Trypanosoma brucei J-Protein 2 Functionally Co-Operates with the Cytosolic Hsp70 and Hsp70.4 Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
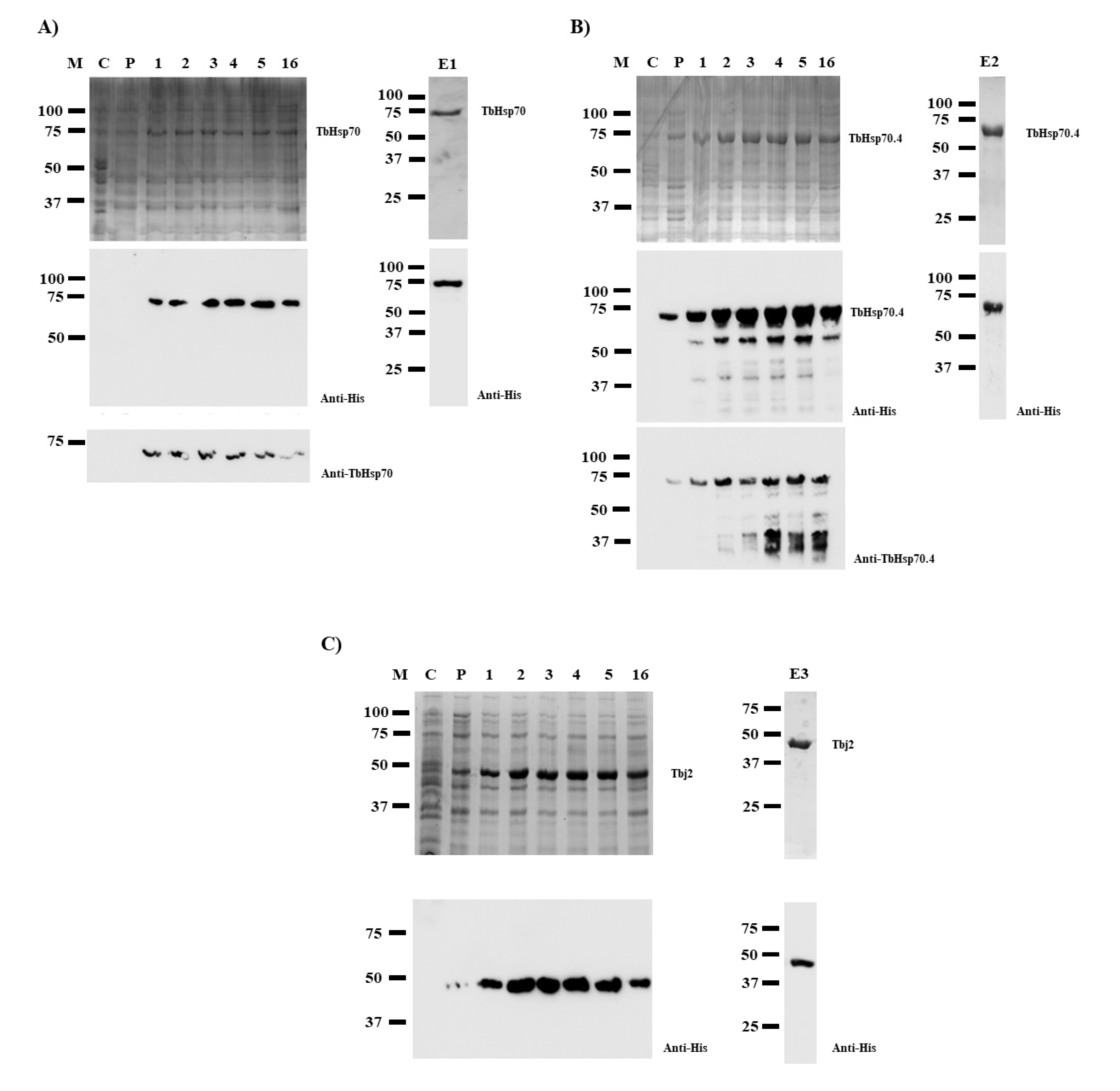
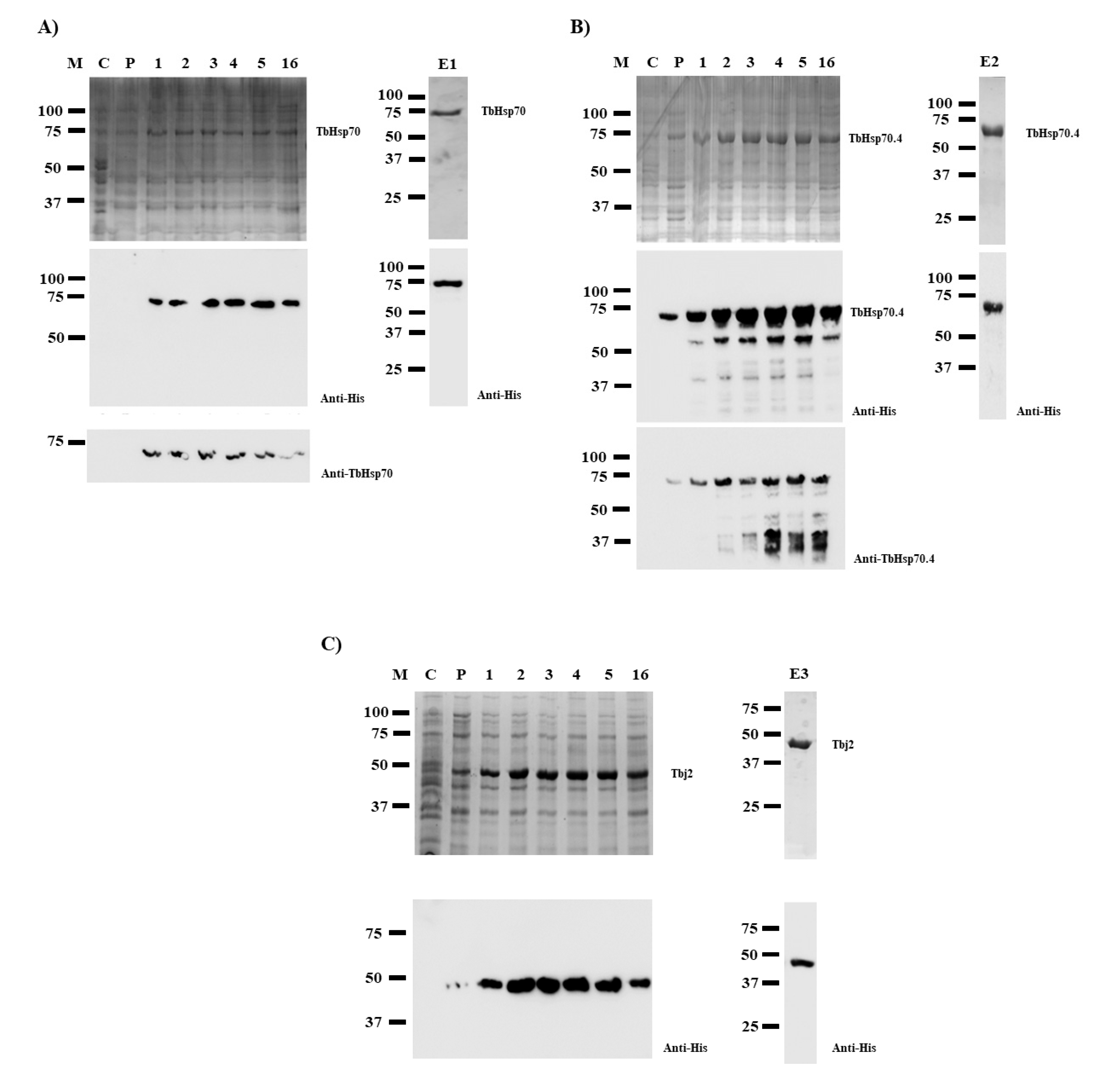
2.1. Recombinant Protein Production and Purification
2.2. Protein Expression of TbHsp70 and TbHsp70.4 Is Modulated in Response to Heat Stress
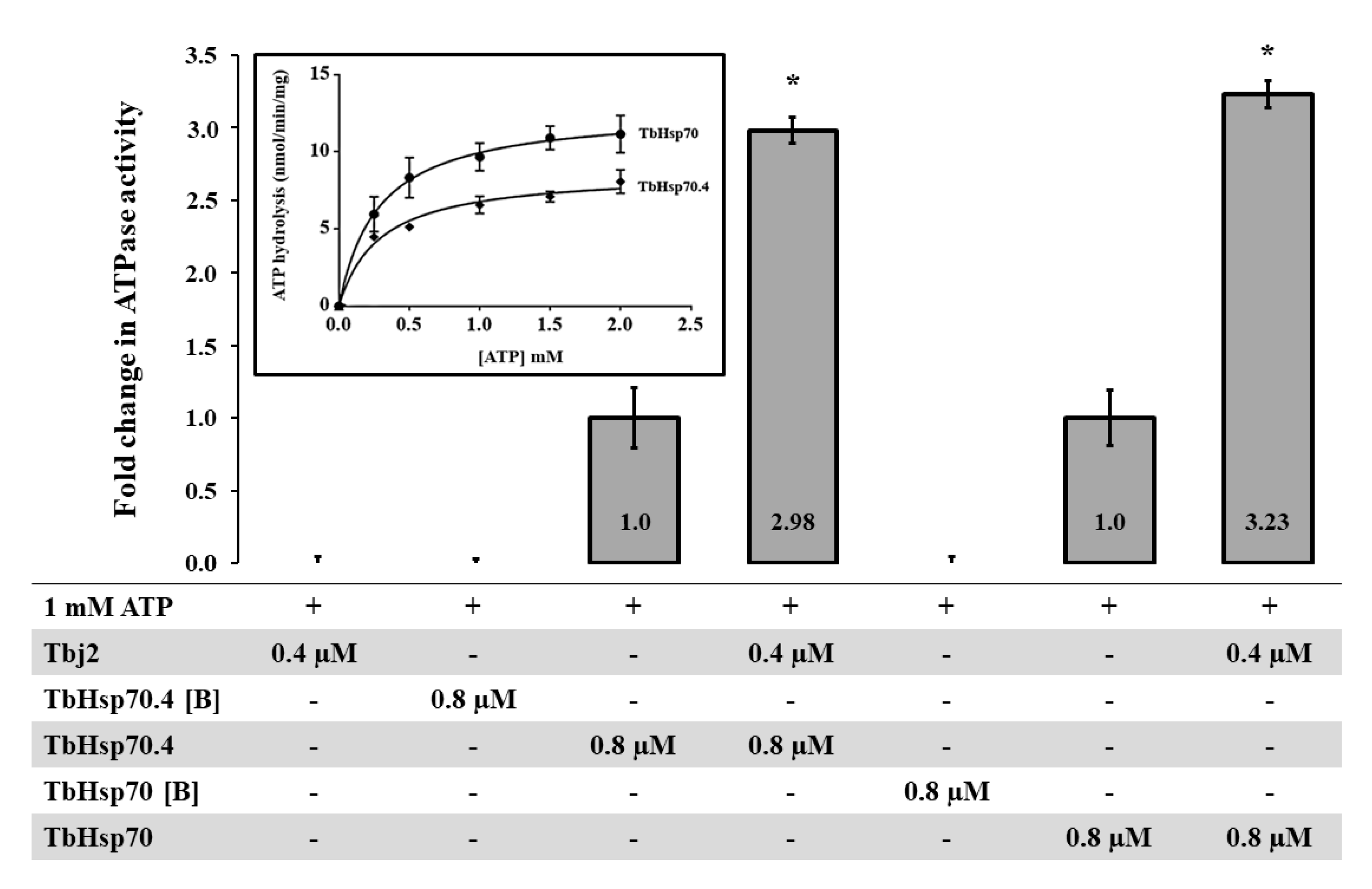
2.3. TbHsp70 and TbHsp70.4 Both Possess Intrinsic ATPase Activity, Which Is Stimulated by Tbj2
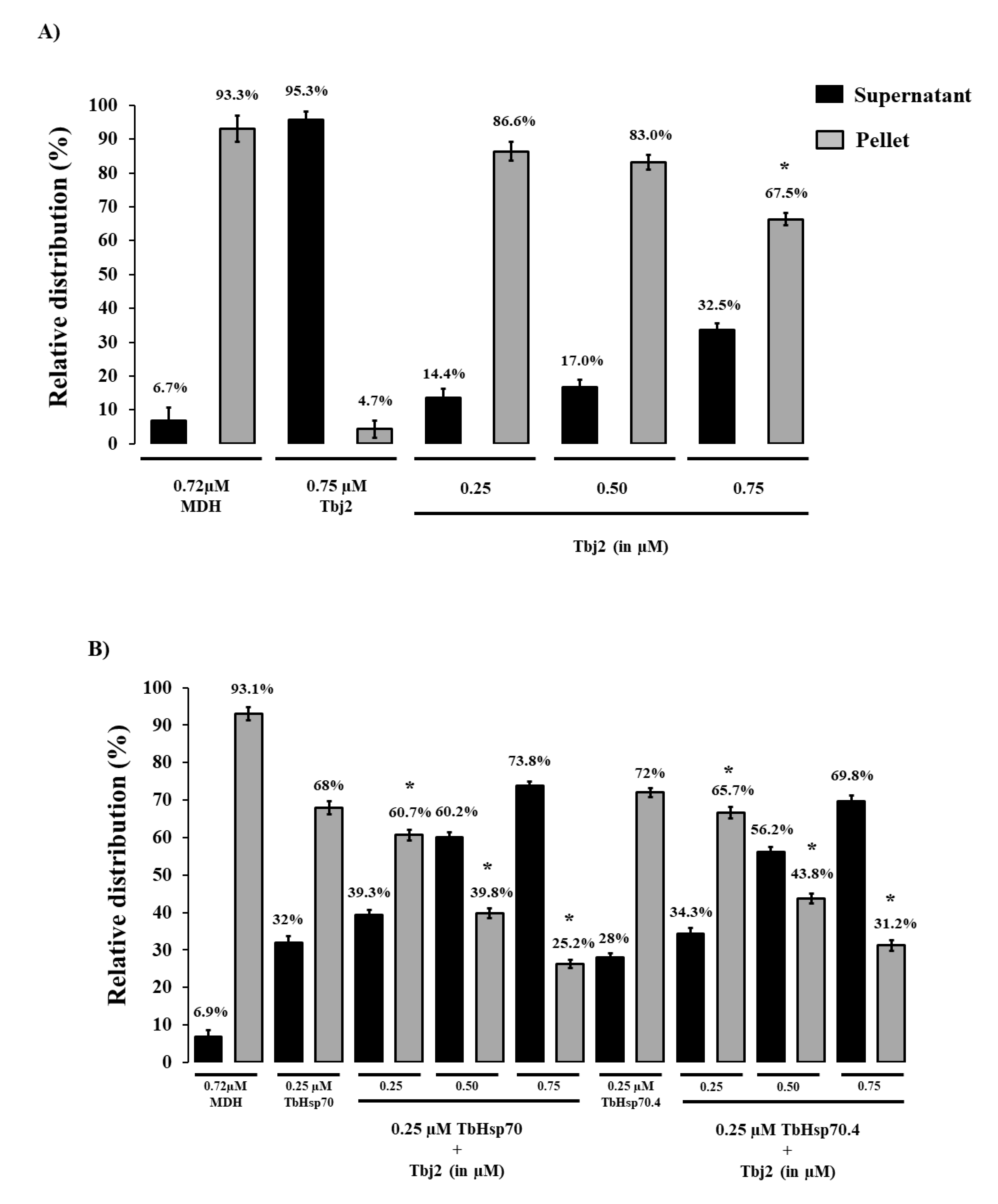
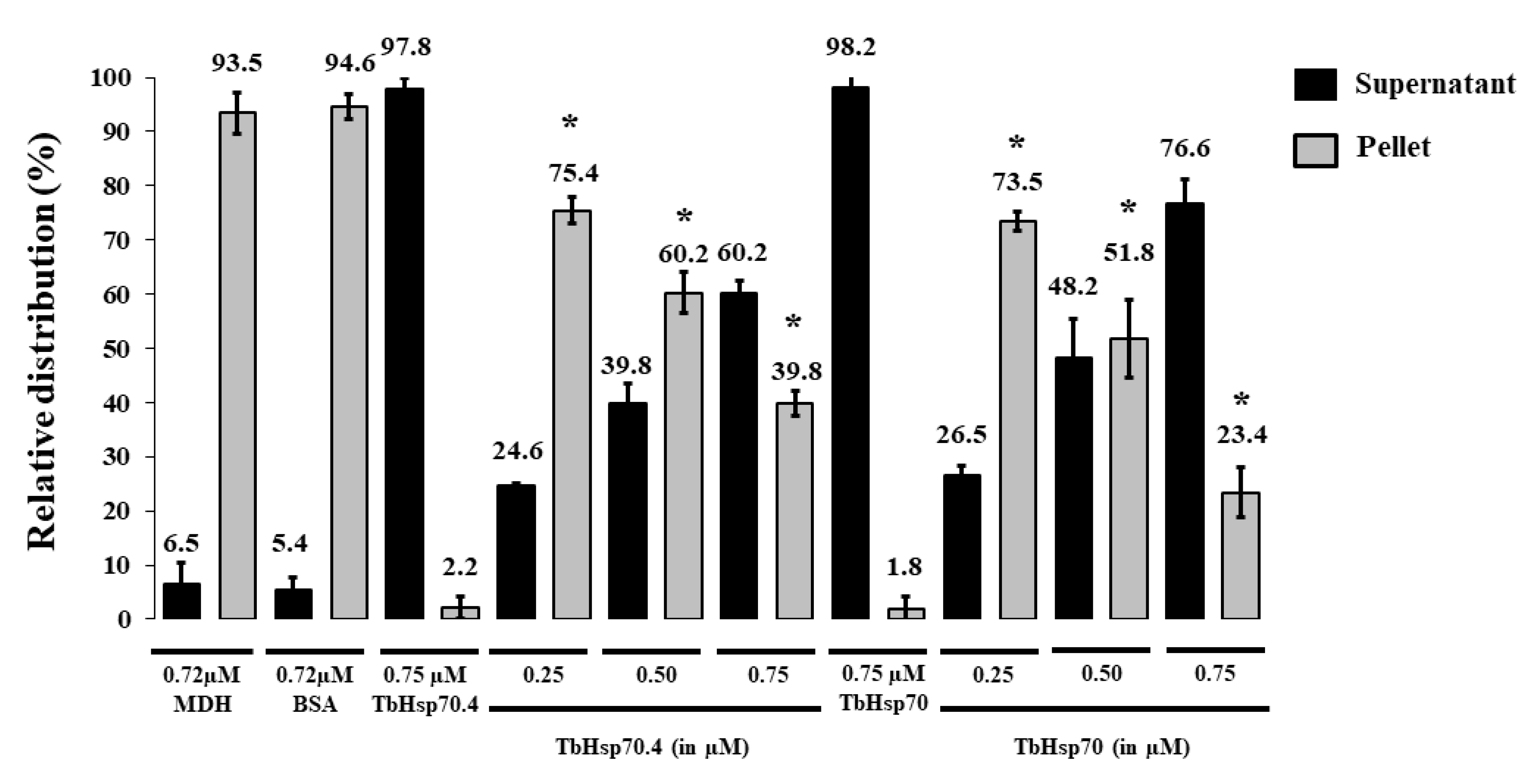
2.4. TbHsp70 and TbHsp70.4 Both Suppress the Thermal Aggregation of Malate Dehydrogenase (MDH)
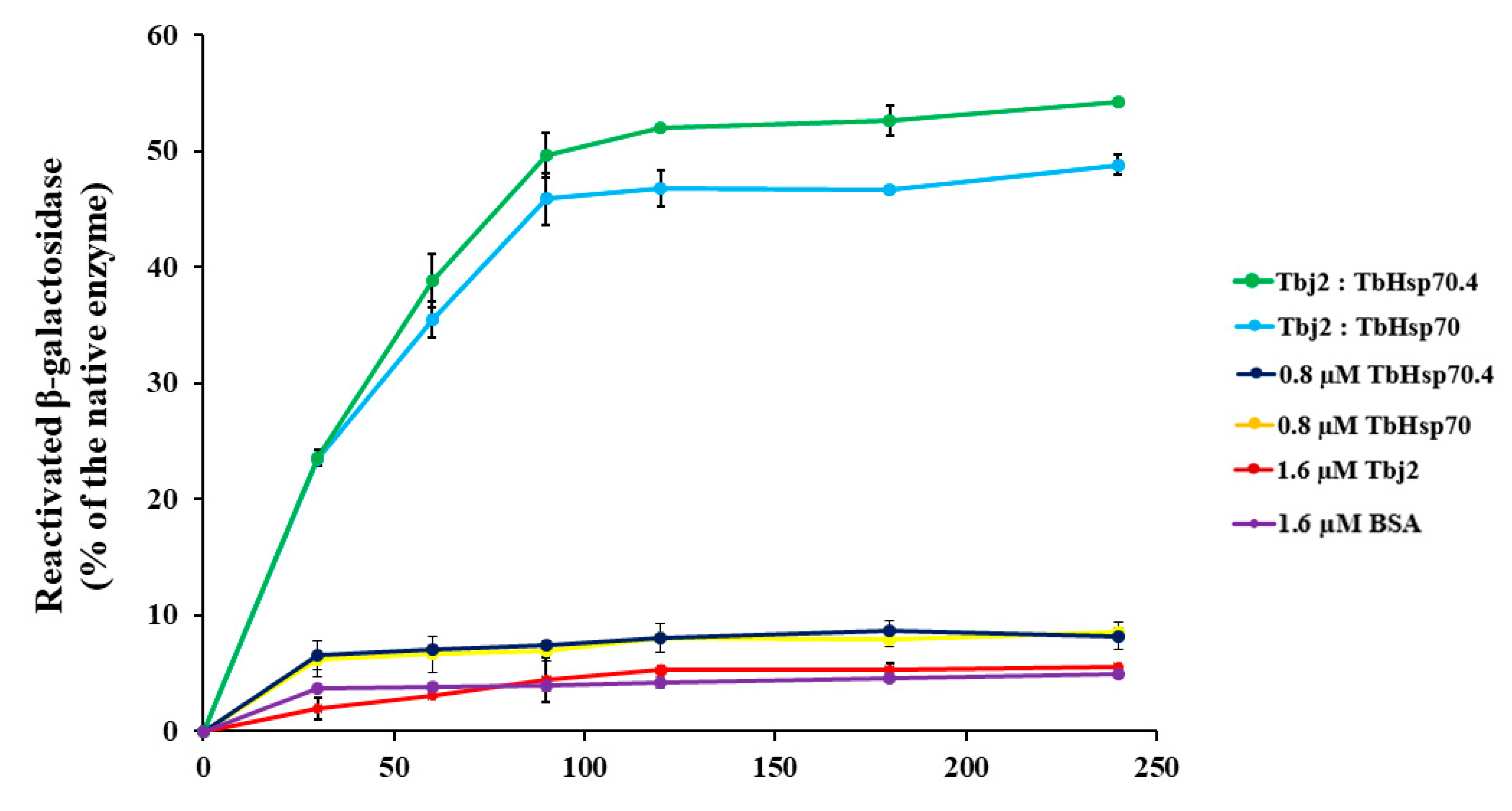
2.5. Tbj2 Stimulates the Refoldase Activities of TbHsp70 and TbHsp70.4
3. Materials and Methods
3.1. The pQE2/TbHsp70 Expression Vector
3.2. Heterologous Expression and Purification of TbHsp70 and TbHsp70.4
3.3. Purification of Tbj2
3.4. Investigation of Heat-Induced Expression of TbHsp70 and TbHsp70.4 in T. b. brucei Lister 927 V221 Parasites
3.5. MDH Aggregation Suppression Assay
3.6. ATPase Activity Assay
3.7. β-galactosidase Refolding Assay
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Alsan, M. The effect of the tsetse fly on African development. Am. Econ. Rev. 2015, 105, 382–410. [Google Scholar] [CrossRef]
