Transcriptomic Analysis of Stem Cells Treated with Moringin or Cannabidiol: Analogies and Differences in Inflammation Pathways

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
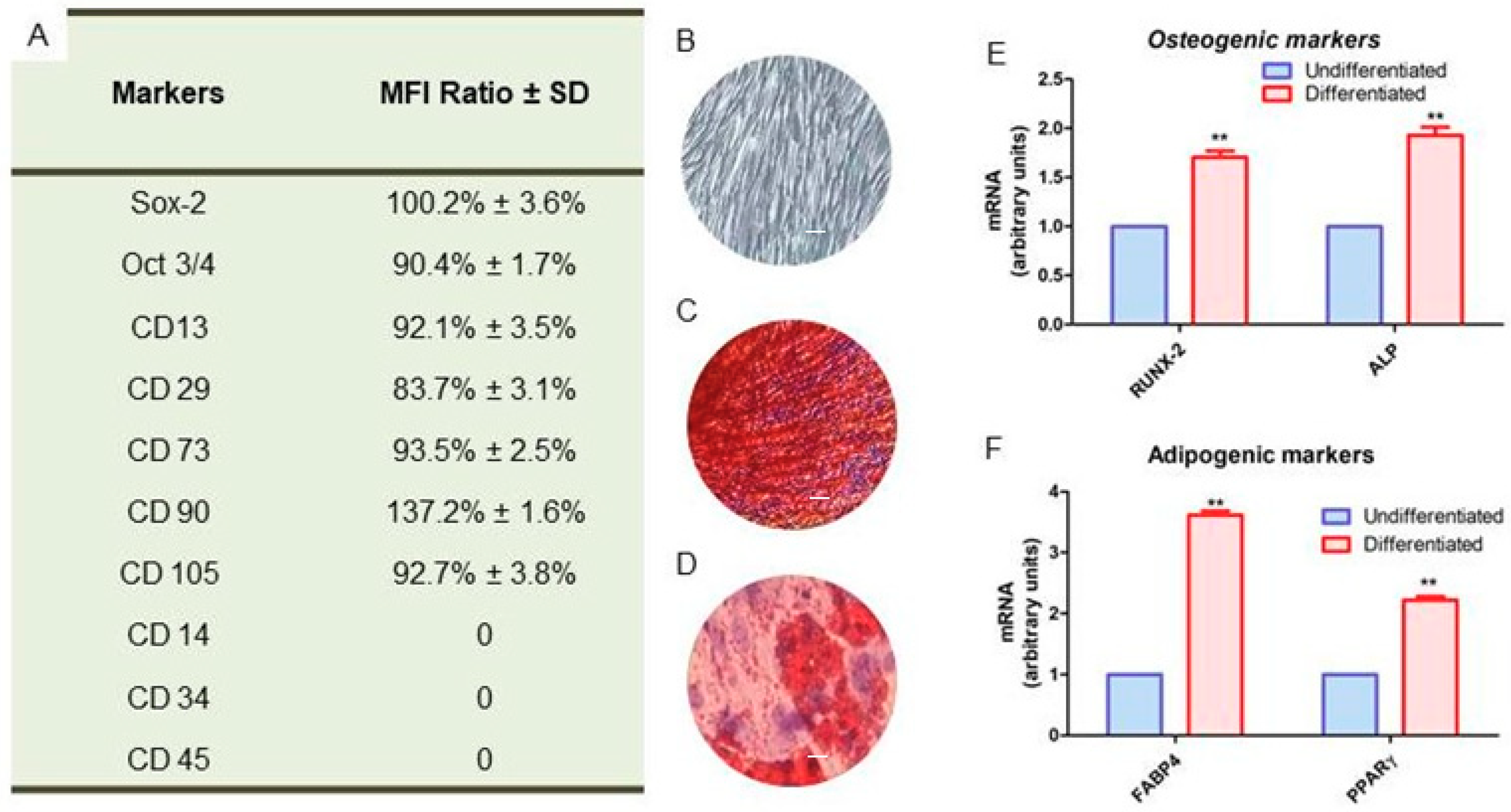
2.1. Cell Characterization
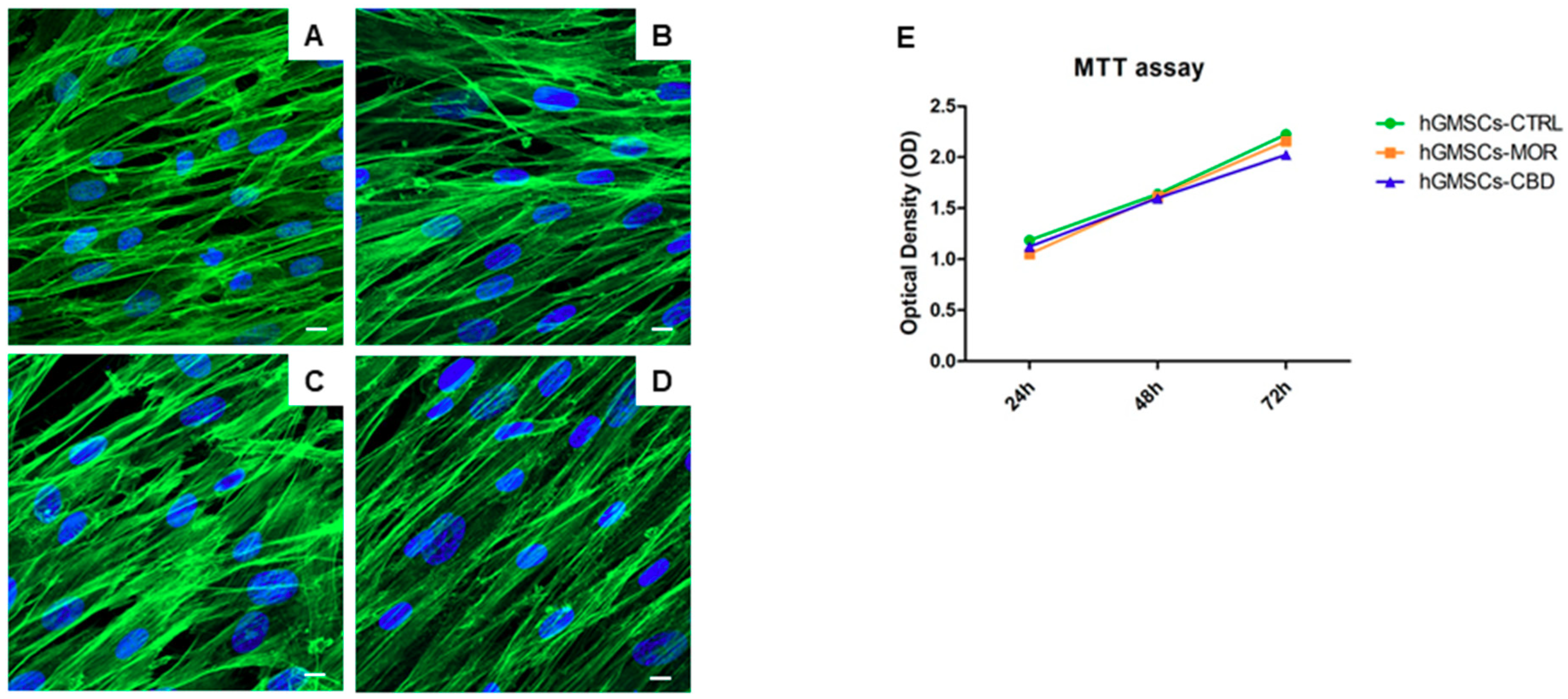
2.2. Effects on Morphology and Viability of MOR and CBD Treatments
2.3. Transcriptomic Analysis
2.4. Protein Expression
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Compounds Isolation and Purification
4.3. Cell Isolation
4.4. Cell Characterization
4.5. Cell Culture and Treatments
4.6. Cell Viability Assay
4.7. Morphological Analyses
4.8. RNA Extraction and NGS Analysis
4.9. Bioinformatics Analysis and Database Inspection
4.10. Western Blot Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s Disease |
| PD | Parkinson’s Disease |
| MS | Multiple Sclerosis |
| ALS | Amyotrophic Lateral Sclerosis |
| hGMSCs | human Gingival Mesenchymal Stem Cells |
| MOR | Moringin |
| CBD | Cannabidiol |
| IL-10 | Interleukin 10 |
| IL-1 | Interleukin 1 |
| IL-6 | Interleukin-6 |
| TNF-α | Tumor Necrosis Factor alpha |
| IL6R | IL-6 Receptor |
| TGF-β | Transform Growth Factor beta |
| hGMSCs-MOR | hGMSCs treated with MOR |
| hGMSCs-CBD | hGMSCs treated with CBD |
| RT-PCR | reverse transcription polymerase chain reaction |
| CLSM | Confocal Laser Scanning Microscopy |
| hGMSCs-CTRL | hGMSCs untreated |
| TNFR1 | TNF receptor 1 |
| RIPK1 | Receptor Interacting Protein Kinase 1 |
| IKK | IκB kinase |
| IL1R1 | IL-1 receptor type I |
| TIR | Toll-IL-1-Receptor |
| MAPK | Mitogen-Activated Protein Kinase |
| MAP3K3 | Mitogen-Activated Protein Kinase Kinase Kinase 3 |
| JAKs | Janus kinases |
| TGFβR1 | TGFβ Receptor type 1 |
| TGFβR2 | TGFβ Receptor type 2 |
| PP1 | Protein Phosphatase 1 |
| PBS | Phosphate Buffered Saline |
| MTT | 3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazoliumbromide |
References
- Hung, C.-W.; Chen, Y.-C.; Hsieh, W.-L.; Chiou, S.-H.; Kao, C.-L. Ageing and neurodegenerative diseases. Ageing Res. Rev. 2010, 9, S36–S46. [Google Scholar] [CrossRef] [PubMed]
- Sakthiswary, R.; Raymond, A.A. Stem cell therapy in neurodegenerative diseases: From principles to practice. Neural Regen. Res. 2012, 7, 1822. [Google Scholar] [PubMed]
- Ritfeld, G.J.; Roos, R.A.; Oudega, M. Stem cells for central nervous system repair and rehabilitation. PMR 2011, 3, S117–S122. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Gugliandolo, A.; Scionti, D.; Merciaro, I.; Cavalcanti, M.; Mazzon, E.; Trubiani, O. Biotherapeutic effect of gingival stem cells conditioned medium in bone tissue restoration. Int. J. Mol. Sci. 2018, 19, 329. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, C.; Akiyama, K.; Chai, Y.; Le, A.; Wang, Z.; Shi, S. Gingivae contain neural-crest-and mesoderm-derived mesenchymal stem cells. J. Dent. Res. 2013, 92, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Nauta, A.J.; Fibbe, W.E. Immunomodulatory properties of mesenchymal stromal cells. Blood 2007, 110, 3499–3506. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Kim, M.K.; Shin, M.S.; Lee, H.J.; Ko, J.H.; Wee, W.R.; Lee, J.H. The anti-inflammatory and anti-angiogenic role of mesenchymal stem cells in corneal wound healing following chemical injury. Stem Cells 2008, 26, 1047–1055. [Google Scholar] [CrossRef]
