3. Discussion
Our analysis of root transcriptomes of two barley genotypes, which differed in their level of drought tolerance, allowed us to select several genes encoding transcription factors that may be involved in better drought survival. Among the ten genes that were specifically activated or repressed in roots of drought-tolerant CamB, one encodes the TSO1-like CXC protein, which is the homolog of the human TESMIN gene that is expressed mainly in testes and ovaries [
21]. In Arabidopsis, the TSO1 gene functions in floral tissue development, inflorescence meristem organization, and control of cell division [
22]. Importantly, it is required for the correct organization of shoot and root apical meristems [
23]. It was shown that TSO1 acts as a repressor of the
MYB3R1 gene, which in turn maintains the repressed state of G2/M-specific genes in the roots, preventing cell proliferation. Thus, high expression of
TSO1 allows the maintenance of root proliferation in the root meristematic zone [
23]. In our study, the up-regulation of the
TSO1-like gene in CamB roots suggests that it may be involved in the promotion of better drought tolerance of this genotype.
In our data set, no differential expression of the
MYB3R1 homologue was noticed, but we found two other genes from the MYB superfamily, belonging to the GARP-G2-like group that were specific to CamB roots (HORVU2Hr1G020140 and HORVU7Hr1G096430). The first one was up-regulated and was annotated as two-component response regulator
ARR18, but the homology search in Ensembl Plants database showed that its orthologues in Arabidopsis (
HRS1,
HHO2, or
HHO3) and rice (
NIGT1) are involved in nitrate and phosphate signaling in roots [
24] and in the presence of nitrate they repress the nitrate-starvation response [
25]. The second one was down-regulated in CamB roots and is the orthologue of Arabidopsis
PHL6 and rice
PHR4 genes, that are activated upon limiting phosphate availability and regulate phosphate-starvation response pathway [
26]. It was shown that high nitrate reduces lateral root elongation [
27] and a similar observation was true for drought treated plants [
28]. On the other hand, phosphorus starvation increases lateral root formation and the density of root hairs [
27], placing these genes in a pathway that may influence the root plasticity in response to the soil environment. Expression pattern of barley orthologues of NIGH1 and PHR4 observed in our study suggests that they may also play a role in drought response although their precise function in this type of stress remains to be elucidated.
An interesting gene (HORVU5Hr1G062040) that was up-regulated in CamB roots shows similarity to human
SCAI, the suppressor of cancer cell invasion gene that inhibits the invasive migration of tumor cells [
29]. SCAI protein interacts with KDM3B, a histone demethylase protein that removes methyl groups from H3K9me1/2, which is the mark of repressed chromatin state [
30], and with the SWI/SNF chromatin remodeling complex [
31]. Thus, the HORVU5Hr1G062040 gene plays a role in the regulation of chromatin status and its transcriptional availability. Two other DEGs involved in epigenetic regulation were found in CamB roots: a down-regulated HORVU5Hr1G061120 gene encoding a bromo-adjacent homology (BAH) domain-containing protein and an up-regulated HORVU6Hr1G034680 gene for a methyl-CpG-binding domain-containing protein. The proteins with a BAH domain recognize methylated state of histone H3 lysines that are the hallmarks of the heterochromatin [
32]. The methyl-CpG-binding proteins interact with methylated CpG sites in DNA and are also responsible for silencing the target chromatin regions [
33]. The direction of expression changes of these genes in the roots of the CamB genotype suggests that the efficient response to drought may require the formation of an active chromatin status of genes that are targeted by SCAI and BAH proteins. The repression of transcription of other genes, which are under the regulation mediated by the methyl-CpG-binding protein, may be also needed.
In the roots of Maresi, contrary to CamB, more genotype-specific DEGs for gene expression regulatory proteins were found, and the great majority of them were down-regulated by the stress. This finding suggests that in a drought-susceptible cultivar, more dynamic changes occur after the drought treatment. It is not clear whether the prevalence of down-regulatory mechanisms is an indicator of a lower acclimation ability to the stress or it just suggests that Maresi enters a substantially different pathway of drought response than CamB. Many detected TFs belong to large families with members of positive and negative expression regulatory functions, and there are reports of their both up- or down-regulation upon drought. Such observations were made, for example, for genes from AP2/ERF [
34,
35], bHLH, MADS, MYB [
8], NAC [
9], or WRKY families [
7]. Thus, a more detailed analysis targeted to single genes is needed to detect their specific impact on drought response in the Maresi cultivar. Nevertheless, some possible molecular response pathways have emerged that may be characteristic of the roots of this drought-susceptible genotype.