- Barrett, M.P.; Croft, S.L. Management of trypanosomiasis and leishmaniasis. Br. Med. Bull. 2012, 104, 175–196. [Google Scholar] [CrossRef] [PubMed]
- Shonhai, A.; Maier, A.G.; Przyborski, J.M.; Blatch, G.L. Intracellular protozoan parasites of humans: The role of molecular chaperones in development and pathogenesis. Protein Pept. Lett. 2011, 18, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Requena, J.M.; Montalvo, A.M.; Fraga, J. Molecular chaperones of Leishmania: Central players in many stress-related and-unrelated physiological processes. BioMed Res. Int. 2015, 2015, 301326. [Google Scholar] [CrossRef] [PubMed]
- Urmenyi, T.P.; Silva, R.; Rondinelli, E. The heat shock proteins of Trypanosoma cruzi. In Proteins and Proteomics of Leishmania and Trypanosoma; Santos, A., Branquinha, M., d’Avila-Levy, C., Kneipp, L., Sodré, C., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 74, pp. 119–135. [Google Scholar]
- Bentley, S.J.; Jamabo, M.; Boshoff, A. The Hsp70/J-protein machinery of the African trypanosome, Trypanosoma brucei. Cell Stress Chaperones 2019, 24, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Bukau, B.; Weissman, J.; Horwich, A. Molecular chaperones and protein quality control. Cell 2006, 125, 443–451. [Google Scholar] [CrossRef]
