Development of Fluorescently Labeled SSEA-3, SSEA-4, and Globo-H Glycosphingolipids for Elucidating Molecular Interactions in the Cell Membrane



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Synthesis of Fluorescent Probes
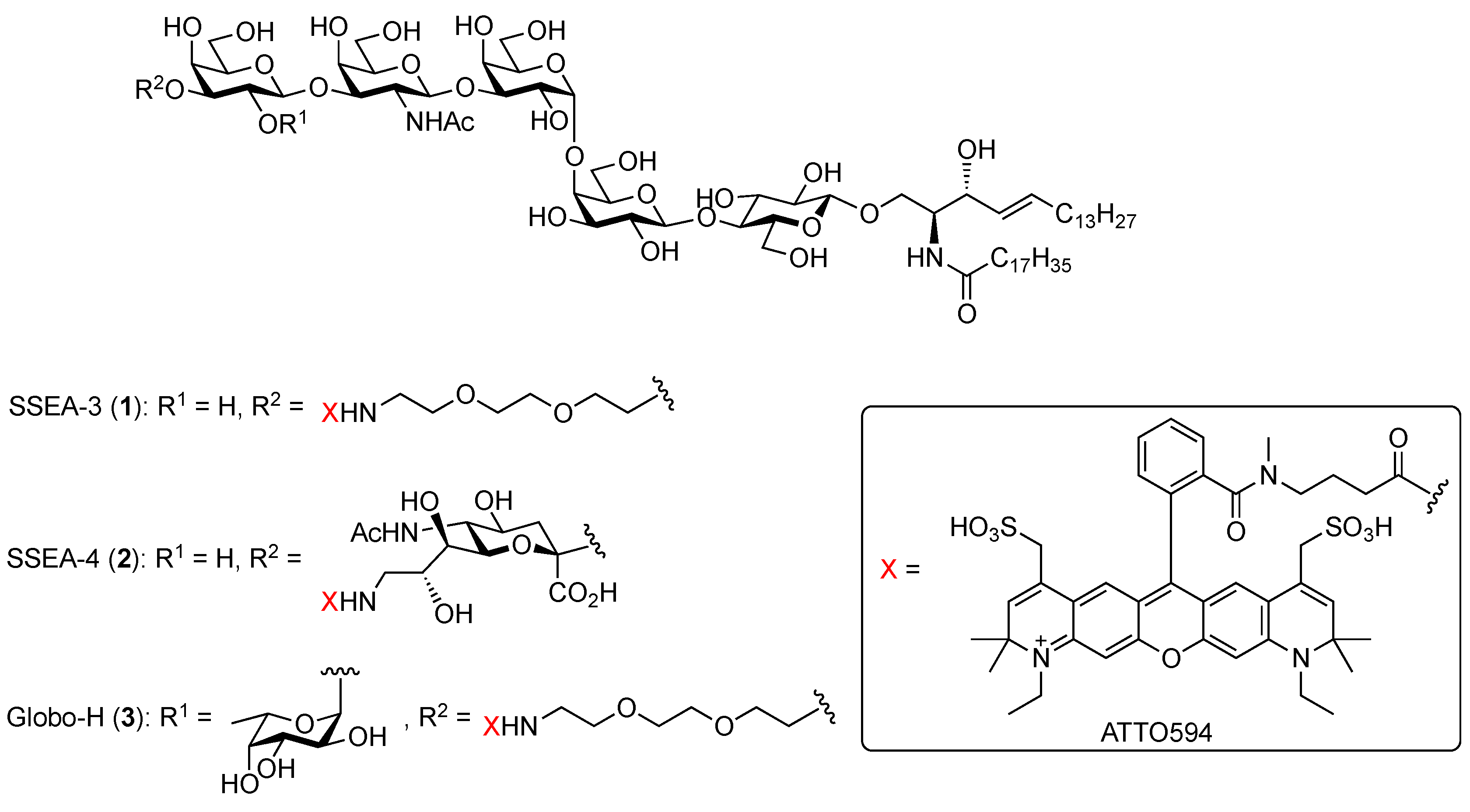
2.1.1. Molecular Design and Synthesis Plan
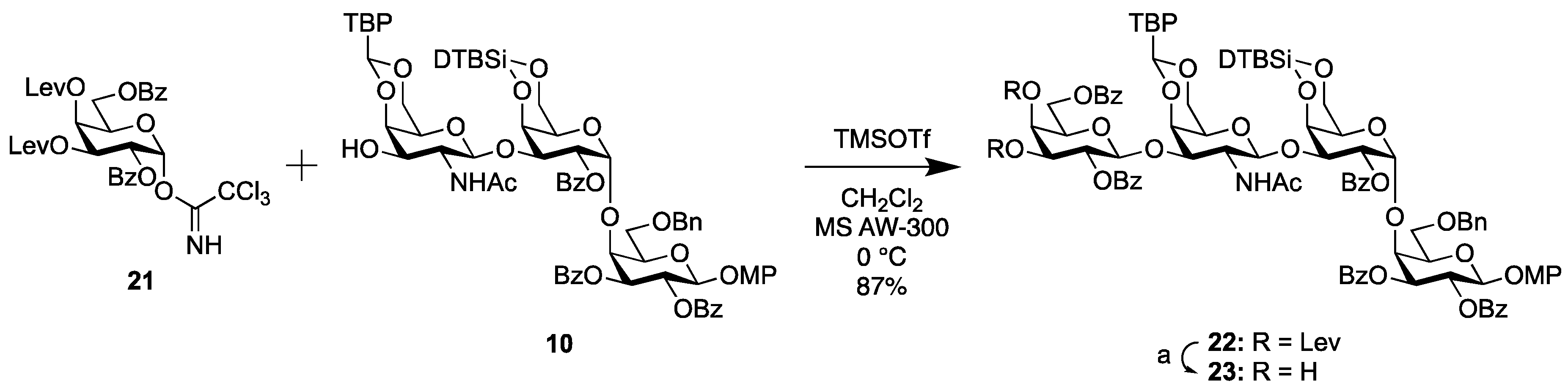
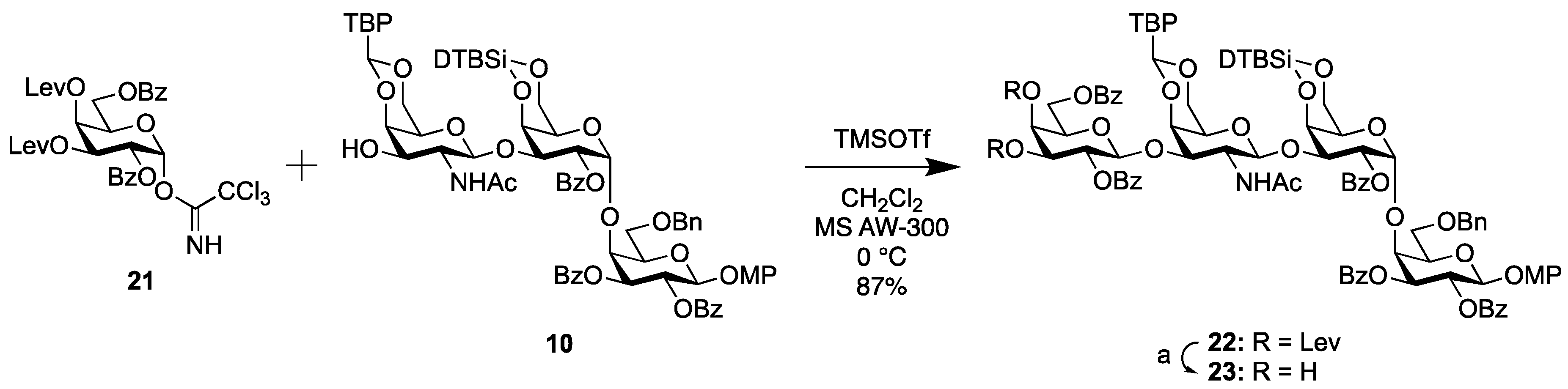
2.1.2. Synthesis of the Trisaccharide Common Acceptor
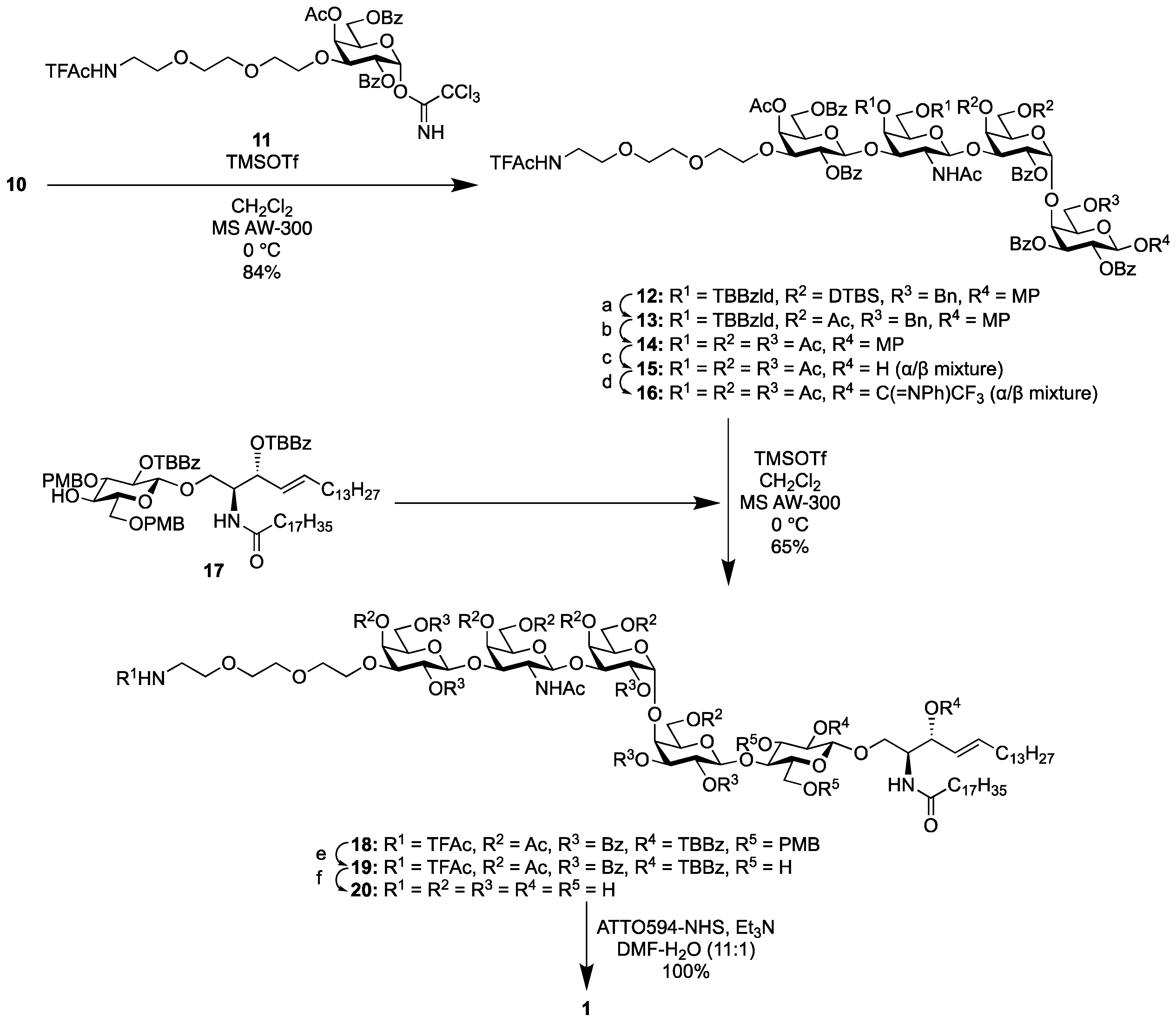
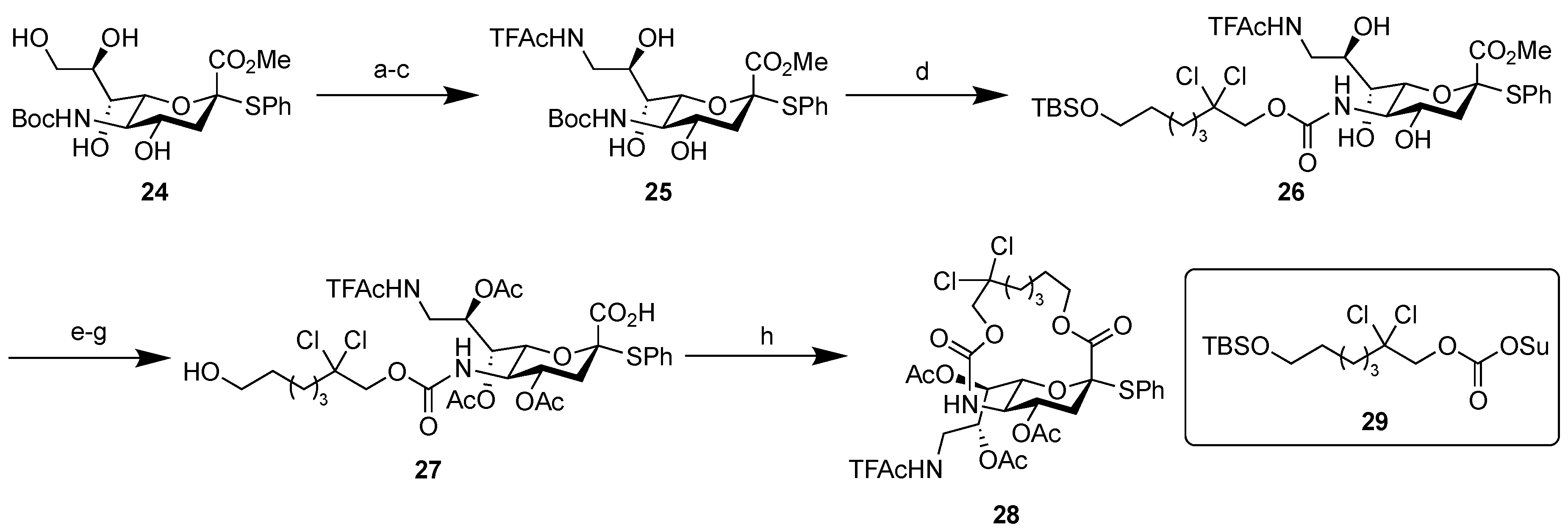
2.1.3. Synthesis of the Fluorescent SSEA-3 Probe
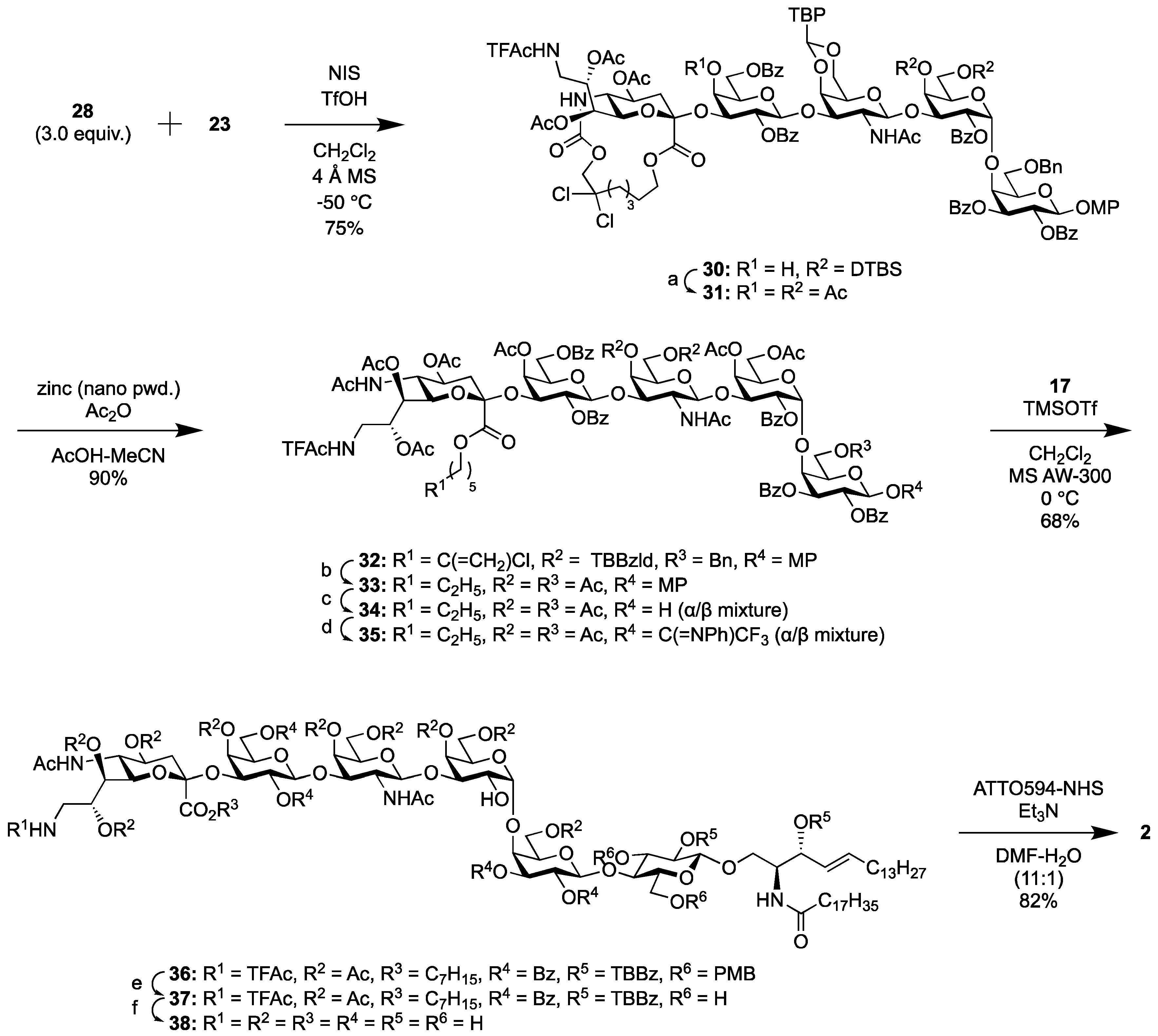
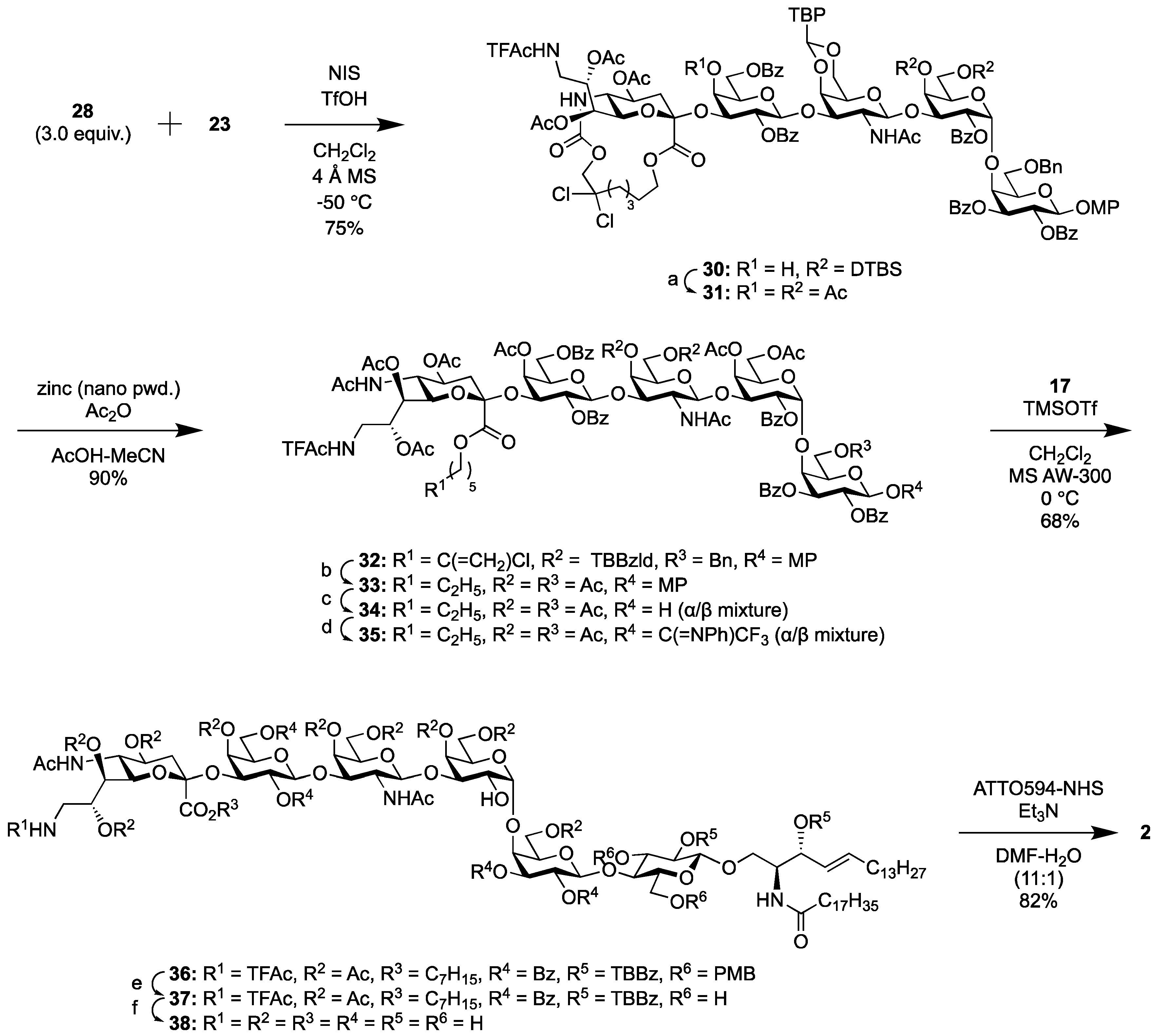
2.1.4. Synthesis of the Fluorescent SSEA-4 Probe
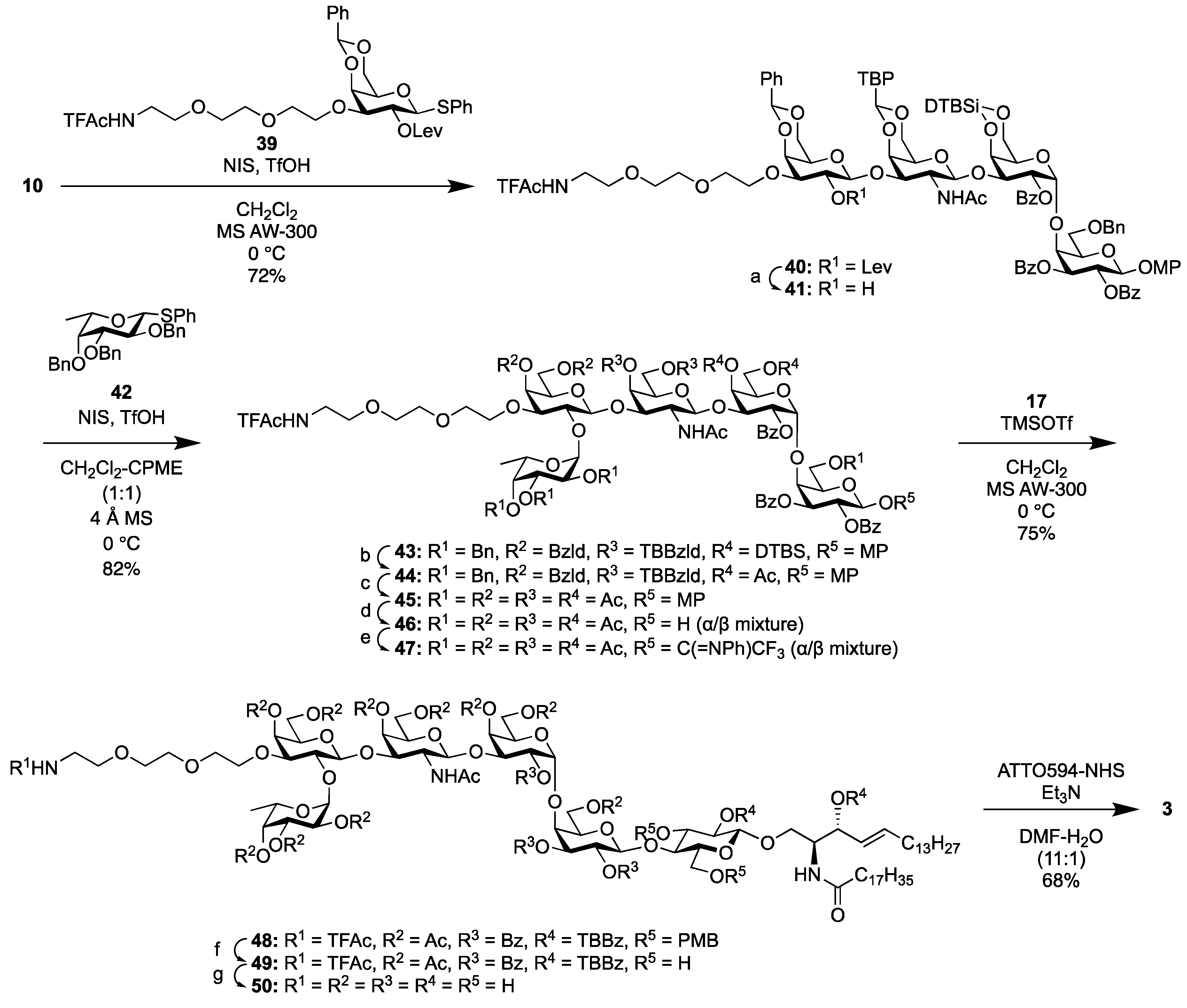
2.1.5. Synthesis of the Fluorescent Globo-H Probe
2.2. Biophysical and Biochemical Evaluation
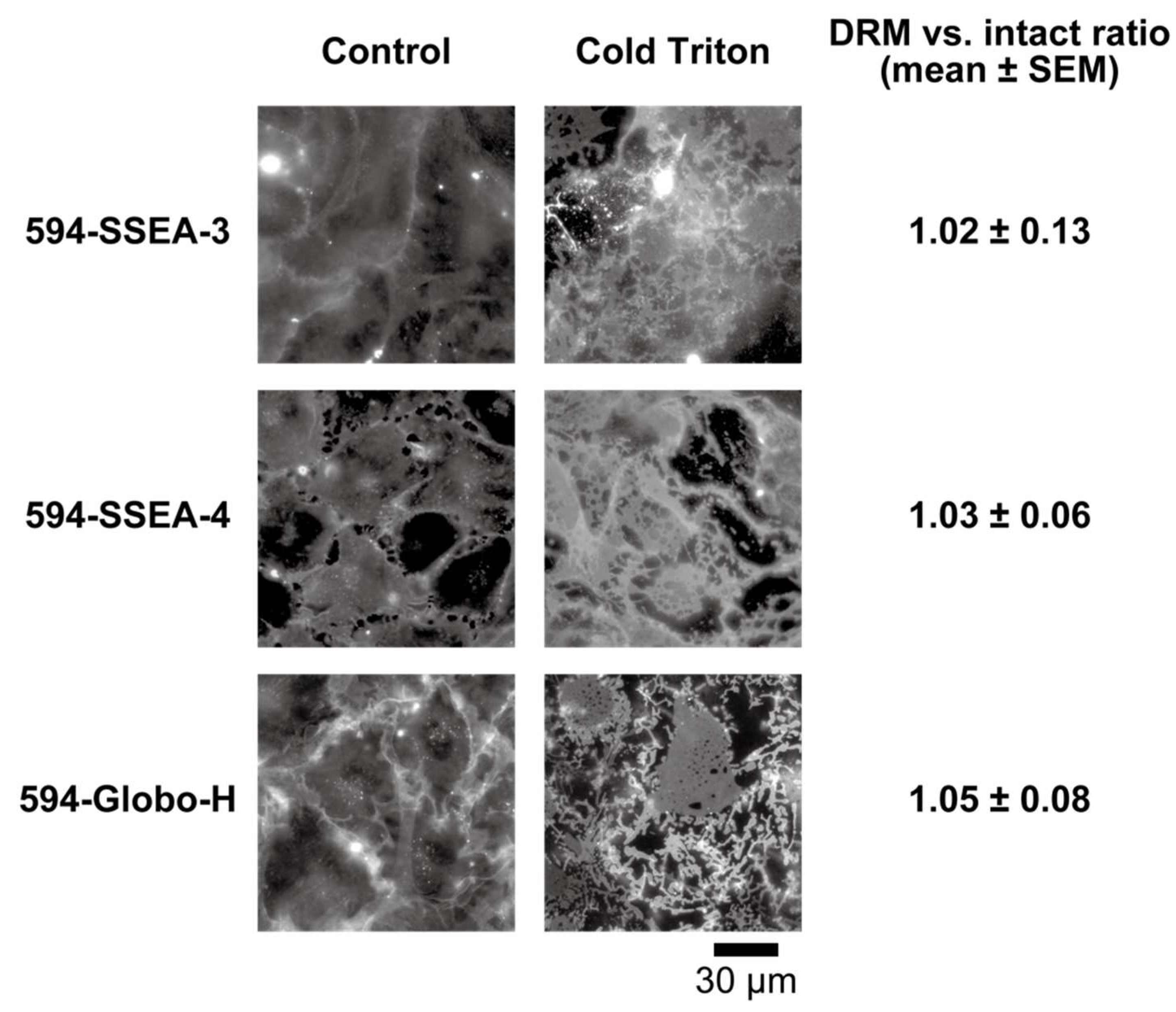
2.2.1. Cold-Triton Solubility of the Fluorescent Probes in the PM
2.2.2. Partitioning of the Fluorescent Probes between the Lo/Ld Phases
3. Materials and Methods
3.1. Chemical Section
3.2. Biological Evaluations
3.2.1. Evaluation of Cold-Triton Solubility
3.2.2. Formation and Microscopic Observation of GPMVs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Cer | Ceramide |
| Fuc | Fucose |
| Gal | Galactose |
| Glc | Glucose |
| Neu | Neuraminic acid |
| PC | Phosphatidylcholine |
References
- Schnaar, R.L.; Kinoshita, T. Glycosphingolipids. In Essentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, R., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Springer Harbor: New York, NY, USA, 2015–2017; Chapter 11.