One pathway may be connected to jasmonic acid (JA) signaling and the action of three genes: HORVU7Hr1G041230, HORVU3Hr1G050590, and HORVU4Hr1G003040 encoding a TIFY3A TF, WRKY and an NPR1 protein, respectively. The TIFY3A TF belongs to the JAZ subfamily of TIFY, which is known to repress the action of Arabidopsis MYC2 TF and its homologues—the positive regulators of JA-dependent responses. The important effect of such response is an inhibition of primary root growth [
36]. The involvement of TIFY TFs in drought tolerance was demonstrated by the overexpression study of rice
OsTIFY11a, which resulted in the enhancement of dehydration stress resistance [
37]. Down-regulation of this gene in Maresi roots suggests that this pathway of possible drought tolerance may be negatively affected in this cultivar.
Similar, negative regulation of JA response was found in Arabidopsis for another factor,
WRKY50 [
38], a homologue of which HORVU3Hr1G050590 was again down-regulated in roots of Maresi. Another down-regulated gene (HORVU4Hr1G003040), encoding a regulatory protein NPR1, may also be placed within the JA-related response. It is a repressor of JA signaling that acts through the induction of the salicylic acid pathway [
39], and its overexpression in Arabidopsis leads to oxidative stress resistance [
40]. Taking all of this together, down-regulation of the above-mentioned genes may lead to the lack or lower rate of JA signaling repression in Maresi roots, which in turn may diminish the efficient drought response.
Nonetheless, more complexity to the image of this response is added by the down-regulation of two
KNOTTED1-like (
KN1) transcription factor genes (HORVU2Hr1G061320 and HORVU5Hr1G098570). The KN1 TF negatively modulates the accumulation of gibberellin (GA) through the control of ga2ox1, which is an enzyme that leads to the catalysis of the GA [
41]. Thus, down-regulation of
KN1 genes in Maresi roots may slow-down the degradation of GA, which in turn may promote root cell proliferation and elongation [
42]. Moreover, it was shown that there is a crosstalk between GA and JA, mediated by MYC and DELLA proteins that compete with each other to bind with JAZ TF. In the presence of GA, DELLA protein is directed to degradation, and then JAZ and MYC may interact with each other, which prevents MYC from activation of JA-dependent genes and leads to the repression of JA-driven response [
43]. This complex response of Maresi indicates the necessity of a substantial rebuilding of root metabolism under the mild drought in a susceptible cultivar, which does not take place in a tolerant genotype.
