Inducible Nitric Oxide Synthase (iNOS) Mediates Vascular Endothelial Cell Apoptosis in Grass Carp Reovirus (GCRV)-Induced Hemorrhage



Abstract
:
1. Introduction
2. Results
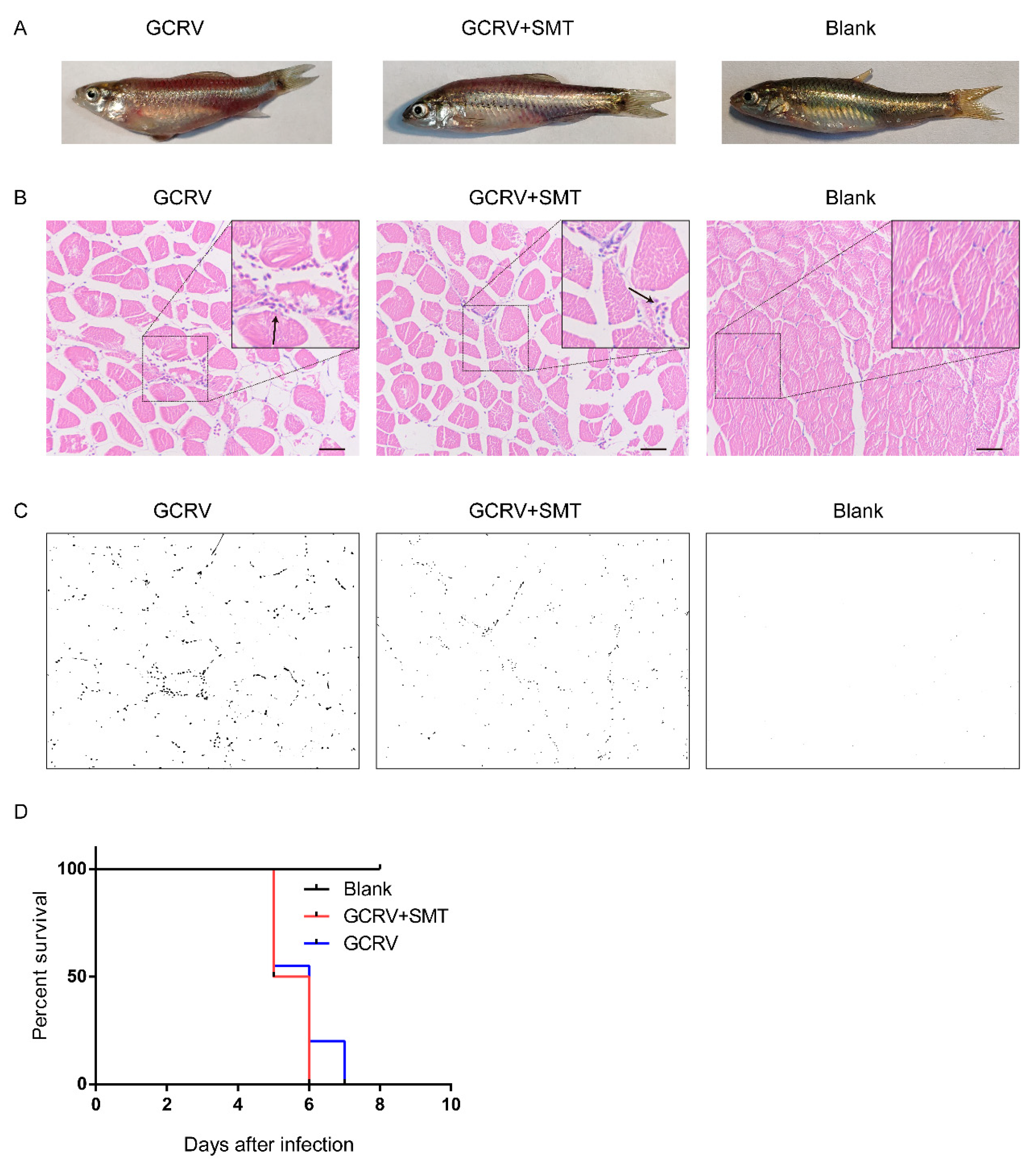
2.1. iNOS Relates to Hemorrhage Symptom Caused by GCRV Infection
2.2. iNOS Induces Cell Apoptosis
2.3. SMT Inhibits Cell Apoptosis Caused by GCRV Infection
2.4. SMT Suppresses the Function of iNOS in Vivo
2.5. The Hemorrhage is Reduced When Inhibiting the Function of iNOS
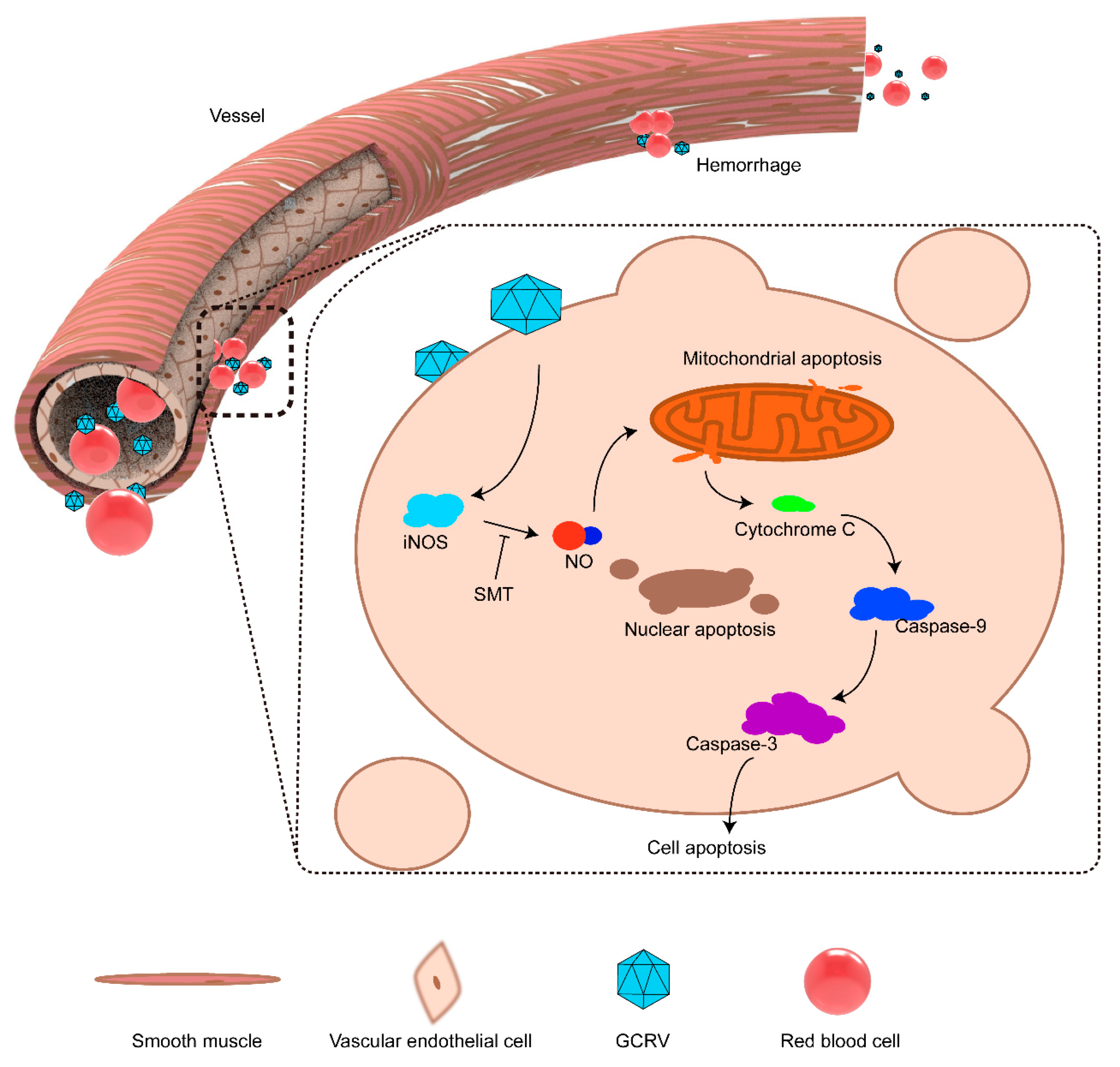
2.6. iNOS Breaks the Vascular Wall Via Activating Cell Apoptosis
2.7. iNOS Activates Both the Coagulation and the Anticoagulation Systems
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animal Experiments
4.3. Cloning of iNOS, Plasmids Construction, Cell Culture, and Transfection
4.4. GCRV Infection and SMT Inhibitor
4.5. Fluorescence Microscopy
4.6. Protein Extraction and Western Blot
4.7. Caspase Activity Assay, NO Measure, and Histopathology
4.8. qRT-PCR Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| bNOS | Bacterial NOS |
| eNOS | Endothelial NOS |
| FHM | Fathead Minnow |
| GCRV | Grass Carp Reovirus |
| iNOS | Inducible Nitric Oxide Synthase |
| kng-1 | Kininogen-1 |
| nNOS | Neuronal NOS |
| NO | Nitric Oxide |
| NOSs | Nitric Oxide Synthases |
| PBS | Phosphate Buffer Saline |
| pCCE | pCMV-3×Flag-CMV-eGFP |
| CICE | pCMV-iNOS-3×Flag-CMV-eGFP |
| PMSF | Phenylmethylsulfonyl fluoride |
| qRT-PCR | Quantitative real-time RT-PCR |
| serpin | Serine protease inhibitor |
| SMT | S-methylisothiourea sulfate |
| TBST | Tris-buffered saline tween-20 |
References
- Gavito-Higuera, J.; Mullins, C.B.; Ramos-Duran, L.; Chacon, C.I.O.; Hakim, N.; Palacios, E. Fungal infections of the central nervous system: A pictorial review. J. Clin. Imaging Sci. 2016, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Novotny, W.E.; Dixit, A. Pulmonary hemorrhage in an infant following 2 weeks of fungal exposure. Arch. Pediatr. Adolesc. Med. 2000, 154, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.M.; Zhou, Z.M.; Pan, X.Y. Study on the pathogen of red-skin disease of Aerossocheilus fasciatus. J. Shanghai Ocean Univ. 2010, 19, 631–634, (In Chinese with English abstract). [Google Scholar]