- Lingwood, C.A. Glycosphingolipid functions. Cold Spring Harb. Perspect. Biol. 2011, 3, a004788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Gerl, M.J. Revitalizing membrane rafts: New tools and insights. Nat. Rev. Mol. Cell Biol. 2010, 11, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S. Cell adhesion/recognition and signal transduction through glycosphingolipid microdomains. Glycoconj. J. 2000, 17, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Komura, N.; Suzuki, K.G.N.; Ando, H.; Konishi, M.; Imamura, A.; Ishida, H.; Kusumi, A.; Kiso, M. Syntheses of fluorescent gangliosides for the studies of raft domains. Methods Enzymol. 2017, 597, 239–263. [Google Scholar] [PubMed]
- Koikeda, M.; Komura, N.; Tanaka, H.-N.; Imamura, A.; Ishida, H.; Kiso, M.; Ando, H. Synthesis of ganglioside analogs containing fluorescently labeled GalNAc for single-molecule imaging. J. Carbohydr. Chem. 2019, 38, 509–527. [Google Scholar] [CrossRef]
- Komura, N.; Suzuki, K.G.N.; Ando, H.; Konishi, M.; Koikeda, M.; Imamura, A.; Chadda, R.; Fujuwara, T.K.; Tsuboi, H.; Sheng, R.; et al. Raft-based interactions of gangliosides with a GPI-anchored receptor. Nat. Chem. Biol. 2016, 12, 402–410. [Google Scholar] [CrossRef]
- Suzuki, K.G.N.; Ando, H.; Komura, N.; Fujiwara, T.K.; Kiso, M.; Kusumi, A. Development of new ganglioside probes and unraveling of raft domain structure by single-molecule imaging. Biochim. Biophys. Acta 2017, 1861, 2494–2506. [Google Scholar] [CrossRef]