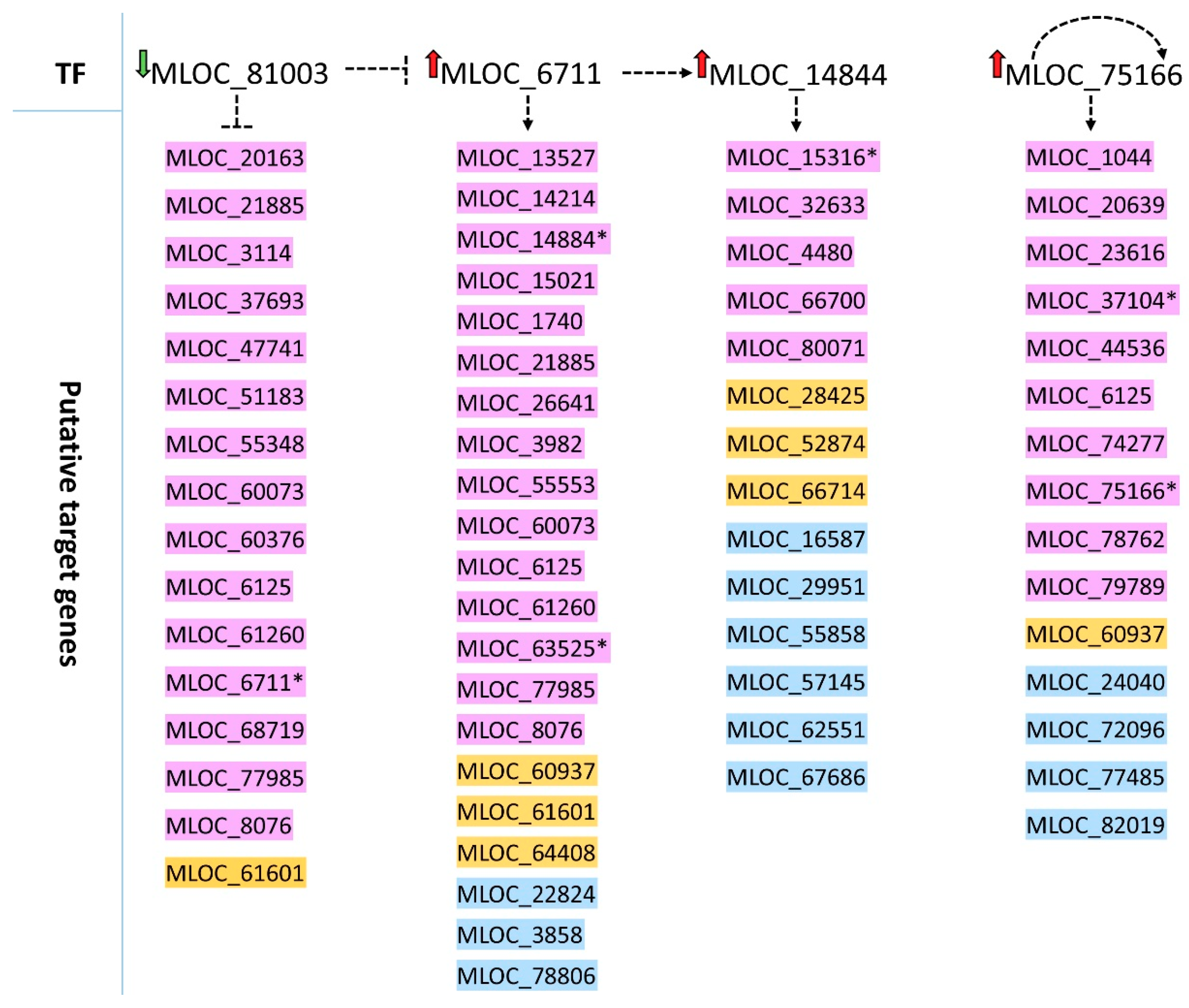
Based on the enrichment analysis, we performed a prediction of possible relationships between differentially expressed TF genes and all DEGs that were identified in the roots of both genotypes. We found four TFs that had overrepresented targets among root DEGs, which may place them near the top of the regulatory network of the mild drought response in roots. Two of them encode TFs from the bZIP family: MLOC_81003 (HORVU5Hr1G014170) with the highest similarity to rice bZIP88 and MLOC_6711 (HORVU1Hr1G090030), similar to Arabidopsis G-box binding factor 2 (GBF2). Many bZIP TFs are involved in stress signaling and act in an ABA-dependent manner [
44]. Our data suggest a possible negative regulation of
GBF2 by bZIP88 (HORVU5Hr1G014170) in drought response in barley roots, as the up-regulated
GBF2 gene was found among targets of down-regulated bZIP88. A down-regulation of
bZIP88 was also observed in rice subjected to various abiotic stresses, including drought [
45]. Interestingly, a DNA binding activity of some G-group bZIPs was found to be regulated by light and reactive oxygen species (ROS) [
46], which are commonly produced in tissues upon the abiotic stresses. Thus, one may hypothesize that ROS produced in drought treated roots may trigger the regulation of gene expression downstream of
GBF2. Another TF gene with overrepresented targets encodes heat stress factor (HORVU7Hr1G056820) orthologous to rice, HSFB-2b gene. HSFs from class B are considered to be transcriptional repressors [
47], contrary to our enrichments analysis data, which pointed to several up-regulated DEGs downstream to the
HFSB2b gene (also up-regulated). On the other hand, a study of seed germination under heat stress in Arabidopsis showed the induction of the
HSFB2b gene, which was accompanied by the accumulation of 49 heat shock proteins [
48] (HSP). In our analysis, two up-regulated genes (HORVU3Hr1G007500 and HORVU3Hr1G020520) encoding HSPs were found as targets of HSFB2b. Moreover, a study of maize transcriptome suggests that the
HSFB2b gene may be important in drought tolerance in this species, as
HSFB2b expression was specifically activated in a drought-tolerant genotype [
49]. Nevertheless, at this stage of our analysis, we treat the above enrichment data only as putative interactions that may serve to drive hypothesis on novel regulatory connections in the gene network, and which need to be verified based on gene-specific studies.
