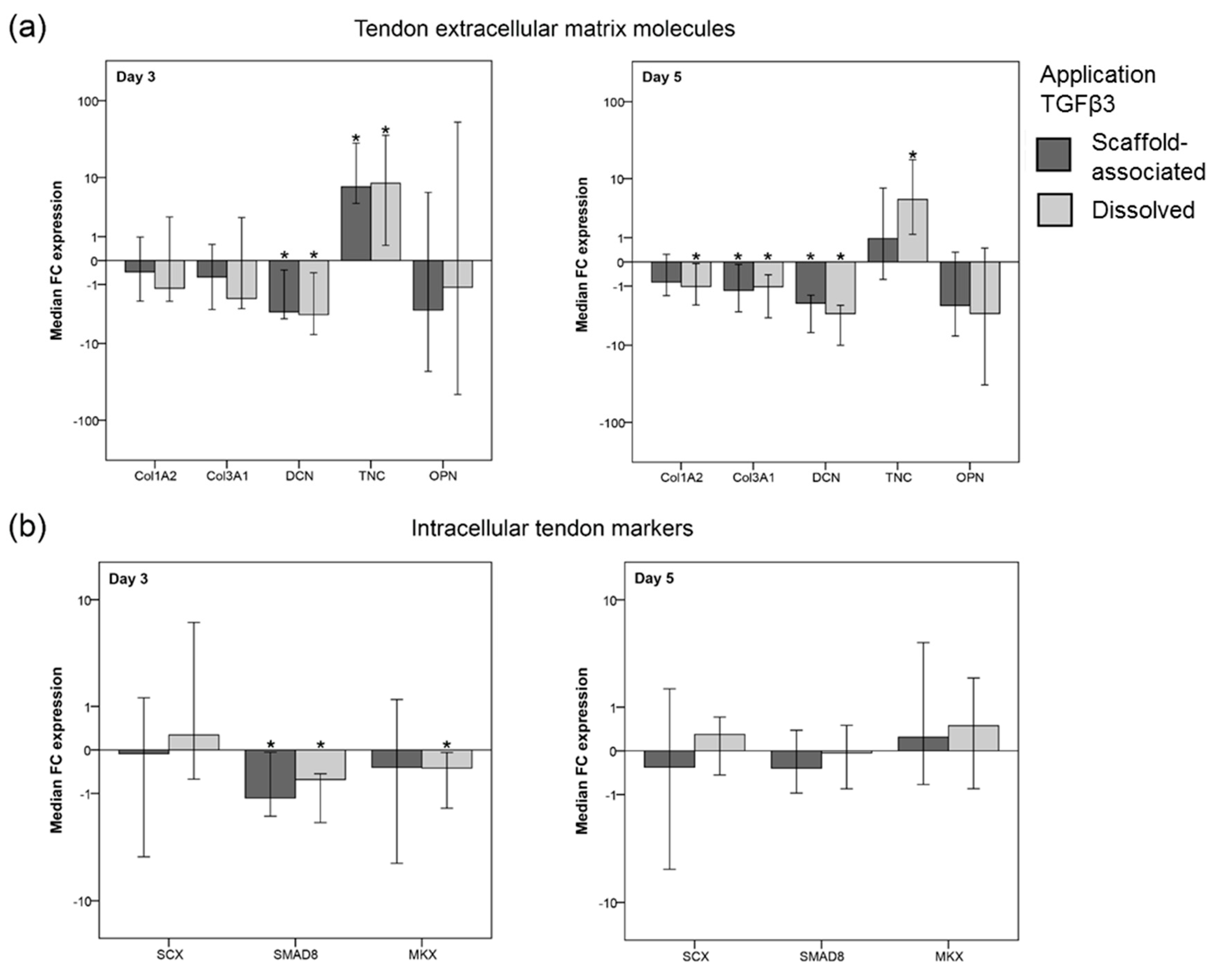
2.2.4. TGFβ3-Mediated Effects on the Gene Expression of ECM Molecules
2.2.5. TGFβ3-Mediated Effects on the Gene Expression of Intracellular Tendon Markers
3. Discussion
4. Materials and Methods
4.1. Tendon Scaffolds
4.2. TGFβ3 Dilutions and Scaffold Loading
4.3. Quantification of TGFβ3 by ELISA
4.4. MSC Culture and Scaffold Seeding
4.5. Macroscopic Assessment of the Scaffold Morphology
4.6. Histology
4.7. Real-Time PCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TGFβ3 | Transforming growth factor beta 3 |
| ECM | Extracellular matrix |
| MSC | Mesenchymal stromal cells |
| PBS | Phosphate buffered saline |
| BSA | Bovine serum albumin |
| FBS | Fetal bovine serum |
References
- Yang, G.; Rothrauff, B.B.; Tuan, R.S. Tendon and ligament regeneration and repair: Clinical relevance and developmental paradigm. Birth Defects Res. C Embryo Today Rev. 2013, 99, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Verdiyeva, G.; Koshy, K.; Glibbery, N.; Mann, H.; Seifalian, A.M. Tendon reconstruction with tissue engineering approach—A review. J. Biomed. Nanotechnol. 2015, 11, 1495–1523. [Google Scholar] [CrossRef]
- Siegel, L.; Vandenakker-Albanese, C.; Siegel, D. Anterior cruciate ligament injuries: Anatomy, physiology, biomechanics, and management. Clin. J. Sport Med. 2012, 22, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Tanzil, G.; Al-Sadi, O.; Ertel, W.; Lohan, A. Decellularized tendon extracellular matrix—A valuable approach for tendon reconstruction? Cells 2012, 1, 1010–1028. [Google Scholar] [CrossRef] [PubMed]
- Longo, U.G.; Lamberti, A.; Maffulli, N.; Denaro, V. Tissue engineered biological augmentation for tendon healing: A systematic review. Br. Med. Bull. 2011, 98, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Ladewig, K. Drug delivery in soft tissue engineering. Expert Opin. Drug Deliv. 2011, 8, 1175–1188. [Google Scholar] [CrossRef]
- Tessmar, J.K.; Gopferich, A.M. Matrices and scaffolds for protein delivery in tissue engineering. Adv. Drug Deliv. Rev. 2007, 59, 274–291. [Google Scholar] [CrossRef]
- Han, P.; Cui, Q.; Yang, S.; Wang, H.; Gao, P.; Li, Z. Tumor necrosis factor-alpha and transforming growth factor-beta1 facilitate differentiation and proliferation of tendon-derived stem cells in vitro. Biotechnol. Lett. 2017. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Lu, W.W.; Zhen, W.; Yang, D.; Peng, S. Novel biomaterial strategies for controlled growth factor delivery for biomedical applications. NPG Asia Mater. 2017, 9, e435. [Google Scholar] [CrossRef]
- Liu, Y.; Ramanath, H.S.; Wang, D.-A. Tendon tissue engineering using scaffold enhancing strategies. Trends Biotechnol. 2008, 26, 201–209. [Google Scholar] [CrossRef]