- Tekedar, H.C.; Karsi, A.; Akgul, A.; Kalindamar, S.; Waldbieser, G.C.; Sonstegard, T.; Schroeder, S.G.; Lawrence, M.L. Complete genome sequence of fish pathogen Aeromonas hydrophila AL06-06. Genome Announc. 2015, 3, 549–556. [Google Scholar] [CrossRef] [Green Version]
- Cosgriff, T.M.; Lewis, R.M. Mechanisms of disease in hemorrhagic fever with renal syndrome. Kidney Int. Suppl. 1991, 35, S72. [Google Scholar]
- Shcheblyakov, D.; Esmagambetov, I.; Simakin, P.; Kostina, L.; Kozlov, A.; Tsibezov, V.; Grebennikova, T.; Chifanov, D.; Rumyantseva, I.; Boyarskaya, N.; et al. Development and characterization of two GP-specific monoclonal antibodies, which synergistically protect non-human primates against Ebola lethal infection. Antivir. Res. 2019, 104617. [Google Scholar] [CrossRef]
- Du, F.; Su, J.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. Cloning and preliminary functional studies of the JAM-A gene in grass carp (Ctenopharyngodon idellus). Fish Shellfish Immunol. 2013, 34, 1476–1484. [Google Scholar] [CrossRef]
- Jiang, Y.L.; Liopo, G. Hemorrhagic disease of grass carp: Status of outbreaks, diagnosis, surveillance, and research. Isr. J. Aquac.-Bamidgeh 2009, 61, 188–197. [Google Scholar]
- Rao, Y.; Su, J. Insights into the antiviral immunity against grass carp (Ctenopharyngodon idella) reovirus (GCRV) in grass carp. J. Immunol. Res. 2015, 2015, 670437. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.; Shao, J.; Hang, Q.; Zhang, N. On pathogen of hemorrhage of grass carp (Ctenopharyngodon idellus C. ET V.). J. Fish. China 1989, 13, 1–4, (In Chinese with English abstract). [Google Scholar]
- Yan, C.; Xiao, J.; Li, J.; Chen, H.; Liu, J.; Wang, C.; Feng, C.; Feng, H. TBK1 of black carp plays an important role in host innate immune response against SVCV and GCRV. Fish Shellfish Immunol. 2017, 69, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, P.; Chen, H.; Liu, H.; Yi, Y.; Guo, W. Preliminary study on the susceptibility of Gobiocypris rarus to hemorrhagic virus of grass carp (GCHV). Acta Hydrobiol. Sin. 1994, 18, 144–149, (In Chinese with English abstract). [Google Scholar]