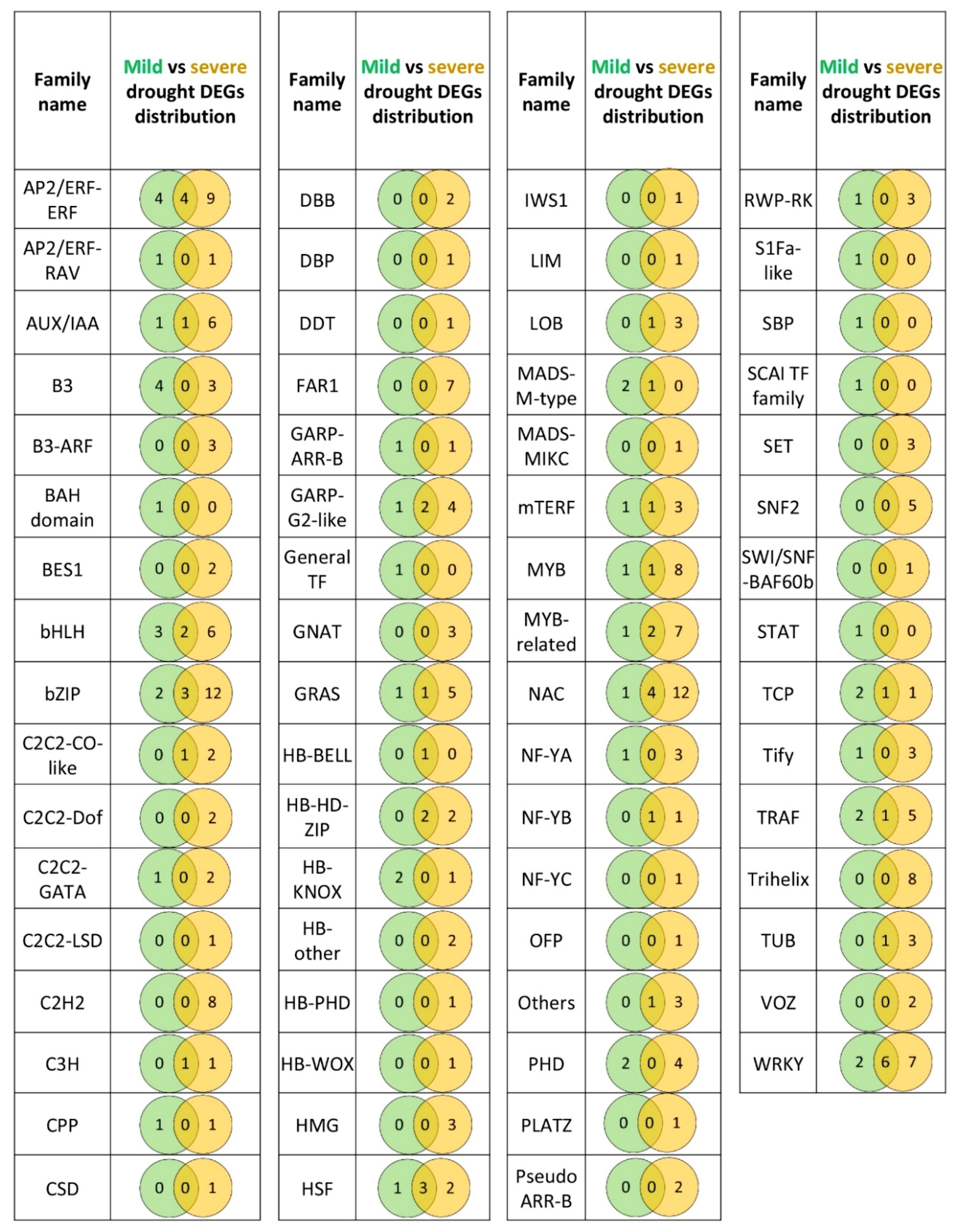
The comparison of mild and severe drought stress regimes showed a significant increase in the number of DEGs involved in gene expression regulation when a severe drought was applied. Among all the regulatory DEGs detected after mild drought treatment, about 50% were also differentially expressed after severe stress. Additionally, the number of DEGs that were specific to only one stress regime was four times higher after severe stress, compared to the mild drought. Such results suggest that a substantial part of root drought response that is initiated upon moderate stress may also persist in more severe conditions. It is important to emphasize, however, that the details of this response differed in a genotype-dependent manner. A general scheme that emerged from our analysis suggests that the drought-sensitive genotype goes through more profound changes in the transcriptome than the drought-tolerant form, especially after the severe stress. The differences between the two drought regimes that were observed for DEGs with expression regulatory function were also reflected in the expression of all remaining genes involved in the other biological processes. Gene ontology analysis showed that DEGs identified after mild drought belong to similar biological and functional categories as genes differentially expressed after severe stress [
20]. However, when we take into consideration the number of DEGs, then more DEGs are found within each GO category when the stress is stronger, and again, more DEGs are present in the drought-sensitive genotype [
20]. The same was true for the TF DEGs detected in the presented study—a much higher number of TF DEGs were found after severe stress in each TF family, and this increase in number was more prominent in the Maresi genotype. A remarkably higher number of DEGs characteristic for severe drought were noticed, for example, within the AP/ERF family of ethylene responsive factors and AUX/IAA family involved in auxin signaling. A similar increase in gene number was noticed for the bZIP family, including factors involved in ABA signaling. It was shown that both hormones enter a crosstalk important for a drought response and trigger the inhibition of lateral root formation [