- Suzuki, K.G.N.; Ando, H.; Komura, N.; Fujiwara, T.; Kiso, M.; Kusumi, A. Unraveling of lipid raft organization in cell plasma membranes by single-molecule imaging of ganglioside probes. Adv. Exp. Med. Biol. 2018, 1104, 41–58. [Google Scholar]
- Hakomori, S.; Siddiqui, B.; Li, Y.-T.; Li, S.-C.; Hellerqvist, C.G. Anomeric structures of “globoside” and ceramide trihexoside of human erythrocytes and “BHK” fibroblasts. J. Biol. Chem. 1971, 246, 2271–2277. [Google Scholar]
- Hakomori, S. Structure and function of glycosphingolipids and sphingolipids: Recollections and future trends. Biochim. Biophys. Acta 2008, 1780, 325–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, J.K.; Draper, J.S.; Baillie, H.S.; Fishel, S.; Thomson, J.A.; Moore, H.; Andrews, P.W. Preimplantation human embryos and embryonic stem cells show comparable expression of stage-specific embryonic antigens. Stem Cells 2002, 20, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brimble, S.N.; Sherrer, E.S.; Uhl, E.W.; Wang, E.; Kelly, S.; Merrill, A.H., Jr.; Robins, A.J.; Schulz, T.C. The cell surface glycosphingolipids SSEA-3 and SSEA-4 are not essential for human ESC pluripotency. Stem Cells 2007, 25, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Slambrouck, S.V.; Steelant, F.A. Clustering of monosialyl-Gb5 initiates downstream signaling events leading to invasion of MCF-7 breast cancer cells. Biochem. J. 2007, 401, 689–699. [Google Scholar] [CrossRef] [Green Version]
- Saito, S.; Orikasa, S.; Satoh, M.; Ohyama, C.; Ito, A.; Takahashi, T. Expression of globo-series gangliosides in human renal cell carcinoma. Jpn. J. Cancer Res. 1997, 88, 652–659. [Google Scholar] [CrossRef]