- Chung, H.J.; Park, T.G. Surface engineered and drug releasing pre-fabricated scaffolds for tissue engineering. Adv. Drug Deliv. Rev. 2007, 59, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Docheva, D.; Muller, S.A.; Majewski, M.; Evans, C.H. Biologics for tendon repair. Adv. Drug Deliv. Rev. 2015, 84, 222–239. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Yin, H.; Nerlich, M.; Pfeifer, C.G.; Docheva, D. Boosting tendon repair: Interplay of cells, growth factors and scaffold-free and gel-based carriers. J. Exp. Orthop. 2018, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-J.; Chen, X.; Li, G.; Chan, K.-M.; Heng, B.C.; Yin, Z.; Ouyang, H.-w. Concise Review: Stem Cell Fate Guided By Bioactive Molecules for Tendon Regeneration. Stem Cells Transl. Med. 2018, 7, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.K.; Petersen, B.C.; Tuan, R.S. Spatiotemporal protein distribution of TGF-betas, their receptors, and extracellular matrix molecules during embryonic tendon development. Dev. Dyn. 2008, 237, 1477–1489. [Google Scholar] [CrossRef]
- Chan, K.-M.; Fu, S.-C.; Wong, Y.-P.; Hui, W.-C.; Cheuk, Y.-C.; Wong, M.W.-N. Expression of transforming growth factor beta isoforms and their roles in tendon healing. Wound Repair Regen. 2008, 16, 399–407. [Google Scholar] [CrossRef]
- Berthet, E.; Chen, C.; Butcher, K.; Schneider, R.A.; Alliston, T.; Amirtharajah, M. Smad3 binds scleraxis and mohawk and regulates tendon matrix organization. J. Orthop. Res. 2013, 31, 1475–1483. [Google Scholar] [CrossRef]
- Havis, E.; Bonnin, M.-A.; Olivera-Martinez, I.; Nazaret, N.; Ruggiu, M.; Weibel, J.; Durand, C.; Guerquin, M.-J.; Bonod-Bidaud, C.; Ruggiero, F.; et al. Transcriptomic analysis of mouse limb tendon cells during development. Development 2014, 141, 3683–3696. [Google Scholar] [CrossRef]
- Pryce, B.A.; Watson, S.S.; Murchison, N.D.; Staverosky, J.A.; Dünker, N.; Schweitzer, R. Recruitment and maintenance of tendon progenitors by TGFbeta signaling are essential for tendon formation. Development 2009, 136, 1351–1361. [Google Scholar] [CrossRef]
- Barsby, T.; Guest, D. Transforming growth factor beta3 promotes tendon differentiation of equine embryo-derived stem cells. Tissue Eng. Part A 2013, 19, 2156–2165. [Google Scholar] [CrossRef]
- Kapacee, Z.; Yeung, C.-Y.C.; Lu, Y.; Crabtree, D.; Holmes, D.F.; Kadler, K.E. Synthesis of embryonic tendon-like tissue by human marrow stromal/mesenchymal stem cells requires a three-dimensional environment and transforming growth factor β3. Matrix Biol. 2010, 29, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Macri, L.; Silverstein, D.; Clark, R.A. Growth factor binding to the pericellular matrix and its importance in tissue engineering. Adv. Drug Deliv. Rev. 2007, 59, 1366–1381. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, J.; Samiric, T.; Ilic, M.Z.; Cook, J.; Handley, C.J. Involvement of proteoglycans in tendinopathy. J. Musculoskelet. Neuronal Interact. 2011, 11, 86–93. [Google Scholar] [PubMed]