50], making their regulatory network essential for shaping the root system architecture under stress. Severe stress also resulted in a higher number of genes from GRAS family, which may be placed within gibberellin signaling [
51] and the genes from the Tify family involved in jasmonate signaling [
52]. Interestingly, the Tify TFs detected under severe stress included down-regulated genes in CamB, suggesting that the drought-tolerant genotype may enter a similar response pathway related to jasmonates as the sensitive cultivar, but only after much more adverse environmental conditions.
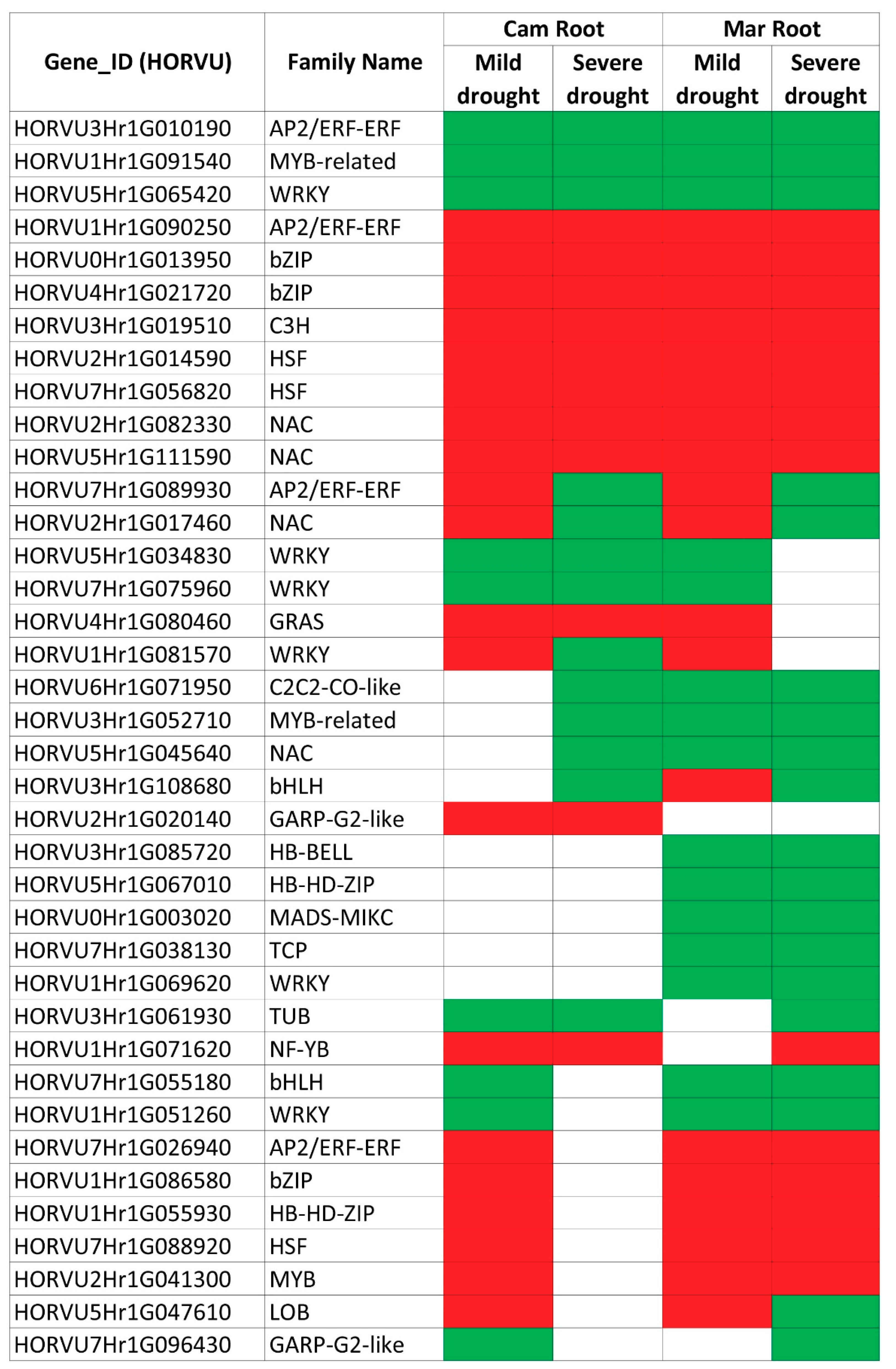
An interesting finding was the detection of five genes, which exhibited opposite expression changes after mild and severe stresses, and in the case of two of them, some functional characteristics may be related to the root function under drought. The first one (HORVU7Hr1G089930), which encodes a TF from the AP/ERF family is an orthologue of the Arabidopsis gene for the SHINE factor that is involved in wax synthesis. Overexpression of this gene caused the increase of wax biosynthesis and altered its composition, resulting in a higher cuticle permeability in leaves and better drought tolerance [
53]. The Arabiopsis
SHINE gene is also expressed in roots and is probably involved in suberin deposition [
54]. Suberisation is considered to have a negative impact on water and solute transport, but it may also prevent an uncontrolled backflow of water to the surrounding soil under unfavorable environmental conditions [
55]. During the day and in optimal soil moisture, the main force driving water transport is the transpiration, and water is transported via the apoplast. During the night or when stomata are closed due to drought stress, water flow from cell-to-cell and is driven by the root pressure. In the latter scenario, the suberin acts as a barrier limiting apoplastic transport [