- Daugaard, M.; Rohde, M.; Jäättelä, M. The heat shock protein 70 family: Highly homologous proteins with overlapping and distinct functions. FEBS Lett. 2007, 581, 3702–3710. [Google Scholar] [CrossRef]
- Brocchieri, L.; de Macario, E.C.; Macario, A.J.L. Hsp70 genes in the human genome: Conservation and differentiation patterns predict a wide array of overlapping and specialized functions. BMC Evol. Biol. 2008, 8, 19. [Google Scholar] [CrossRef]
- Flaherty, K.M.; Deluca-Flaherty, C.; McKay, D.B. Three-dimensional structure of the ATPase fragment of a 70K heat shock cognate protein. Nature 1990, 346, 623–628. [Google Scholar] [CrossRef]
- Wang, T.F.; Chang, J.H.; Wang, C. Identification of the peptide binding domain of hsc70. J. Biol. Chem. 1993, 268, 26049–26051. [Google Scholar] [PubMed]
- Slepenkov, S.V.; Witt, S.N. The unfolding story of the Escherichia coli Hsp70 DnaK: Is DnaK a holdase or an unfoldase? Mol. Microbiol. 2002, 45, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Clerico, E.M.; Tilitsky, J.M.; Meng, W.; Gierasch, L.M. How hsp70 molecular machines interact with their substrates to mediate diverse physiological functions. J. Mol. Biol. 2015, 427, 1575–1588. [Google Scholar] [CrossRef] [PubMed]