- Charafe-Jauffret, E.; Ginestier, C.; Iovino, F.; Wicinski, J.; Cervera, N.; Finetti, P.; Hur, M.H.; Diebel, M.E.; Monville, F.; Dutcher, J.; et al. Breast cancer cell lines contain functional cancer stem cells with metastatic capacity and a distinct molecular signature. Cancer Res. 2009, 69, 1302–1313. [Google Scholar] [CrossRef] [Green Version]
- Hung, T.-C.; Lin, C.-W.; Hsu, T.-L.; Wu, C.-Y.; Wong, C.-W. Investigation of SSEA-4 binding protein in breast cancer cells. J. Am. Chem. Soc. 2013, 135, 5934–5937. [Google Scholar] [CrossRef]
- Bremer, E.G.; Levery, S.B.; Sonnino, S.; Ghidoni, R.; Canevari, S.; Kannagi, R.; Hakomori, S. Characterization of a glycosphingolipid antigen defined by the monoclonal antibody MBr1 expressed in normal and neoplastic epithelial cells of human mammary gland. J. Biol. Chem. 1984, 259, 14773–14777. [Google Scholar]
- Sato, B.; Katagiri, Y.U.; Miyado, K.; Akutsu, H.; Miyagawa, Y.; Horiuchi, Y.; Nakajima, H.; Okita, H.; Umezawa, A.; Hata, J.; et al. Preferential localization of SSEA-4 in interfaces between blastomeres of mouse preimplantation embryos. Biochem. Biophys. Res. Commun. 2007, 364, 838–843. [Google Scholar] [CrossRef] [Green Version]
- Steelant, W.F.; Kawakami, Y.; Ito, A.; Hanada, K.; Bruyneel, E.A.; Mareel, M.; Hakomori, S. Monosialyl-Gb5 organized with cSrc and FAK in GEM of human breast carcinoma MCF-7 cells defines their invasive properties. FEBS Lett. 2002, 531, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Nunomura, S.; Ogawa, T. A total synthesis of a stage specific embryonic antigen-3 (SSEA-3), globopentaosyl ceramide, IV3GalGb4Cer. Use of 2,4,6-trimethylbenzoyl group as a stereocontrolling auxiliary. Tetrahedron Lett. 1988, 29, 5681–5684. [Google Scholar] [CrossRef]