56]. The barley
SHINE orthologue was up-regulated under mild drought in our study, but down-regulated after the severe water shortage conditions in both genotypes. It is possible that SHINE allows the increase of suberization when the signal of water shortage is perceived by the roots, and consequently, it helps to actively maintain the transport of limited, but still available water to the shoot, even if the transpiration rate is reduced. On the other hand, under the severe drought, when water is very limited, a high deposition of suberin may be unfavorable as it may increase hydraulic resistance of the root to the extent it prevents water acquisition.
The second gene with the opposite expression change under moderate and severe drought (HORVU5Hr1G047610) belongs to the LOB (LATERAL ORGAN BOUNDARIES) family and was also up-regulated after mild drought in both genotypes and down-regulated in severe drought, but only in the Maresi cultivar. LOB transcription factors are involved in the development of lateral organs. The first LOB gene identified in Arabidopsis is expressed at the boundaries between stem cells and developing leaf primordia at shoot apical meristem [
57]. Several
LOB genes are important for lateral root formation driven by auxin signaling and modulated by ABA under stress conditions [
58,
59]. A maize
RTCS gene from this family is responsible for the initiation and maintenance of embryonic seminal roots and shoot-born root primordia [
60], and the
ARL1 gene in rice is involved in adventitious roots formation [
61]. At present, it is difficult to predict the precise function of the
LOB gene identified in the presented study in barley, but its differential expression in roots only and not in leaves suggests its importance for shaping root architecture and building an appropriate root system under the moderate but not in the severe stress conditions.