- Jaja-Chimedza, A.; Graf, B.L.; Simmler, C.; Kim, Y.; Kuhn, P.; Pauli, G.F.; Raskin, I. Biochemical characterization and anti-inflammatory properties of an isothiocyanate-enriched moringa (moringa oleifera) seed extract. PLoS ONE 2017, 12, e0182658. [Google Scholar] [CrossRef]
- Jaafaru, M.S.; Nordin, N.; Shaari, K.; Rosli, R.; Razis, A.F.A. Isothiocyanate from moringa oleifera seeds mitigates hydrogen peroxide-induced cytotoxicity and preserved morphological features of human neuronal cells. PLoS ONE 2018, 13, e0196403. [Google Scholar] [CrossRef]
- Rajan, T.S.; De Nicola, G.R.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. Anticancer activity of glucomoringin isothiocyanate in human malignant astrocytoma cells. Fitoterapia 2016, 110, 1–7. [Google Scholar] [CrossRef]
- Giacoppo, S.; Galuppo, M.; Montaut, S.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. An overview on neuroprotective effects of isothiocyanates for the treatment of neurodegenerative diseases. Fitoterapia 2015, 106, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Hannan, M.A.; Kang, J.-Y.; Mohibbullah, M.; Hong, Y.-K.; Lee, H.; Choi, J.-S.; Choi, I.S.; Moon, I.S. Moringa oleifera with promising neuronal survival and neurite outgrowth promoting potentials. J. Ethnopharmacol. 2014, 152, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Romeo, L.; Diomede, F.; Gugliandolo, A.; Scionti, D.; Giudice, F.L.; Cariccio, V.L.; Iori, R.; Bramanti, P.; Trubiani, O.; Mazzon, E. Moringin induces neural differentiation in the stem cell of the human periodontal ligament. Sci. Rep. 2018, 8, 9153. [Google Scholar] [CrossRef] [PubMed]
- Chiricosta, L.; Gugliandolo, A.; Diomede, F.; Pizzicannella, J.; Trubiani, O.; Iori, R.; Tardiolo, G.; Guarnieri, S.; Bramanti, P.; Mazzon, E. Moringin pretreatment inhibits the expression of genes involved in mitophagy in the stem cell of the human periodontal ligament. Molecules 2019, 24, 3217. [Google Scholar] [CrossRef]
- Chen, J.; Hou, C.; Chen, X.; Wang, D.; Yang, P.; He, X.; Zhou, J.; Li, H. Protective effect of cannabidiol on hydrogen peroxide-induced apoptosis, inflammation and oxidative stress in nucleus pulposus cells. Mol. Med. Rep. 2016, 14, 2321–2327. [Google Scholar] [CrossRef]
- Oláh, A.; Tóth, B.I.; Borbíró, I.; Sugawara, K.; Szöllõsi, A.G.; Czifra, G.; Pál, B.; Ambrus, L.; Kloepper, J.; Camera, E. Cannabidiol exerts sebostatic and antiinflammatory effects on human sebocytes. J. Clin. Investig. 2014, 124, 3713–3724. [Google Scholar] [CrossRef]
- Libro, R.; Scionti, D.; Diomede, F.; Marchisio, M.; Grassi, G.; Pollastro, F.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. Cannabidiol modulates the immunophenotype and inhibits the activation of the inflammasome in human gingival mesenchymal stem cells. Front. Physiol. 2016, 7, 559. [Google Scholar] [CrossRef]