- Li, X.; Zeng, W.W.; Wang, Q.; Wang, Y.Y.; Li, Y.Y.; Shi, C.B.; Wu, S.Q. Progress on grass carp reovirus. Prog. Veter. Med. 2016, 37, 94–101, (In Chinese with English abstract). [Google Scholar]
- Wang, Q.; Zeng, W.; Liu, C.; Zhang, C.; Wang, Y.; Shi, C.; Wu, S. Complete genome sequence of a reovirus isolated from grass carp, indicating different genotypes of GCRV in China. J. Virol. 2012, 86, 12466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Q.; Su, J. Transcriptome analysis provides insights into the regulatory function of alternative splicing in antiviral immunity in grass carp (Ctenopharyngodon idella). Sci. Rep. 2015, 5, 12946. [Google Scholar] [CrossRef] [Green Version]
- Wan, Q.; Liao, Z.; Rao, Y.; Yang, C.; Ji, J.; Chen, X.; Su, J. Transferrin Receptor 1-Associated Iron Accumulation and Oxidative Stress Provides a Way for Grass Carp to Fight against Reovirus Infection. Int. J. Mol. Sci. 2019, 20, 5857. [Google Scholar] [CrossRef] [Green Version]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2011, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Oswald, I.P.; Eltoum, I.; Wynn, T.A.; Schwartz, B.; Caspar, P.; Paulin, D.; Sher, A.; James, S.L. Endothelial cells are activated by cytokine treatment to kill an intravascular parasite, Schistosoma mansoni, through the production of nitric oxide. Proc. Natl. Acad. Sci. USA 1994, 91, 999–1003. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.-w.; Cho, H.J.; Calaycay, J.; Mumford, R.A.; Swiderek, K.M.; Lee, T.D.; Ding, A.; Troso, T.; Nathan, C. Cloning and characterization of inducible nitric oxide synthase from mouse macrophages. Science 1992, 256, 225–228. [Google Scholar] [CrossRef]
- Gusarov, I.; Nudler, E. NO-mediated cytoprotection: Instant adaptation to oxidative stress in bacteria. Proc. Natl. Acad. Sci. USA 2005, 102, 13855–13860. [Google Scholar] [CrossRef] [Green Version]
- Nathan, C.F.; Hibbs, J.B. Role of nitric oxide synthesis in macrophage antimicrobial activity. Curr. Opin. Immunol. 1991, 3, 65–70. [Google Scholar] [CrossRef]
- Stuehr, D.J.; Griffith, O.W. Mammalian nitric oxide synthases. Biochim. Biophys. Acta Bioenerg. 1999, 1411, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907. [Google Scholar] [CrossRef] [PubMed]