Another aspect that emerged from our transcriptomic study was the discovery of genes that function in the photosynthesis process, but were differentially expressed under drought in roots of one or both barley genotypes. They included genes encoding proteins of oxygen-evolving complex, together with proteins that build photosystem I (PSI) and PSII in leaf chloroplasts, and most of them were up-regulated by drought stress. This finding is in agreement with our previous study, where several genes annotated as photosynthesis-related were also differentially expressed in roots upon severe drought [
20]. We hypothesize that such result may suggest a specific role of root plastids as ROS scavenging centers, which may help root cells to cope with oxidative stress conditions generated by drought. Differential expression of genes involved in photosynthesis in roots under drought is not uncommon and was also detected in other studies, for example, in cotton [
62], chickpea [
63] poplar [
64], and pine [
65].
Our analysis also suggests that under the mild drought stress, different genes involved in photosynthesis are differentially expressed in leaves and the roots. We found, for example, up-regulation of a gene for RuBisCO activase A in Maresi leaves and differential expression of two genes encoding chlorophyll a-b binding proteins in leaves of both genotypes, whereas another gene for chlorophyll a-b binding protein was up-regulated in roots only. Similarly, we noticed down-regulation of a gene encoding one of PSII components in leaves, whereas, in roots, two other genes for PSII proteins were up-regulated. This may suggest that drought results in an organ-specific expression of genes with similar function, although we are aware that we analyzed only one time-point of the stress treatment, so we only spotted a subset of the dynamic changes of the transcriptomes and we cannot rule out that some of these genes exhibit different expression patterns at the early stage of the stress. Such possibility is partially supported by the research of Chmielewska et al. [
18] who used plant material from the same experiment (the same genotypes and drought treatment) for the analysis of leaves and roots proteomes and metabolomes. On the protein level, they found a higher accumulation of oxygen-evolving enhancer protein in leaves of Maresi and a ferredoxin-NADP+ oxidoreductase in leaves of CamB, while in our study the differential expression of their transcripts was not found. It is likely that the up-regulation of transcription could occur at an earlier stage of the stress application and on the 10th day, the differences were visible only on the protein level. Similarly, Chmielewska et al. [
18] found a higher accumulation of RuBisCO activase B in both genotypes and RuBisCO activase A protein in the leaves of Maresi only, while in our analysis, only the latter protein is in agreement with its transcript up-regulation. Nevertheless, taking together the above aspects of our analysis, the photosynthesis-related genes may be considered as a source of better drought response of roots, due to their possible anti-oxidative role protecting macromolecules from oxidative damage in this organ.
To conclude, we have found a substantial number of transcription factor encoding genes that are involved in the drought response in roots of barley. The use of microarrays, despite their limitations, allowed us to make an unbiased comparison of transcriptome changes induced by mild drought to the response of the same genotypes to severe stress, which we studied previously [
20]. The comparability of both analyses was ensured by the use of the same type of arrays and experimental methodology, except for the drought regime, in both experiments. We are aware, however, that additional use of the RNA-Seq method may give a more in-depth view on global gene expression changes. Nevertheless, our analysis suggests that genotype-dependent drought tolerance is accompanied by a lower number of gene expression changes than response of the susceptible genotype. The gene expression regulation is probably directed toward the maintenance of root meristems proliferation and a yet unidentified network of epigenetic changes that may allow a better acclimation of the plant to the stress conditions and may be triggered by factors involved in chromatin remodeling and DNA methylation. We found that a part of the drought response mechanisms is controlled by transcription regulators from Tiffy, WRKY, HB-KNOX, and mTERF families that may act within jasmonate and gibberellin signaling pathways. Our data also show that the activation of several TFs may be important in mild drought but not in severe drought response. We believe that the TF-encoding genes identified in the present study may be used as future targets for the enhancement of drought tolerance in barley and related species.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}