Septoria Leaf Blotch and Reduced Nitrogen Availability Alter WRKY Transcription Factor Expression in a Codependent Manner

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
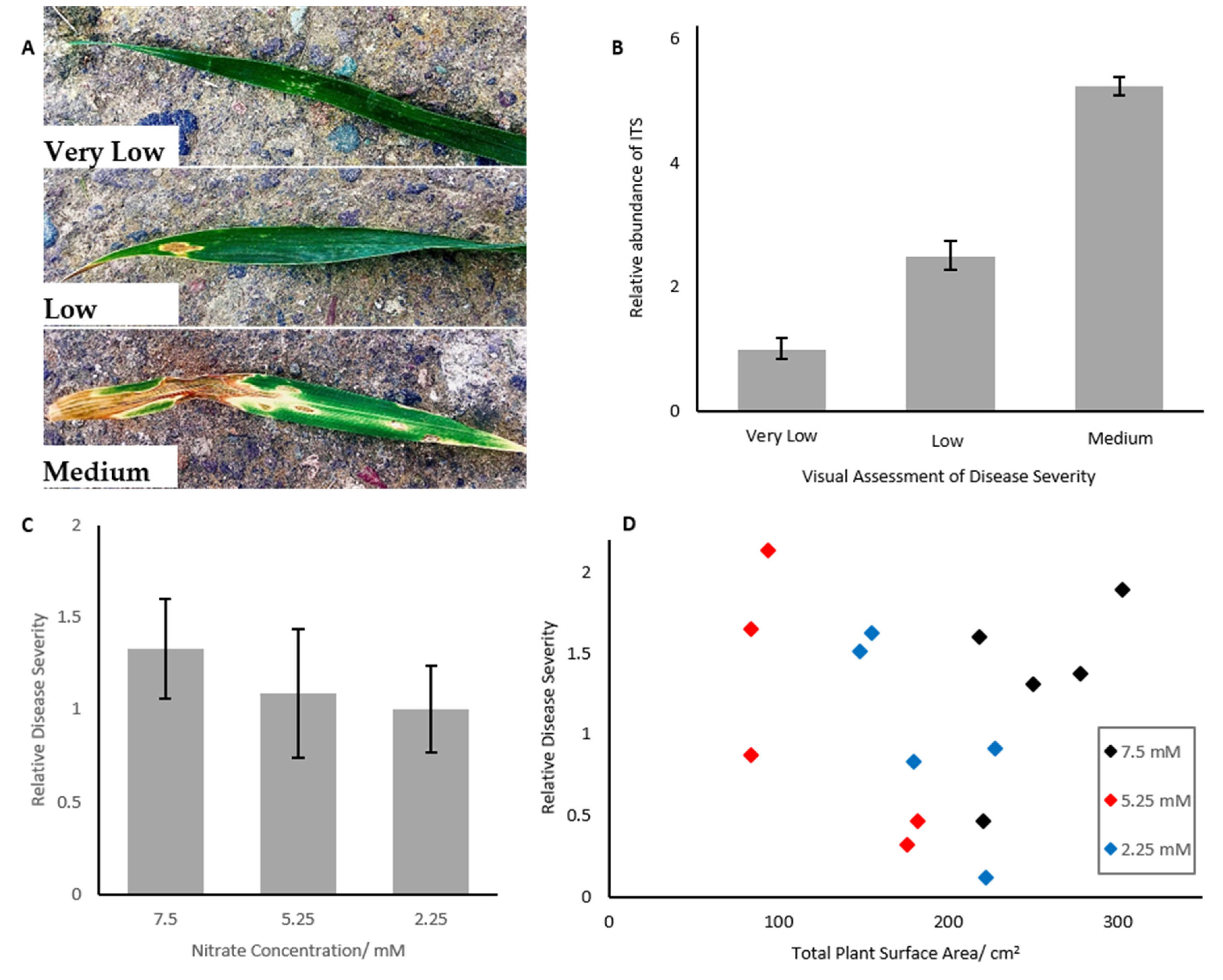
2.1. Septoria Abundance
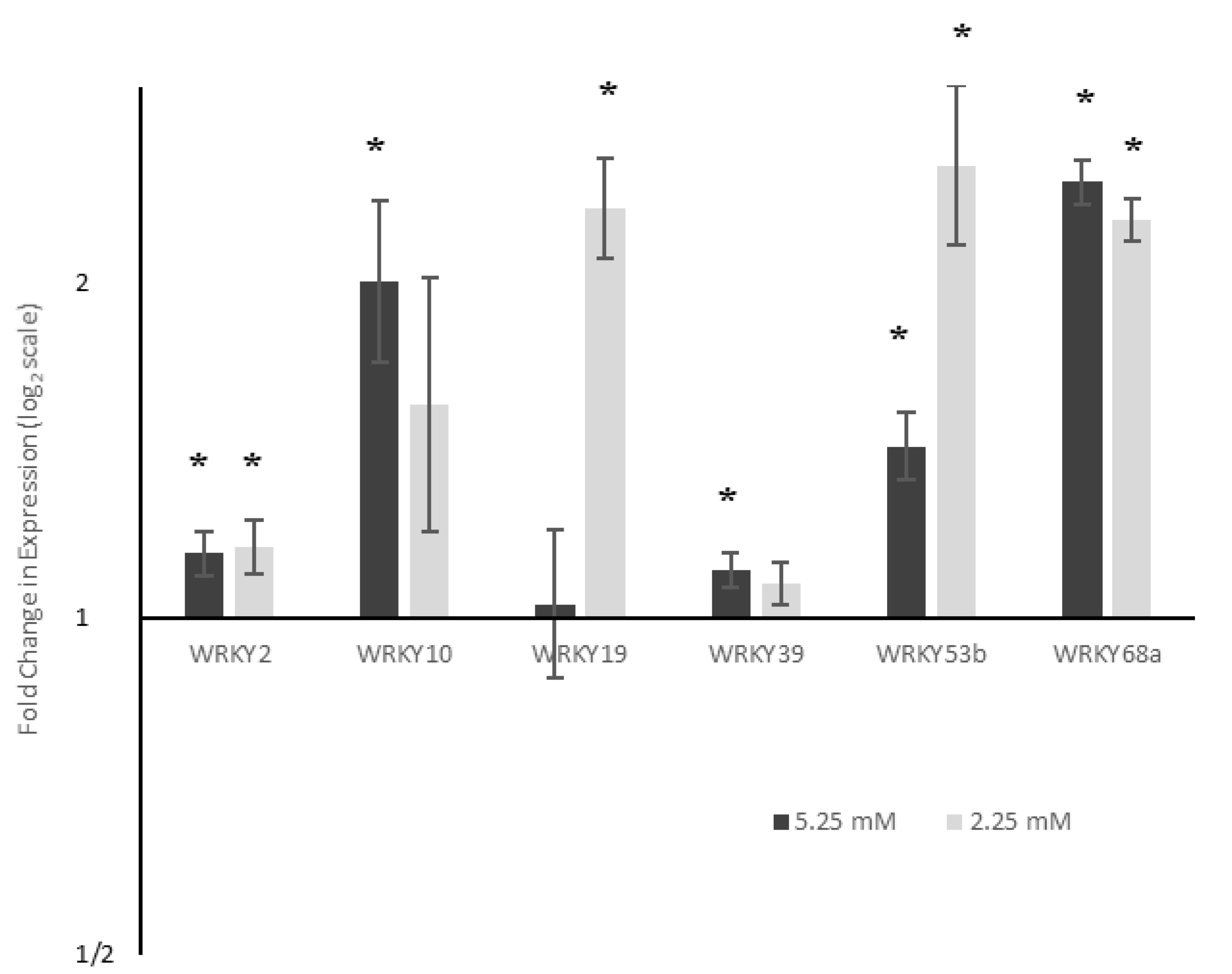
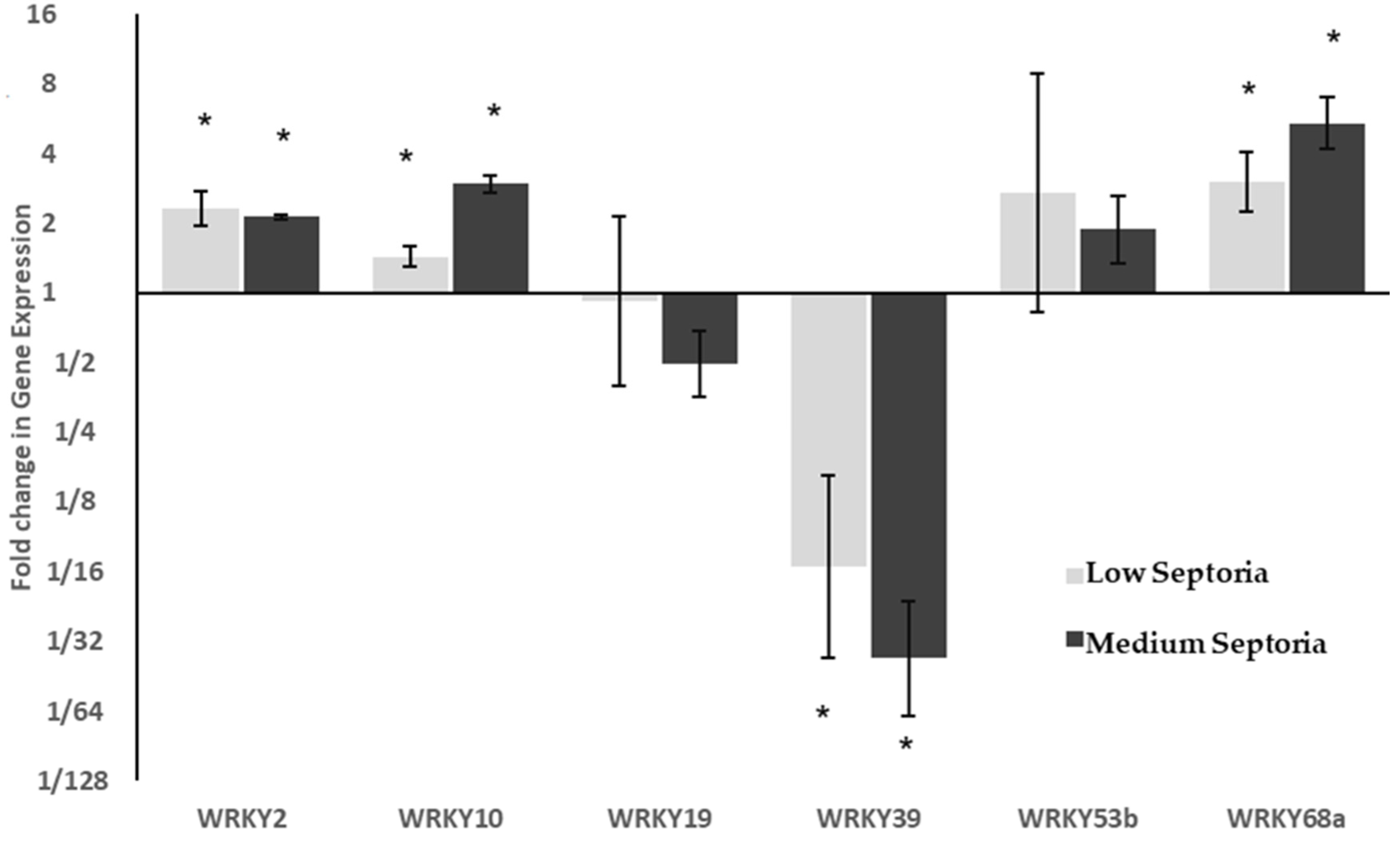
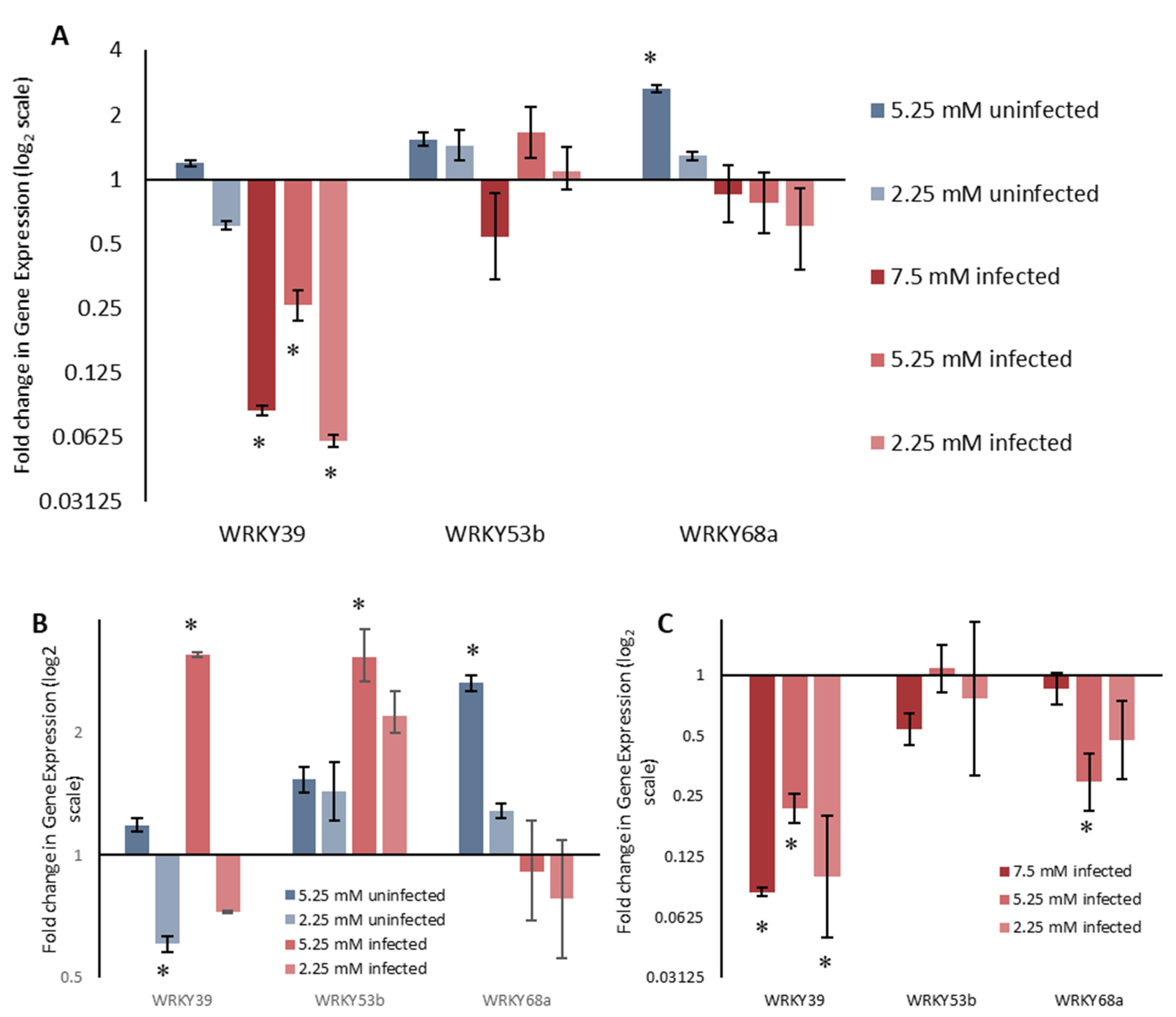
2.2. WRKY Expression
3. Discussion