- Libro, R.; Diomede, F.; Scionti, D.; Piattelli, A.; Grassi, G.; Pollastro, F.; Bramanti, P.; Mazzon, E.; Trubiani, O. Cannabidiol modulates the expression of alzheimer’s disease-related genes in mesenchymal stem cells. Int. J. Mol. Sci. 2016, 18, 26. [Google Scholar] [CrossRef]
- Rajan, T.S.; Scionti, D.; Diomede, F.; Grassi, G.; Pollastro, F.; Piattelli, A.; Cocco, L.; Bramanti, P.; Mazzon, E.; Trubiani, O. Gingival stromal cells as an in vitro model: Cannabidiol modulates genes linked with amyotrophic lateral sclerosis. J. Cell. Biochem. 2017, 118, 819–828. [Google Scholar] [CrossRef]
- Soundara Rajan, T.; Giacoppo, S.; Scionti, D.; Diomede, F.; Grassi, G.; Pollastro, F.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. Cannabidiol activates neuronal precursor genes in human gingival mesenchymal stromal cells. J. Cell. Biochem. 2017, 118, 1531–1546. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Rajan, T.S.; Giacoppo, S.; Diomede, F.; Ballerini, P.; Paolantonio, M.; Marchisio, M.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. The secretome of periodontal ligament stem cells from ms patients protects against eae. Sci. Rep. 2016, 6, 38743. [Google Scholar] [CrossRef] [PubMed]
- Augello, A.; Tasso, R.; Negrini, S.M.; Amateis, A.; Indiveri, F.; Cancedda, R.; Pennesi, G. Bone marrow mesenchymal progenitor cells inhibit lymphocyte proliferation by activation of the programmed death 1 pathway. Eur. J. Immunol. 2005, 35, 1482–1490. [Google Scholar] [CrossRef] [PubMed]
- Salami, F.; Tavassoli, A.; Mehrzad, J.; Parham, A. Immunomodulatory effects of mesenchymal stem cells on leukocytes with emphasis on neutrophils. Immunobiology 2018, 223, 786–791. [Google Scholar] [CrossRef]
- Rajan, T.S.; Giacoppo, S.; Iori, R.; De Nicola, G.R.; Grassi, G.; Pollastro, F.; Bramanti, P.; Mazzon, E. Anti-inflammatory and antioxidant effects of a combination of cannabidiol and moringin in lps-stimulated macrophages. Fitoterapia 2016, 112, 104–115. [Google Scholar] [CrossRef]
- Giacoppo, S.; Rajan, T.S.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. The α-cyclodextrin complex of the moringa isothiocyanate suppresses lipopolysaccharide-induced inflammation in raw 264.7 macrophage cells through akt and p38 inhibition. Inflamm. Res. 2017, 66, 487–503. [Google Scholar] [CrossRef]
- McCoy, M.K.; Tansey, M.G. Tnf signaling inhibition in the cns: Implications for normal brain function and neurodegenerative disease. J. Neuroinflamm. 2008, 5, 45. [Google Scholar] [CrossRef]
- Wajant, H.; Pfizenmaier, K.; Scheurich, P. Tumor necrosis factor signaling. Cell Death Differ. 2003, 10, 45. [Google Scholar] [CrossRef]
- Peltzer, N.; Walczak, H. Cell death and inflammation-a vital but dangerous liaison. Trends Immunol. 2019, 40, 387–402. [Google Scholar] [CrossRef]
- Weinlich, R.; Green, D.R. The two faces of receptor interacting protein kinase-1. Mol. Cell 2014, 56, 469–480. [Google Scholar] [CrossRef]