- Park, T.K.; Kim, I.J.; Danishefsky, S.J. A total synthesis of a stage specific pentasaccharide embryogenesis marker. Tetrahedron Lett. 1995, 36, 9089–9092. [Google Scholar] [CrossRef]
- Ishida, H.; Miyawaki, R.; Kiso, M.; Hasegawa, A. Synthetic studies on sialoglycoconjugates 82: First total synthesis of sialyl globopentaosyl ceramide (V3Neu5AcGb5Cer) and its positional isomer (V6Neu5AcGb5Cer). J. Carbohydr. Chem. 1996, 15, 163–182. [Google Scholar] [CrossRef]
- Lassaletta, J.M.; Schmidt, R.R. Versatile approach to the synthesis of globoside glycosphingolipids synthesis of sialyl-galactosyl-globoside. Tetrahedron Lett. 1995, 36, 4209–4212. [Google Scholar] [CrossRef]
- Bilodeau, M.T.; Park, T.K.; Hu, S.; Randolph, J.T.; Danishefsky, S.J.; Livingston, P.O.; Zhang, S. Total synthesis of a human breast tumor associated antigen. J. Am. Chem. Soc. 1995, 117, 7840–7841. [Google Scholar] [CrossRef]
- Imamura, A.; Ando, H.; Ishida, H.; Kiso, M. Ganglioside GQ1b: Efficient total synthesis and the expansion to synthetic derivatives to elucidate its biological roles. J. Org. Chem. 2009, 74, 3009–3023. [Google Scholar] [CrossRef]
- Imamura, A.; Ando, H.; Korogi, S.; Tanabe, G.; Muraoka, O.; Ishida, H.; Kiso, M. Di-tert-butylsilylene (DTBS) group-directed α-selective galactosylation unaffected by C-2 participating functionalities. Tetrahedron Lett. 2003, 44, 6725–6728. [Google Scholar] [CrossRef]