4. Materials and Methods
4.1. Plant Growth Conditions and Physiological Assessment
4.2. Septoria Infection
4.3. Tissue Collection from Field Samples
4.4. RNA Extraction and cDNA Synthesis
4.5. Primer Design
4.6. qPCR
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| C | Celsius |
| cDNA | Complementary DNA |
| EF1α | Elongation factor 1 alpha |
| ITS | Internally transcribed region |
| L | Litre |
| M | Molar |
| PCR | Polymerase chain reaction |
| SLB | Septoria leaf blotch |
References
- FAOSTAT. Crop Production. 2017. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 10 June 2020).
- Valin, H.; Sands, R.; Van Der Mensbrugghe, D.; Nelson, G.C.; Ahammad, H.; Blanc, É.; Bodirsky, B.; Fujimori, S.; Hasegawa, T.; Havlik, P.; et al. The future of food demand: Understanding differences in global economic models. Agric. Econ. 2013, 45, 51–67. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef]
- Fones, H.N.; Gurr, S.J. The impact of Septoria Tritici Blotch disease on wheat: An EU perspective. Fungal Genet. Biol. 2015, 79, 3–7. [Google Scholar] [CrossRef]
- AHDB. Septoria Tritici in Winter Wheat Topic Sheet. 2012, Volume 2017. Available online: https://ahdb.org.uk/septoriatritici (accessed on 10 June 2020).