- Cau, S.B.D.A.; Carneiro, F.S.; Tostes, R. Differential modulation of nitric oxide synthases in aging: Therapeutic opportunities. Front. Physiol. 2012, 3, 218. [Google Scholar] [CrossRef] [Green Version]
- Green, S.J.; Scheller, L.F.; Marletta, M.A.; Seguin, M.C.; Klotz, F.W.; Slayter, M.; Nelson, B.J.; Nacy, C.A. Nitric oxide: Cytokine-regulation of nitric oxide in host resistance to intracellular pathogens. Immunol. Lett. 1994, 43, 87. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Dempsey, S.K.; Daneva, Z.; Azam, M.; Li, N.; Li, P.-L.; Ritter, J.K. Role of nitric oxide in the cardiovascular and renal systems. Int. J. Mol. Sci. 2018, 19, 2605. [Google Scholar] [CrossRef] [Green Version]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol.-Cell Physiol. 1996, 271, 1424–1437. [Google Scholar] [CrossRef] [Green Version]
- Marla, S.S.; Lee, J.; Groves, J.T. Peroxynitrite rapidly permeates phospholipid membranes. Proc. Natl. Acad. Sci. USA 1997, 94, 14243–14248. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Ding, J.; Wang, J.; Zhou, C.; Zhang, W. Effects and mechanism of action of inducible nitric oxide synthase on apoptosis in a rat model of cerebral ischemia-reperfusion injury. Anat. Rec. 2016, 299, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.P. Nitric oxide and cell death. Biochim. Biophys. Acta Bioenerg. 1999, 1411, 401–414. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Lin, S.C.; Chen, W.Y.; Yen, Y.T.; Lai, C.W.; Tao, M.H.; Lin, Y.L.; Miaw, S.C.; Wu-Hsieh, B.A. Dengue viral protease interaction with NF-kappaB inhibitor alpha/beta results in endothelial cell apoptosis and hemorrhage development. J. Immunol. 2014, 193, 1258–1267. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, X. Cytochrome C-mediated apoptosis. Annu. Rev. Biochem. 2004, 73, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Miest, J.J.; Adamek, M.; Pionnier, N.; Harris, S.; Matras, M.; Rakus, K.Ł.; Irnazarow, I.; Steinhagen, D.; Hoole, D. Differential effects of alloherpesvirus CyHV-3 and rhabdovirus SVCV on apoptosis in fish cells. Vet. Microbiol. 2015, 176, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Sanjeev, P.; Richa, S.; Anshu, P. Overview of the coagulation system. Indian J. Anaesth. 2014, 58, 515–523. [Google Scholar]