- Imamura, A.; Matsuzawa, N.; Sakai, S.; Udagawa, T.; Nakashima, S.; Ando, H.; Ishida, H.; Kiso, M. The origin of high stereoselectivity in di-tert-butylsilylene-directed α-galactosylation. J. Org. Chem. 2016, 81, 9086–9104. [Google Scholar] [CrossRef]
- Asano, S.; Tanaka, H.-N.; Imamura, A.; Ishida, H.; Ando, H. p-tert-Butyl groups improve the utility of aromatic protective groups in carbohydrate synthesis. Org. Lett. 2019, 21, 4197–4200. [Google Scholar] [CrossRef]
- Furusawa, K. Removal of cyclic di-t-butylsilanediyl protecting groups using tributylamine hydrofluoride (TBAHF) reagent. Chem. Lett. 1989, 18, 509–510. [Google Scholar] [CrossRef]
- Yu, B.; Tao, H. Glycosyl trifluoroacetimidates. Part 1: Preparation and application as new glycosyl donors. Tetrahedron Lett. 2001, 42, 2405–2407. [Google Scholar] [CrossRef]
- Komura, N.; Kato, K.; Udagawa, T.; Asano, S.; Tanaka, H.-N.; Imamura, A.; Ishida, H.; Kiso, M.; Ando, H. Constrained sialic acid donors enable selective synthesis of α-glycosides. Science 2019, 364, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Ando, H.; Ishihara, H.; Koketsu, M. Sialylation reactions with 5-N,7-O-carbonyl-protected sialyl donors: Unusual stereoselectivity with nitrile solvent assistance. Carbohydr. Res. 2008, 343, 1585–1593. [Google Scholar] [CrossRef]
- Sugimura, T.; Hagiya, K. Di-2-methoxyethyl azodicarboxylate (DMEAD): An inexpensive and separation-friendly alternative reagent for the Mitsunobu reaction. Chem. Lett. 2007, 36, 566–567. [Google Scholar] [CrossRef] [Green Version]