- Prabhath, A.; Vernekar, V.N.; Sanchez, E.; Laurencin, C.T. Growth factor delivery strategies for rotator cuff repair and regeneration. Int. J. Pharm. 2018, 544, 358–371. [Google Scholar] [CrossRef]
- Moshaverinia, A.; Xu, X.; Chen, C.; Ansari, S.; Zadeh, H.H.; Snead, M.L.; Shi, S. Application of stem cells derived from the periodontal ligament or gingival tissue sources for tendon tissue regeneration. Biomaterials 2014, 35, 2642–2650. [Google Scholar] [CrossRef]
- Zhao, H.-Y.; Wu, J.; Zhu, J.-J.; Xiao, Z.-C.; He, C.-C.; Shi, H.-X.; Li, X.-K.; Yang, S.-L.; Xiao, J. Research advances in tissue engineering materials for sustained release of growth factors. BioMed Res. Int. 2015, 2015, 808202. [Google Scholar] [CrossRef]
- Lomas, A.J.; Ryan, C.N.M.; Sorushanova, A.; Shologu, N.; Sideri, A.I.; Tsioli, V.; Fthenakis, G.C.; Tzora, A.; Skoufos, I.; Quinlan, L.R.; et al. The past, present and future in scaffold-based tendon treatments. Adv. Drug Deliv. Rev. 2015, 84, 257–277. [Google Scholar] [CrossRef]
- Longo, U.G.; Lamberti, A.; Petrillo, S.; Maffulli, N.; Denaro, V. Scaffolds in tendon tissue engineering. Stem Cells Int. 2012, 2012, 517165. [Google Scholar] [CrossRef]
- Basile, P.; Dadali, T.; Jacobson, J.; Hasslund, S.; Ulrich-Vinther, M.; Søballe, K.; Nishio, Y.; Drissi, M.H.; Langstein, H.N.; Mitten, D.J.; et al. Freeze-dried tendon allografts as tissue-engineering scaffolds for Gdf5 gene delivery. Mol. Ther. 2008, 16, 466–473. [Google Scholar] [CrossRef]
- Abbah, S.A.; Spanoudes, K.; O’Brien, T.; Pandit, A.; Zeugolis, D.I. Assessment of stem cell carriers for tendon tissue engineering in pre-clinical models. Stem Cell Res. Ther. 2014, 5, 38. [Google Scholar] [CrossRef]
- Youngstrom, D.W.; Rajpar, I.; Kaplan, D.L.; Barrett, J.G. A bioreactor system for in vitro tendon differentiation and tendon tissue engineering. J. Orthop. Res. 2015, 33, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Burk, J.; Plenge, A.; Brehm, W.; Heller, S.; Pfeiffer, B.; Kasper, C. Induction of Tenogenic Differentiation Mediated by Extracellular Tendon Matrix and Short-Term Cyclic Stretching. Stem Cells Int. 2016, 2016, 7342379. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Chen, X.; Zhu, T.; Hu, J.J.; Song, H.X.; Shen, W.L.; Jiang, L.Y.; Heng, B.C.; Ji, J.F.; Ouyang, H.W. The effect of decellularized matrices on human tendon stem/progenitor cell differentiation and tendon repair. Acta Biomater. 2013, 9, 9317–9329. [Google Scholar] [CrossRef] [PubMed]
- Ning, L.J.; Zhang, Y.J.; Zhang, Y.; Qing, Q.; Jiang, Y.L.; Yang, J.L.; Luo, J.C.; Qin, T.W. The utilization of decellularized tendon slices to provide an inductive microenvironment for the proliferation and tenogenic differentiation of stem cells. Biomaterials 2015, 52, 539–550. [Google Scholar] [CrossRef]
- Qin, T.-W.; Chen, Q.; Sun, Y.-L.; Steinmann, S.P.; Amadio, P.C.; An, K.-N.; Zhao, C. Mechanical characteristics of native tendon slices for tissue engineering scaffold. J. Biomed. Mater. Res. B Appl. Biomater. 2012, 100, 752–758. [Google Scholar] [CrossRef]