- Katherine, H.; Michael, L.; Henderson, S.O. Anticoagulation drug therapy: A review. Western J. Emerg. Med. 2015, 16, 11. [Google Scholar]
- Furie, B.; Furie, B.C. Thrombus formation in vivo. J. Clin. Investig. 2005, 115, 3355. [Google Scholar] [CrossRef]
- Long, A.T.; Kenne, E.; Jung, R.; Fuchs, T.A.; Renné, T. Contact system revisited: An interface between inflammation, coagulation, and innate immunity. J. Thromb. Haemost. 2016, 14, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Di, N.M.; Middeldorp, S.; Büller, H.R. Direct thrombin inhibitors. Thromb. Res. 2005, 353, 1028. [Google Scholar]
- Triplett, D.A. Coagulation and bleeding disorders: Review and update. Clin. Chem. 2000, 46, 1260. [Google Scholar]
- Sergei, M.; Wan-Ming, Z.; Komar, A.A.; Schmaier, A.H.; Ellen, B.; Yihua, Z.; Xincheng, L.; Takayuki, I.; Castellino, F.J.; Guangbin, L. Deletion of murine kininogen gene 1 (mKng1) causes loss of plasma kininogen and delays thrombosis. Blood 2008, 111, 1274–1281. [Google Scholar]
- Van Steijn, J.H.; Van Tol, K.M.; Van Essen, L.H.; Gans, R.O. Disseminated intravascular coagulation as an unusual presentation of an Epstein-Barr virus infection. Neth. J. Med. 2000, 57, 169–171. [Google Scholar] [CrossRef]
- Chen, L.; Huang, R.; Zhu, D.; Wang, Y.; Mehjabin, R.; Li, Y.; Liao, L.; He, L.; Zhu, Z.; Wang, Y. Cloning of six serpin genes and their responses to GCRV infection in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2019, 86, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Van, R.J.; Stangier, J.; Haertter SLiesenfeld, K.H.; Wienen, W.; Feuring, M.; Clemens, A. Dabigatran etexilate--a novel, reversible, oral direct thrombin inhibitor: Interpretation of coagulation assays and reversal of anticoagulant activity. Thromb. Haemost. 2010, 103, 1116–1127. [Google Scholar]
- He, L.; Zhang, A.; Pei, Y.; Chu, P.; Li, Y.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. Differences in responses of grass carp to different types of grass carp reovirus (GCRV) and the mechanism of hemorrhage revealed by transcriptome sequencing. BMC Genom. 2017, 18, 452. [Google Scholar] [CrossRef]