- Meimaridou, E.; Gooljar, S.B.; Chapple, J.P. From hatching to dispatching: The multiple cellular roles of the Hsp70 molecular chaperone machinery. J. Mol. Endocrinol. 2009, 42, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, F.; Nicoll, W.S.; Zimmermann, R.; Cheetham, M.E.; Blatch, G.L. Not all J domains are created equal: Implications for the specificity of Hsp40–Hsp70 interactions. Protein Sci. 2005, 14, 1697–1709. [Google Scholar] [CrossRef]
- Cheetham, M.E.; Caplan, A.J. Structure, function and evolution of DnaJ: Conservation and adaptation of chaperone function. Cell Stress Chaperones 1998, 3, 28–36. [Google Scholar] [CrossRef]
- Botha, M.; Pesce, E.R.; Blatch, G.L. The Hsp40 proteins of Plasmodium falciparum and other apicomplexa: Regulating chaperone power in the parasite and the host. Int. J. Biochem. Cell Biol. 2007, 39, 1781–1803. [Google Scholar] [CrossRef]
- Alsford, S.; Turner, D.J.; Obado, S.O.; Sanchez-Flores, A.; Glover, L.; Berriman, M.; Hertz-Fowler, C.; Horn, D. High-throughput phenotyping using parallel sequencing of RNA interference targets in the African trypanosome. Genome Res. 2011, 21, 915–924. [Google Scholar] [CrossRef]
- Miller, M.A.; McGowan, S.E.; Gantt, K.R.; Champion, M.; Novick, S.L.; Andersen, K.A.; Bacchi, C.J.; Yarlett, N.; Britigan, B.E.; Wilson, M.E. Inducible resistance to oxidant stress in the protozoan Leishmania chagasi. J. Biol. Chem. 2000, 275, 33883–33889. [Google Scholar] [CrossRef]