- Almeida, H.V.; Liu, Y.; Cunniffe, G.M.; Mulhall, K.J.; Matsiko, A.; Buckley, C.T.; O’Brien, F.J.; Kelly, D.J. Controlled release of transforming growth factor-beta3 from cartilage-extra-cellular-matrix-derived scaffolds to promote chondrogenesis of human-joint-tissue-derived stem cells. Acta Biomater. 2014, 10, 4400–4409. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, S.H.; Jung, Y. TGF-beta3 encapsulated PLCL scaffold by a supercritical CO2-HFIP co-solvent system for cartilage tissue engineering. J. Control. Release 2015, 206, 101–107. [Google Scholar] [CrossRef]
- Yang, Q.; Teng, B.-H.; Wang, L.-N.; Li, K.; Xu, C.; Ma, X.-L.; Zhang, Y.; Kong, D.-L.; Wang, L.-Y.; Zhao, Y.-H. Silk fibroin/cartilage extracellular matrix scaffolds with sequential delivery of TGF-beta3 for chondrogenic differentiation of adipose-derived stem cells. Int. J. Nanomed. 2017, 12, 6721–6733. [Google Scholar] [CrossRef]
- Roth, S.P.; Schubert, S.; Scheibe, P.; Gross, C.; Brehm, W.; Burk, J. Growth Factor-Mediated Tenogenic Induction of Multipotent Mesenchymal Stromal Cells Is Altered by the Microenvironment of Tendon Matrix. Cell Transplant. 2018, 27, 1434–1450. [Google Scholar] [CrossRef]
- King, W.J.; Krebsbach, P.H. Growth factor delivery: How surface interactions modulate release in vitro and in vivo. Adv. Drug Deliv. Rev. 2012, 64, 1239–1256. [Google Scholar] [CrossRef]
- Rifkin, D.B. Latent transforming growth factor-beta (TGF-beta) binding proteins: Orchestrators of TGF-beta availability. J. Biol. Chem. 2005, 280, 7409–7412. [Google Scholar] [CrossRef] [PubMed]
- Munger, J.S.; Sheppard, D. Cross talk among TGF-beta signaling pathways, integrins, and the extracellular matrix. Cold Spring Harb. Perspect. Biol. 2011, 3, a005017. [Google Scholar] [CrossRef] [PubMed]
- Danielpour, D.; Roberts, A.B. Specific and sensitive quantitation of transforming growth factor beta 3 by sandwich enzyme-linked immunosorbent assay. J. Immunol. Methods 1995, 180, 265–272. [Google Scholar] [CrossRef]
- Taylor, S.E.; Vaughan-Thomas, A.; Clements, D.N.; Pinchbeck, G.; Macrory, L.C.; Smith, R.K.W.; Clegg, P.D. Gene expression markers of tendon fibroblasts in normal and diseased tissue compared to monolayer and three dimensional culture systems. BMC Musculoskelet. Disord. 2009, 10, 27. [Google Scholar] [CrossRef]
- Chiquet-Ehrismann, R. Tenascins. Int. J. Biochem. Cell Biol. 2004, 36, 986–990. [Google Scholar] [CrossRef]
- Lu, P.; Zhang, G.-R.; Cai, Y.-Z.; Heng, B.C.; Ren, H.; Wang, L.-L.; Ji, J.; Zou, X.-H.; Ouyang, H.W. Lentiviral-encoded shRNA silencing of proteoglycan decorin enhances tendon repair and regeneration within a rat model. Cell Transplant. 2013, 22, 1507–1517. [Google Scholar] [CrossRef]
- Nakamura, N.; Hart, D.A.; Boorman, R.S.; Kaneda, Y.; Shrive, N.G.; Marchuk, L.L.; Shino, K.; Ochi, T.; Frank, C.B. Decorin antisense gene therapy improves functional healing of early rabbit ligament scar with enhanced collagen fibrillogenesis in vivo. J. Orthop. Res. 2000, 18, 517–523. [Google Scholar] [CrossRef]