- Tokunaga, F.; Nakagawa, T.; Nakahara, M.; Saeki, Y.; Taniguchi, M.; Sakata, S.-i.; Tanaka, K.; Nakano, H.; Iwai, K. Sharpin is a component of the nf-κb-activating linear ubiquitin chain assembly complex. Nature 2011, 471, 633. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Gaynor, R.B. Ikappab kinases: Key regulators of the nf-kappab pathway. Trends Biochem. Sci. 2004, 29, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Israël, A. The ikk complex, a central regulator of nf-κb activation. Cold Spring Harb. Perspect. Biol. 2010, 2, a000158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Descargues, P.; Sil, A.K.; Karin, M. Ikkα, a critical regulator of epidermal differentiation and a suppressor of skin cancer. EMBO J. 2008, 27, 2639–2647. [Google Scholar] [CrossRef] [Green Version]
- Dondelinger, Y.; Jouan-Lanhouet, S.; Divert, T.; Theatre, E.; Bertin, J.; Gough, P.J.; Giansanti, P.; Heck, A.J.; Dejardin, E.; Vandenabeele, P.; et al. Nf-kappab-independent role of ikkalpha/ikkbeta in preventing ripk1 kinase-dependent apoptotic and necroptotic cell death during tnf signaling. Mol. Cell 2015, 60, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Yao, F.; Long, L.-Y.; Deng, Y.-Z.; Feng, Y.-Y.; Ying, G.-Y.; Bao, W.-D.; Li, G.; Guan, D.-X.; Zhu, Y.-Q.; Li, J.-J. Rack1 modulates nf-κb activation by interfering with the interaction between traf2 and the ikk complex. Cell Res. 2014, 24, 359. [Google Scholar] [CrossRef] [Green Version]
- Fujikura, D.; Ito, M.; Chiba, S.; Harada, T.; Perez, F.; Reed, J.; Uede, T.; Miyazaki, T. Clipr-59 regulates tnf-α-induced apoptosis by controlling ubiquitination of rip1. Cell Death Dis. 2012, 3, e264. [Google Scholar] [CrossRef]
- Xu, G.; Tan, X.; Wang, H.; Sun, W.; Shi, Y.; Burlingame, S.; Gu, X.; Cao, G.; Zhang, T.; Qin, J. Ubiquitin-specific peptidase 21 inhibits tumor necrosis factor α-induced nuclear factor κb activation via binding to and deubiquitinating receptor-interacting protein 1. J. Biol. Chem. 2010, 285, 969–978. [Google Scholar] [CrossRef] [Green Version]
- Walczak, H. Tnf and ubiquitin at the crossroads of gene activation, cell death, inflammation, and cancer. Immunol. Rev. 2011, 244, 9–28. [Google Scholar] [CrossRef]
- Mihaly, S.R.; Ninomiya-Tsuji, J.; Morioka, S. Tak1 control of cell death. Cell Death Differ. 2014, 21, 1667–1676. [Google Scholar] [CrossRef] [Green Version]
- Micheau, O.; Tschopp, J. Induction of tnf receptor i-mediated apoptosis via two sequential signaling complexes. Cell 2003, 114, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Tummers, B.; Green, D.R. Caspase-8: Regulating life and death. Immunol. Rev. 2017, 277, 76–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedmann, E.; Hauben, E.; Maylandt, K.; Schleeger, S.; Vreugde, S.; Lichtenthaler, S.F.; Kuhn, P.-H.; Stauffer, D.; Rovelli, G.; Martoglio, B. Sppl2a and sppl2b promote intramembrane proteolysis of tnfα in activated dendritic cells to trigger il-12 production. Nat. Cell Biol. 2006, 8, 843. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, N.C.; Shayakhmetov, D.M. Interleukin 1α and the inflammatory process. Nat. Immunol. 2016, 17, 906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suren, C.; Lukashova, L.; Wischmann, J.; Wulsten, D.; Wildemann, B.; Von, R.E.-R.; Holzmann, B.; Mayer-Kuckuk, P. Abnormal bone architecture in mice expressing myd88 in cells of the osteoclast lineage. J. Biol. Regul. Homeost. Agents 2019, 33, 1105–1111. [Google Scholar] [PubMed]