- Maharjan, M.; Madhubala, R. Heat shock protein 70 (HSP70) expression in antimony susceptible/resistant clinical isolates of Leishmania donovani. Nepal J. Biotechnol. 2015, 3, 22–28. [Google Scholar] [CrossRef]
- Codonho, B.S.; Costa, S.; Peloso Ede, F.; Joazeiro, P.P.; Gadelha, F.R.; Giorgio, S. HSP70 of Leishmania amazonensis alters resistance to different stresses and mitochondrial bioenergetics. Mem. Inst. Oswaldo Cruz. 2016, 111, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Searle, S.; Smith, D.F. Leishmania major: Characterisation and expression of a cytoplasmic stress-related protein. Exp. Parasitol. 1993, 77, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Ludewig, M.H.; Boshoff, A.; Horn, D.; Blatch, G.L. Trypanosoma brucei J protein 2 is a stress inducible and essential Hsp40. Int. J. Biochem. Cell Biol. 2015, 60, 93–98. [Google Scholar] [CrossRef]
- Burger, A.; Ludewig, M.L.; Boshoff, A. Investigating the chaperone properties of a novel heat shock protein, Hsp70.c, from Trypanosoma brucei. J. Parasitol. Res. 2014, 2014, 172582. [Google Scholar] [CrossRef] [PubMed]