- Hoffmann, A.; Pelled, G.; Turgeman, G.; Eberle, P.; Zilberman, Y.; Shinar, H.; Keinan-Adamsky, K.; Winkel, A.; Shahab, S.; Navon, G.; et al. Neotendon formation induced by manipulation of the Smad8 signalling pathway in mesenchymal stem cells. J. Clin. Investig. 2006, 116, 940–952. [Google Scholar] [CrossRef]
- Shahab-Osterloh, S.; Witte, F.; Hoffmann, A.; Winkel, A.; Laggies, S.; Neumann, B.; Seiffart, V.; Lindenmaier, W.; Gruber, A.D.; Ringe, J.; et al. Mesenchymal stem cell-dependent formation of heterotopic tendon-bone insertions (osteotendinous junctions). Stem Cells 2010, 28, 1590–1601. [Google Scholar] [CrossRef]
- Pelled, G.; Snedeker, J.G.; Ben-Arav, A.; Rigozzi, S.; Zilberman, Y.; Kimelman-Bleich, N.; Gazit, Z.; Muller, R.; Gazit, D. Smad8/BMP2-engineered mesenchymal stem cells induce accelerated recovery of the biomechanical properties of the Achilles tendon. J. Orthop. Res. 2012, 30, 1932–1939. [Google Scholar] [CrossRef]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-beta and the TGF-beta family: Context-dependent roles in cell and tissue physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef] [PubMed]
- Ivaska, J.; Heino, J. Cooperation between integrins and growth factor receptors in signaling and endocytosis. Annu. Rev. Cell Dev. Biol. 2011, 27, 291–320. [Google Scholar] [CrossRef] [PubMed]
- Lovati, A.B.; Bottagisio, M.; Moretti, M. Decellularized and Engineered Tendons as Biological Substitutes: A Critical Review. Stem Cells Int. 2016, 2016, 7276150. [Google Scholar] [CrossRef]
- Patterson-Kane, J.C.; Rich, T. Achilles tendon injuries in elite athletes: Lessons in pathophysiology from their equine counterparts. ILAR J. 2014, 55, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Nerlich, M.; Docheva, D. Tendon injuries: Basic science and new repair proposals. EFORT Open Rev. 2017, 2, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.T.; Reis, R.L.; Gomes, M.E. Engineering tendon and ligament tissues: Present developments towards successful clinical products. J. Tissue Eng. Regen. Med. 2013, 7, 673–686. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.P.; Erbe, I.; Burk, J. Decellularization of Large Tendon Specimens: Combination of Manually Performed Freeze-Thaw Cycles and Detergent Treatment. Methods Mol. Biol. 2017. [Google Scholar] [CrossRef]
- Burk, J.; Erbe, I.; Berner, D.; Kacza, J.; Kasper, C.; Pfeiffer, B.; Winter, K.; Brehm, W. Freeze-thaw cycles enhance decellularization of large tendons. Tissue Eng. Part. C Methods 2014, 20, 276–284. [Google Scholar] [CrossRef]
- Ryan, J.A. Evolution of Cell Culture Surfaces. Available online: http://www.sigmaaldrich.com/china-mainland/zh/technical-documents/articles/biofiles/evolution-of-cell.html (accessed on 11 August 2017).