- Lappas, M. The adaptor protein p62 mediates nuclear factor κb activation in response to inflammation and facilitates the formation of prolabor mediators in human myometrium. Reprod. Sci. 2017, 24, 762–772. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Yang, J.; Lin, Y.; Walker, C.; Cheng, J.; Liu, Z.-g.; Su, B. Differential regulation of interleukin 1 receptor and toll-like receptor signaling by mekk3. Nat. Immunol. 2004, 5, 98. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. Il-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Hibi, M.; Murakami, M.; Saito, M.; Hirano, T.; Taga, T.; Kishimoto, T. Molecular cloning and expression of an il-6 signal transducer, gp130. Cell 1990, 63, 1149–1157. [Google Scholar] [CrossRef]
- Waetzig, G.H.; Chalaris, A.; Rosenstiel, P.; Suthaus, J.; Holland, C.; Karl, N.; Uriarte, L.V.; Till, A.; Scheller, J.; Grötzinger, J. N-linked glycosylation is essential for the stability but not the signaling function of the interleukin-6 signal transducer glycoprotein 130. J. Biol. Chem. 2010, 285, 1781–1789. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.S.; Xu, P.-Z.; Gottlob, K.; Chen, M.-L.; Sokol, K.; Shiyanova, T.; Roninson, I.; Weng, W.; Suzuki, R.; Tobe, K. Growth retardation and increased apoptosis in mice with homozygous disruption of the akt1 gene. Genes Dev. 2001, 15, 2203–2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryskalin, L.; Lazzeri, G.; Flaibani, M.; Biagioni, F.; Gambardella, S.; Frati, A.; Fornai, F. Mtor-dependent cell proliferation in the brain. BioMed Res. Int. 2017, 2017, 7082696. [Google Scholar] [CrossRef] [PubMed]
- Roussa, E.; Farkas, L.M.; Krieglstein, K. Tgf-β promotes survival on mesencephalic dopaminergic neurons in cooperation with shh and fgf-8. Neurobiol. Dis. 2004, 16, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Annes, J.P.; Munger, J.S.; Rifkin, D.B. Making sense of latent tgfβ activation. J. Cell Sci. 2003, 116, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heldin, C.-H.; Moustakas, A. Signaling receptors for tgf-β family members. Cold Spring Harb. Perspect. Biol. 2016, 8, a022053. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Donahoe, P.K.; Zervos, A.S. Specific interaction of type i receptors of the tgf-beta family with the immunophilin fkbp-12. Science 1994, 265, 674–676. [Google Scholar] [CrossRef]
- Heldin, C.-H.; Miyazono, K.; Ten Dijke, P. Tgf-β signalling from cell membrane to nucleus through smad proteins. Nature 1997, 390, 465. [Google Scholar] [CrossRef]
- Varelas, X.; Sakuma, R.; Samavarchi-Tehrani, P.; Peerani, R.; Rao, B.M.; Dembowy, J.; Yaffe, M.B.; Zandstra, P.W.; Wrana, J.L. Taz controls smad nucleocytoplasmic shuttling and regulates human embryonic stem-cell self-renewal. Nat. Cell Biol. 2008, 10, 837. [Google Scholar] [CrossRef]