- Su, H.; Su, J. Cyprinid viral diseases and vaccine development. Fish Shellfish Immunol. 2018, 83, 84–95. [Google Scholar] [CrossRef]
- Croen, K.D. Evidence for antiviral effect of nitric oxide. Inhibition of herpes simplex virus type 1 replication. J. Clin. Investig. 1993, 91, 2446–2452. [Google Scholar] [CrossRef]
- Kluck, R.M.; Bossy-Wetzel, E.; Green, D.R.; Newmeyer, D.D. The release of cytochrome c from mitochondria: A primary site for bcl-2 regulation of apoptosis. Science 1997, 275, 1132–1136. [Google Scholar] [CrossRef] [Green Version]
- Satake, K.; Matsuyama, Y.; Kamiya, M.; Kawakami, H.; Iwata, H.; Adachi, K.; Kiuchi, K. Nitric oxide via macrophage iNOS induces apoptosis following traumatic spinal cord injury. Mol. Brain Res. 2000, 85, 114–122. [Google Scholar] [CrossRef]
- Szabó, C.; Southan, G.J.; Thiemermann, C. Beneficial effects and improved survival in rodent models of septic shock with S-methylisothiourea sulfate, a potent and selective inhibitor of inducible nitric oxide synthase. Proc. Natl. Acad. Sci. USA 1994, 91, 12472–12476. [Google Scholar]
- Zheng, D.; Huang, Q.; Cai, W.; Lu, H. Histopathological studies on the hemorrage disease of grass carp. J. Fish. China 1986, 10, 151–159. [Google Scholar]
- Oztas, Y.; Uysal, B.; Kaldirim, U.; Poyrazoglu, Y.; Yasar, M.; Cayci, T.; Cekli, Y.; Sadir, S.; Ozler, M.; Topal, T. Inhibition of iNOS reduces the therapeutic effects of ozone in acute necrotizing pancreatitis: An in vivo animal study. Scand. J. Clin. Lab. Investig. 2011, 71, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Gravell, M.; Malsberger, R. A permanent cell line from the fathead minnow (Pimephales promelas). Ann. N. Y. Acad. Sci. 1965, 126, 555–565. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ → 3′) |
|---|---|
| NOS1F | CGTCAGATCGGAATTCGGTACCAATGGGGAACCAGGCCACTAAAGACA |
| NOS1R | GCTGCAAACTTATCGTCCACTGCGTGAG |
| NOS2F | CGCAGTGGACGATAAGTTTGCAGCACT |
| NOS2R | GTCAGCCCGGGATCCTCTAGAGGATCCGAAGATGTCCTCATGGTACCGC |
| Gene Name | Primer Name | Sequence (5′ → 3′) | Accession Number |
|---|---|---|---|
| Serpinb1 | Sb1F | TCTCTGATGTTCACTCTCGGTTTG | MN326820 |
| Sb1R | TCTTTGATTTTATTCTCCGTCTGC | ||
| Serpinc1 | Sc1F | TCGATAAAGGAGAAGACATCTGACC | MN326821 |