- Hagiya, K.; Muramoto, N.; Misaki, T.; Sugimura, T. DMEAD: A new dialkyl azodicarboxylate for the Mitsunobu reaction. Tetrahedron 2009, 65, 6109–6114. [Google Scholar] [CrossRef]
- Tamai, H.; Ando, H.; Tanaka, H.-N.; Hosoda-Yabe, R.; Yabe, T.; Ishida, H.; Kiso, M. The total synthesis of the neurogenic ganglioside LLG-3 isolated from the Starfish Linckia laevigata. Angew. Chem. Int. Ed. 2011, 50, 2330–2333. [Google Scholar] [CrossRef]
- Ishiwata, A.; Munemura, Y.; Ito, Y. Synergistic solvent effect in 1,2-cis-glycoside formation. Tetrahedron 2008, 64, 92–102. [Google Scholar] [CrossRef]
- Mandal, S.S.; Liao, G.; Guo, Z. Chemical synthesis of the tumor-associated Globo-H antigen. RSC Adv. 2015, 5, 23311–23319. [Google Scholar] [CrossRef] [Green Version]
- Kenworthy, A.K.; Nichols, B.J.; Remmert, C.L.; Hendirix, G.M.; Kumar, M.; Zimmerberg, J.; Lippincott-Schwartz, J. Dynamics of putative raft-associated proteins at the cell surface. J. Cell Biol. 2004, 165, 735–746. [Google Scholar] [CrossRef]
- Lingwood, D.; Ries, J.; Schwille, P.; Simons, K. Plasma membranes are poised for activation of raft phase coalescence at physiological temperature. Proc. Natl. Acad. Sci. USA 2008, 105, 10005–10010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgart, T.; Hammond, A.T.; Sengupta, P.; Hess, S.T.; Holowka, D.A.; Baird, B.A.; Webb, W.W. Large-scale fluid/fluid phase separation of proteins and lipids in giant plasma membrane vesicles. Proc. Natl. Acad. Sci. USA 2007, 104, 3165–3170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levental, I.; Grzybek, M.; Simons, K. Raft domains of variable properties and compositions in plasma membrane vesicles. Proc. Natl. Acad. Sci. USA 2011, 108, 11411–11416. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asano, S.; Pal, R.; Tanaka, H.-N.; Imamura, A.; Ishida, H.; Suzuki, K.G.N.; Ando, H. Development of Fluorescently Labeled SSEA-3, SSEA-4, and Globo-H Glycosphingolipids for Elucidating Molecular Interactions in the Cell Membrane. Int. J. Mol. Sci. 2019, 20, 6187. https://doi.org/10.3390/ijms20246187
Asano S, Pal R, Tanaka H-N, Imamura A, Ishida H, Suzuki KGN, Ando H. Development of Fluorescently Labeled SSEA-3, SSEA-4, and Globo-H Glycosphingolipids for Elucidating Molecular Interactions in the Cell Membrane. International Journal of Molecular Sciences. 2019; 20(24):6187. https://doi.org/10.3390/ijms20246187
Chicago/Turabian StyleAsano, Sachi, Rita Pal, Hide-Nori Tanaka, Akihiro Imamura, Hideharu Ishida, Kenichi G. N. Suzuki, and Hiromune Ando. 2019. "Development of Fluorescently Labeled SSEA-3, SSEA-4, and Globo-H Glycosphingolipids for Elucidating Molecular Interactions in the Cell Membrane" International Journal of Molecular Sciences 20, no. 24: 6187. https://doi.org/10.3390/ijms20246187
APA StyleAsano, S., Pal, R., Tanaka, H.-N., Imamura, A., Ishida, H., Suzuki, K. G. N., & Ando, H. (2019). Development of Fluorescently Labeled SSEA-3, SSEA-4, and Globo-H Glycosphingolipids for Elucidating Molecular Interactions in the Cell Membrane. International Journal of Molecular Sciences, 20(24), 6187. https://doi.org/10.3390/ijms20246187