- Stroschein, S.L.; Wang, W.; Zhou, S.; Zhou, Q.; Luo, K. Negative feedback regulation of tgf-beta signaling by the snon oncoprotein. Science 1999, 286, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Liang, M.; Feng, X.-H. Smurf2 is a ubiquitin e3 ligase mediating proteasome-dependent degradation of smad2 in transforming growth factor-β signaling. J. Biol. Chem. 2000, 275, 36818–36822. [Google Scholar] [CrossRef] [Green Version]
- Datta, P.K.; Chytil, A.; Gorska, A.E.; Moses, H.L. Identification of strap, a novel wd domain protein in transforming growth factor-β signaling. J. Biol. Chem. 1998, 273, 34671–34674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Feng, X.-H. Regulation of tgf-β signalling by protein phosphatases. Biochem. J. 2010, 430, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunelli, D.; Tavecchio, M.; Falcioni, C.; Frapolli, R.; Erba, E.; Iori, R.; Rollin, P.; Barillari, J.; Manzotti, C.; Morazzoni, P. The isothiocyanate produced from glucomoringin inhibits nf-kb and reduces myeloma growth in nude mice in vivo. Biochem. Pharmacol. 2010, 79, 1141–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, C.; van Loon, J.; Ruschioni, S.; De Nicola, G.R.; Olsen, C.E.; Iori, R.; Agerbirk, N. Taste detection of the non-volatile isothiocyanate moringin results in deterrence to glucosinolate-adapted insect larvae. Phytochemistry 2015, 118, 139–148. [Google Scholar] [CrossRef]
- Taglialatela-Scafati, O.; Pagani, A.; Scala, F.; De Petrocellis, L.; Di Marzo, V.; Grassi, G.; Appendino, G. Cannabimovone, a cannabinoid with a rearranged terpenoid skeleton from hemp. Eur. J. Org. Chem. 2010, 2010, 2067–2072. [Google Scholar] [CrossRef]
- Mammana, S.; Gugliandolo, A.; Cavalli, E.; Diomede, F.; Iori, R.; Zappacosta, R.; Bramanti, P.; Conti, P.; Fontana, A.; Pizzicannella, J. Human gingival mesenchymal stem cells (gmscs) pre-treated with vesicular moringin nanostructures as a new therapeutic approach in a mouse model of spinal cord injury. J. Tissue Eng. Regen. Med. 2019, 13, 1109–1121. [Google Scholar]
- Diomede, F.; Zini, N.; Pizzicannella, J.; Merciaro, I.; Pizzicannella, G.; D’Orazio, M.; Piattelli, A.; Trubiani, O. 5-aza exposure improves reprogramming process through embryoid body formation in human gingival stem cells. Front. Genet. 2018, 9, 419. [Google Scholar] [CrossRef] [Green Version]
- Pizzicannella, J.; Diomede, F.; Merciaro, I.; Caputi, S.; Tartaro, A.; Guarnieri, S.; Trubiani, O. Endothelial committed oral stem cells as modelling in the relationship between periodontal and cardiovascular disease. J. Cell. Physiol. 2018, 233, 6734–6747. [Google Scholar] [CrossRef]
- Trubiani, O.; Ballerini, P.; Murmura, G.; Pizzicannella, J.; Giuliani, P.; Buccella, S.; Caputi, S. Toll-like receptor 4 expression, interleukin-6,-8 and ccl-20 release, and nf-kb translocation in human periodontal ligament mesenchymal stem cells stimulated with lps-p. Gingivalis. Eur. J. Inflamm. 2012, 10, 81–89. [Google Scholar] [CrossRef]