- Fenn, K.; Matthews, K.R. The cell biology of Trypanosoma brucei differentiation. Curr. Opin. Microbiol. 2007, 10, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Snoeckx, L.H.; Cornelussen, R.N.; Van Nieuwenhoven, F.A.; Reneman, R.S.; Van Der Vusse, G.J. Heat shock proteins and cardiovascular pathophysiology. Physiol. Rev. 2001, 81, 1461–1497. [Google Scholar] [CrossRef]
- Häusler, T.; Clayton, C. Post-transcriptional control of hsp70 mRNA in Trypanosoma brucei. Mol. Biochem. Parasitol. 1996, 76, 57–71. [Google Scholar] [CrossRef]
- Van der Ploeg, L.H.; Giannini, S.H.; Cantor, C.R. Heat shock genes: Regulatory role for differentiation in parasitic protozoa. Science 1985, 228, 1443–1446. [Google Scholar] [CrossRef]
- de Carvalho, E.F.; de Castro, F.T.; Rondinelli, E.; Soares, C.M.; Carvalho, J.F. HSP70 gene expression in Trypanosoma cruzi is regulated at different levels. J. Cell Physiol. 1990, 143, 439–444. [Google Scholar] [CrossRef]
- Vos, M.J.; Hageman, J.; Carra, S.; Kampinga, H.H. Structural and functional diversities between members of the human HSPB, HSPH, HSPA, and DNAJ chaperone families. Biochemistry 2008, 47, 7001–7011. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Craig, E.A. The HSP70 chaperone machinery: J proteins as drivers of functional specificity. Nat. Rev. Mol. Cell Biol. 2010, 11, 579–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, S.; Sunter, J.D.; Wheeler, R.J. TrypTag.org: A trypanosome genome-wide protein localisation resource. Trends Parasitol. 