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control TGFβ3 Solution | Rinsing Fluid after Scaffold Loading | |||
|---|---|---|---|---|
| Ultra-Low Attachment Dish (n = 7) | Standard Dish (n = 2) | Full-Thickness Scaffold (n = 2) | 0.3 mm Scaffold (n = 3) | |
| 5 ng TGFβ3: | ||||
| OD (mean ± SD) | 1.325 ± 0.322 | 0.426 ± 0.066 | 0.08 ± 0.005 | 0.139 ± 0.022 |
| Dissolved TGFβ3 (%) | 100 ± 24 | 32 ± 5 | 6 ± 0.4 | 10 ± 2 |
| Scaffold-Associated TGFβ3 (%) | 68 ± 5 | 94 ± 0.4 | 90 ± 2 | |
| 10 ng TGFβ3: | ||||
| OD (mean ± SD) | 2.391 ± 0.454 | 1.416 ± 0.000 | 0.194 ± 0.011 | 0.292 ± 0.050 |
| Dissolved TGFβ3 (%) | 100 ± 19 | 59 ± 0 | 8 ± 0.5 | 12 ± 2 |
| Scaffold-Associated TGFβ3 (%) | 41 ± 0 | 92 ± 0.5 | 88 ± 2 | |
| Equine Gene | Primer Pair Sequences | Accession Number | PCR Product in bp |
|---|---|---|---|
| Beta-actin | For: ATCCACGAAACTACCTTCAAC Rev: CGCAATGATCTTGATCTTCATC | NM_001081838.1 | 174 |
| GAPDH | For: TGGAGAAAGCTGCCAAATACG Rev: GGCCTTTCTCCTTCTCTTGC | NM_001163856.1 | 309 |
| Collagen 1A2 | For: CAACCGGAGATAGAGGACCA Rev: CAGGTCCTTGGAAACCTTGA] | XM_001492939.3 | 243] |
| Collagen 2A1 | For: ATTGTAGGACCCAAAGGACC Rev: CAGCAAAGTTTCCACCAAGG | NM_001081764.1 | 199 |
| Collagen 3A1 | For: AGGGGACCTGGTTACTGCTT Rev: TCTCTGGGTTGGGACAGTCT | XM_001917620.3 | 216 |
| Scleraxis | For: TACCTGGGTTTTCTTCTGGTCACT Rev: TATCAAAGACACAAGATGCCAGC | NM_001105150.1 | 51 |
| Osteopontin | For: TGAAGACCAGTATCCTGATGC Rev: GCTGACTTGTTTCCTGACTG | XM_001496152.3 | 158 |
| Decorin | For: ACCCACTGAAGAGCTCAGGA Rev: GCCATTGTCAACAGCAGAGA | NM_001081925.2 | 239 |
| Tenascin c | For: TCACATCCAGGTGCTTATTCC Rev: CTAGAGTGTCTCACTATCAGG | XM_001916622.3 | 163 |
| Mohawk | For: AAGATACTCTTGGCGCTCGG Rev: ACACTAAGCCGCTCAGCA | XM_014737017.1 | 170 |
| Smad8 | For: AGCCTCCGTGCTCTGCATT Rev: CCCAACTCGGTTGTTTAGTTCAT | AB106117.1 | 200 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roth, S.P.; Brehm, W.; Groß, C.; Scheibe, P.; Schubert, S.; Burk, J. Transforming Growth Factor Beta 3-Loaded Decellularized Equine Tendon Matrix for Orthopedic Tissue Engineering. Int. J. Mol. Sci. 2019, 20, 5474. https://doi.org/10.3390/ijms20215474
Roth SP, Brehm W, Groß C, Scheibe P, Schubert S, Burk J. Transforming Growth Factor Beta 3-Loaded Decellularized Equine Tendon Matrix for Orthopedic Tissue Engineering. International Journal of Molecular Sciences. 2019; 20(21):5474. https://doi.org/10.3390/ijms20215474
Chicago/Turabian StyleRoth, Susanne Pauline, Walter Brehm, Claudia Groß, Patrick Scheibe, Susanna Schubert, and Janina Burk. 2019. "Transforming Growth Factor Beta 3-Loaded Decellularized Equine Tendon Matrix for Orthopedic Tissue Engineering" International Journal of Molecular Sciences 20, no. 21: 5474. https://doi.org/10.3390/ijms20215474
APA StyleRoth, S. P., Brehm, W., Groß, C., Scheibe, P., Schubert, S., & Burk, J. (2019). Transforming Growth Factor Beta 3-Loaded Decellularized Equine Tendon Matrix for Orthopedic Tissue Engineering. International Journal of Molecular Sciences, 20(21), 5474. https://doi.org/10.3390/ijms20215474