- Pizzicannella, J.; Gugliandolo, A.; Orsini, T.; Fontana, A.; Ventrella, A.; Mazzon, E.; Bramanti, P.; Diomede, F.; Trubiani, O. Engineered extracellular vesicles from human periodontal-ligament stem cells increase vegf/vegfr2 expression during bone regeneration. Front. Physiol. 2019, 10, 512. [Google Scholar] [CrossRef] [Green Version]
- Diomede, F.; D’Aurora, M.; Gugliandolo, A.; Merciaro, I.; Ettorre, V.; Bramanti, A.; Piattelli, A.; Gatta, V.; Mazzon, E.; Fontana, A.; et al. A novel role in skeletal segment regeneration of extracellular vesicles released from periodontal-ligament stem cells. Int. J. Nanomed. 2018, 13, 3805–3825. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Name | Fold Change hGMSCs-MOR | Fold Change hGMSCs-CBD | Q-Value hGMSCs-MOR | Q-Value hGMSCs-CBD |
|---|---|---|---|---|---|
| AKT1 | AKT serine/threonine kinase 1 | 0.23 | 0.34 | 2.50 × 10−3 | 9.79 × 10−5 |
| CASP8 | Caspase 8 | −0.65 | −1.41 | 2.11 × 10−4 | 9.79 × 10−5 |
| CHUK | Component of inhibitor of nuclear factor kappa B kinase complex | 0.35 | −1.19 | 9.77 × 10−3 | 9.79 × 10−5 |
| CLIP3 | CAP-Gly domain containing linker protein 3 | −0.71 | −1.03 | 2.11 × 10−4 | 9.79 × 10−5 |
| CYLD | CYLD lysine 63 deubiquitinase | −0.32 | 0.47 | 2.11 × 10−4 | 9.79 × 10−5 |
| GNB2L1 | Receptor for activated C kinase 1 | 0.13 | −0.51 | 1.43 × 10−3 | 9.79 × 10−5 |
| IL1R1 | Interleukin 1 receptor type 1 | 0.43 | −0.46 | 2.11 × 10−4 | 1.91 × 10−3 |
| IL6ST | Interleukin 6 signal transducer | −0.43 | −0.12 | 2.11 × 10−4 | 2.18 × 10−2 |
| MAP3K3 | Mitogen-activated protein kinase kinase kinase 3 | 0.74 | 0.87 | 2.11 × 10−4 | 9.79 × 10−5 |
| MAP3K7 | Mitogen-activated protein kinase kinase kinase 7 | −0.47 | −0.38 | 2.11 × 10−4 | 1.27 × 10−3 |
| MTOR | Mechanistic target of rapamycin kinase | 0.34 | 0.0 | 2.11 × 10−4 | >5 × 10−2 |
| MYD88 | MYD88 innate immune signal transduction adaptor | −1.05 | −2.26 | 2.11 × 10−4 | 9.79 × 10−5 |
| PIK3CA | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha | −0.37 | 0.32 | 1.74 × 10−3 | 1.35 × 10−3 |
| PIK3CB | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit beta | −0.52 | 0.75 | 1.35 × 10−2 | 9.79 × 10−5 |
| PIK3CD | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit delta | −0.79 | −1.59 | 2.11 × 10−4 | 9.79 × 10−5 |
| RIPK1 | Receptor interacting serine/threonine kinase 1 | −0.37 | 0.50 | 2.44 × 10−2 | 2.29 × 10−3 |
| RPS27A | Ribosomal protein S27a | 0.16 | −0.16 | 2.11 × 10−4 | 9.79 × 10−5 |
| SHARPIN | SHANK associated RH domain interactor | 0.50 | −2.45 | 2.70 × 10−2 | 1.90 × 10−4 |
| SPPL2A | Signal peptide peptidase like 2A | −0.22 | 0.32 | 2.56 × 10−2 | 3.64 × 10−4 |
| SQSTM1 | Sequestosome 1 | −0.94 | −0.44 | 2.11 × 10−4 | 9.79 × 10−5 |
| STAT3 | Signal transducer and activator of transcription 3 | −0.40 | −1.15 | 2.11 × 10−4 | 9.79 × 10−5 |
| TNFRSF1A | TNF receptor superfamily member 1A | −0.35 | 0.38 | 2.11 × 10−4 | 9.79 × 10−5 |
| TYK2 | Tyrosine kinase 2 | 0.41 | −1.33 | 7.66 × 10−4 | 9.79 × 10−5 |
| UBA52 | Ubiquitin A-52 residue ribosomal protein fusion product 1 | 0.14 | −0.92 | 2.11 × 10−4 | 9.79 × 10−5 |