| Sc1R | GCTGGAAGGACTCATTAAAAAGTGTAG | ||
| Serpind1 | Sd1F | GCAGAACTATGACTTGATTGACCAC | MN326822 |
| Sd1R | TGATAAAGGCATGAATCCGATGT | ||
| Serpinf1 | Sf1F | CTCTCCAATACTCTCCACGCAGT | MN326823 |
| Sf1R | CGCTTCCAACCATTCAGACATAC | ||
| Serpinf2b | Sf2bF | ACCTTGATACACTCTGTGATGGGG | MN326824 |
| Sf2bR | GGAAATTATGACATTGGGCTGTT | ||
| Serping1 | Sg1F | TTCCTCTCAGACACTGTTGGTGG | MN326825 |
| Sg1R | GAAGATCAAGTTGGATTTGGGGT | ||
| iNOS | NOSF | CAGGTCTGAAAGGGAATCCTATGA | MN326826 |
| NOSR | TTGGGTTGTCCAGTCTGCCTAG | ||
| Kng-1 | Kng1F | ACCGCTCATTCATTCTTTGGC | MN380324 |
| Kng1R | CCTGGTGCAGTTGCTCTTCTC | ||
| F2 | F2F | GGACCTCAAACCCCACAAATC | MN380325 |
| F2R | CCCACGACACAATGCCAATC | ||
| F3a | F3aF | CAGGCCTACCTGCCCTTCC | MN380326 |
| F3aR | TTTTTCGCTCGCTTTCTTCG | ||
| F3b | F3bF | TTTTCGGTGCTTTTTCTCGC | MN380327 |
| F3bR | TCTGTTTGTCCCTACCAACTCTTG | ||
| F10 | F10F | AGTAAAGTTGCCACACAGGGAGC | MN380328 |
| F10R | CAGTTGGGTAGTTGTAAGAATGTAAAGG | ||
| β-actin | ACTF | AATTGTCCGTGACATCAAAGAGAA | MN562268 |
| ACTR | GATACCGCAAGATTCCATACCC | ||
| VP4 | VP4F | CGAAAACCTACCAGTGGATAATG | ADC81088 |
| VP4R | CCAGCTAATACGCCAACGAC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, B.; Su, J. Inducible Nitric Oxide Synthase (iNOS) Mediates Vascular Endothelial Cell Apoptosis in Grass Carp Reovirus (GCRV)-Induced Hemorrhage. Int. J. Mol. Sci. 2019, 20, 6335. https://doi.org/10.3390/ijms20246335
Liang B, Su J. Inducible Nitric Oxide Synthase (iNOS) Mediates Vascular Endothelial Cell Apoptosis in Grass Carp Reovirus (GCRV)-Induced Hemorrhage. International Journal of Molecular Sciences. 2019; 20(24):6335. https://doi.org/10.3390/ijms20246335
Chicago/Turabian StyleLiang, Bo, and Jianguo Su. 2019. "Inducible Nitric Oxide Synthase (iNOS) Mediates Vascular Endothelial Cell Apoptosis in Grass Carp Reovirus (GCRV)-Induced Hemorrhage" International Journal of Molecular Sciences 20, no. 24: 6335. https://doi.org/10.3390/ijms20246335
APA StyleLiang, B., & Su, J. (2019). Inducible Nitric Oxide Synthase (iNOS) Mediates Vascular Endothelial Cell Apoptosis in Grass Carp Reovirus (GCRV)-Induced Hemorrhage. International Journal of Molecular Sciences, 20(24), 6335. https://doi.org/10.3390/ijms20246335