2017, 33, 80–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edkins, A.L.; Ludewig, M.H.; Blatch, G.L. A Trypanosoma cruzi heat shock protein 40 is able to stimulate the adenosine triphosphate hydrolysis activity of heat shock protein 70 and can substitute for a yeast heat shock protein 40. Int. J. Biochem. Cell Biol. 2004, 36, 1585–1598. [Google Scholar] [CrossRef] [PubMed]
- Zininga, T.; Pooe, O.J.; Makhado, P.B.; Ramatsui, L.; Prinsloo, E.; Achilonu, I.; Dirr, H.; Shonhai, A. Polymyxin B inhibits the chaperone activity of Plasmodium falciparum Hsp70. Cell Stress Chaperones 2017, 22, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Cyr, D.M.; Ramos, C.H. Specification of Hsp70 function by Type I and Type II Hsp40. Subcell. Biochem. 2015, 78, 91–102. [Google Scholar]
- Freeman, B.C.; Morimoto, R.I. The human cytosolic molecular chaperones hsp90, hsp70 (hsc70) and hdj-1 have distinct roles in recognition of a non-native protein and protein refolding. EMBO J. 1996, 15, 2969–2979. [Google Scholar] [CrossRef]
- Johnson, B.D.; Schumacher, R.J.; Ross, E.D. Hop modulates Hsp70/Hsp90 interactions in protein folding. J. Biol. Chem. 1998, 273, 3679–3686. [Google Scholar] [CrossRef] [Green Version]