| UBB | Ubiquitin B | −0.52 | 0.70 | 2.11 × 10−4 | 9.79 × 10−5 |
| UBC | Ubiquitin C | 0.30 | −0.43 | 2.11 × 10−4 | 9.79 × 10−5 |
| USP21 | Ubiquitin specific peptidase 21 | −0.51 | 0.98 | 3.46 × 10−2 | 2.77 × 10−4 |
| Gene | Name | Fold Change hGMSCs-MOR | Fold Change hGMSCs-CBD | Q-Value hGMSCs-MOR | Q-Value hGMSCs-CBD |
|---|---|---|---|---|---|
| FKBP1A | FKBP prolyl isomerase 1A | 0.77 | −0.78 | 2.11 × 10−4 | 9.79 × 10−5 |
| FURIN | Furin, paired basic amino acid cleaving enzyme | 0.37 | 0.39 | 1.27 × 10−3 | 9.79 × 10−5 |
| NCOR1 | Nuclear receptor corepressor 1 | −0.17 | 0.13 | 8.18 × 10−3 | 1.71 × 10−2 |
| PPP1CA | Protein phosphatase 1 catalytic subunit alpha | 0.20 | −0.55 | 2.41 × 10−2 | 9.79 × 10−5 |
| PPP1CB | Protein phosphatase 1 catalytic subunit beta | −0.44 | 0.78 | 2.11 × 10−4 | 9.79 × 10−5 |
| PPP1R15A | Protein phosphatase 1 regulatory subunit 15A | −0.24 | 1.01 | 3.32 × 10−2 | 9.79 × 10−5 |
| SMAD3 | SMAD family member 3 | −0.75 | −0.19 | 2.11 × 10−4 | 1.91 × 10−3 |
| SMAD4 | SMAD family member 4 | 0.18 | −0.27 | 3.32 × 10−2 | 2.77 × 10−4 |
| SMURF2 | SMAD specific E3 ubiquitin protein ligase 2 | 0.23 | 0.19 | 1.40 × 10−2 | 9.79 × 10−5 |
| STRAP | Serine/threonine kinase receptor associated protein | 0.20 | 0.70 | 4.56 × 10−2 | 9.79 × 10−5 |
| TGFBR1 | Transforming growth factor beta receptor 1 | 0.44 | 0.56 | 7.66 × 10−4 | 9.79 × 10−5 |
| TGFBR2 | Transforming growth factor beta receptor 2 | 0.86 | −0.52 | 2.11 × 10−4 | 9.79 × 10−5 |
| WWTR1 | WWTR1 | −0.34 | −0.57 | 2.94 × 10−3 | 9.79 × 10−5 |
| XPO1 | Exportin 1 | 0.46 | 0.21 | 2.11 × 10−4 | 1.59 × 10−3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiricosta, L.; Silvestro, S.; Pizzicannella, J.; Diomede, F.; Bramanti, P.; Trubiani, O.; Mazzon, E. Transcriptomic Analysis of Stem Cells Treated with Moringin or Cannabidiol: Analogies and Differences in Inflammation Pathways. Int. J. Mol. Sci. 2019, 20, 6039. https://doi.org/10.3390/ijms20236039
Chiricosta L, Silvestro S, Pizzicannella J, Diomede F, Bramanti P, Trubiani O, Mazzon E. Transcriptomic Analysis of Stem Cells Treated with Moringin or Cannabidiol: Analogies and Differences in Inflammation Pathways. International Journal of Molecular Sciences. 2019; 20(23):6039. https://doi.org/10.3390/ijms20236039
Chicago/Turabian StyleChiricosta, Luigi, Serena Silvestro, Jacopo Pizzicannella, Francesca Diomede, Placido Bramanti, Oriana Trubiani, and Emanuela Mazzon. 2019. "Transcriptomic Analysis of Stem Cells Treated with Moringin or Cannabidiol: Analogies and Differences in Inflammation Pathways" International Journal of Molecular Sciences 20, no. 23: 6039. https://doi.org/10.3390/ijms20236039
APA StyleChiricosta, L., Silvestro, S., Pizzicannella, J., Diomede, F., Bramanti, P., Trubiani, O., & Mazzon, E. (2019). Transcriptomic Analysis of Stem Cells Treated with Moringin or Cannabidiol: Analogies and Differences in Inflammation Pathways. International Journal of Molecular Sciences, 20(23), 6039. https://doi.org/10.3390/ijms20236039