- Bangs, J.D.; Uyetake, L.; Brickman, M.; Balber, A.E.; Boothroyd, J.C. Molecular cloning and cellular localization of a BiP homologue in Trypanosoma brucei: Divergent ER retention signals in a lower eukaryote. J. Cell Sci. 1993, 105, 1101–1113. [Google Scholar]
- Bangs, J.D.; Brouch, E.M.; Ransom, D.M.; Roggy, J.L. A soluble secretory reporter system in Trypanosoma brucei: Studies on endoplasmic reticulum targeting. J. Biol. Chem. 1996, 271, 18387–18393. [Google Scholar] [CrossRef] [Green Version]
- Rotman, B. Regulation of enzymatic activity in the intact cell: The beta-D-galactosidase of Escherichia coli. J. Bacteriol. 1958, 76, 1–14. [Google Scholar]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bentley, S.J.; Boshoff, A. Trypanosoma brucei J-Protein 2 Functionally Co-Operates with the Cytosolic Hsp70 and Hsp70.4 Proteins. Int. J. Mol. Sci. 2019, 20, 5843. https://doi.org/10.3390/ijms20235843
Bentley SJ, Boshoff A. Trypanosoma brucei J-Protein 2 Functionally Co-Operates with the Cytosolic Hsp70 and Hsp70.4 Proteins. International Journal of Molecular Sciences. 2019; 20(23):5843. https://doi.org/10.3390/ijms20235843
Chicago/Turabian StyleBentley, Stephen John, and Aileen Boshoff. 2019. "Trypanosoma brucei J-Protein 2 Functionally Co-Operates with the Cytosolic Hsp70 and Hsp70.4 Proteins" International Journal of Molecular Sciences 20, no. 23: 5843. https://doi.org/10.3390/ijms20235843