- Duncan, K.E.; Howard, R.J. Cytological analysis of wheat infection by the leaf blotch pathogen Mycosphaerella graminicola. Mycol. Res. 2000, 104, 1074–1082. [Google Scholar] [CrossRef]
- Goodwin, S.B.; Ben M’Barek, S.; Dhillon, B.; Wittenberg, A.H.J.; Crane, C.F.; Hane, J.K.; Foster, A.J.; Van Der Lee, T.A.J.; Grimwood, J.; Aerts, A.; et al. Finished genome of the fungal wheat pathogen Mycosphaerella graminicola reveals dispensome structure, chromosome plasticity, and stealth pathogenesis. PLoS Genet. 2011, 7. [Google Scholar] [CrossRef]
- Keon, J.; Antoniw, J.; Carzaniga, R.; Deller, S.; Ward, J.L.; Baker, J.M.; Beale, M.H.; Hammond-Kosack, K.E.; Rudd, J.J. Transcriptional adaptation of Mycosphaerella Graminicola to programmed cell death (PCD) of its susceptible wheat host. Mol. Plant Microbe Interact. 2007, 20, 178–193. [Google Scholar] [CrossRef]
- Henze, M.; Beyer, M.; Klink, H.; Verreet, J.-A. Characterizing meteorological scenarios favorable for Septoria tritici infections in wheat and estimation of latent periods. Plant Dis. 2007, 91, 1445–1449. [Google Scholar] [CrossRef]
- Castro, A.C.; Fleitas, M.C.; Schierenbeck, M.; Gerard, G.S.; Simón, M.R. Evaluation of different fungicides and nitrogen rates on grain yield and bread-making quality in wheat affected by Septoria Tritici Blotch and yellowspot. J. Cereal Sci. 2018, 83, 49–57. [Google Scholar] [CrossRef]
- Simon, M.R.; Cordo, C.A.; Perelló, A.E.; Struik, P.C. Influence of nitrogen supply on the susceptibility of wheat to Septoria tritici. J. Phytopathol. 2003, 151, 283–289. [Google Scholar] [CrossRef]
- Loyce, C.; Meynard, J.; Bouchard, C.; Rolland, B.; Lonnet, P.; Bataillon, P.; Bernicot, M.; Bonnefoy, M.; Charrier, X.; Debote, B.; et al. Interaction between cultivar and crop management effects on winter wheat diseases, lodging, and yield. Crop. Prot. 2008, 27, 1131–1142. [Google Scholar] [CrossRef]
- Prew, R.D.; Church, B.M.; Dewar, A.M.; Lacey, J.; Penny, A.; Plumb, R.T.; Thorne, G.N.; Todd, A.D.; Williams, T.D. Effects of eight factors on the growth and nutrient uptake of winter wheat and on the incidence of pests and diseases. J. Agric. Sci. 1983, 100, 363–382. [Google Scholar] [CrossRef]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Okay, S.; Derelli, E.; Unver, T. Transcriptome-wide identification of bread wheat WRKY transcription factors in response to drought stress. Mol. Genet. Genom. 2014, 289, 765–781. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.-X.; Tian, Y.; Han, L.; Yang, X. Constitutive expression of a salinity-induced wheat WRKY transcription factor enhances salinity and ionic stress tolerance in transgenic Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2013, 441, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Deng, P.; Chen, L.; Wang, X.; Ma, H.; Hu, W.; Yao, N.; Feng, Y.; Chai, R.; Yang, G.; et al. A wheat WRKY transcription factor TaWRKY10 confers tolerance to multiple abiotic stresses in transgenic tobacco. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Wang, J.; Song, N.; Li, M.; Cheng, Q.; Huang, G.; Guo, Y.; Fu, Y.; Xie, C.; Sun, Q.; et al. TaWRKY68 responses to biotic stresses are revealed by the orthologous genes from major cereals. Genet. Mol. Biol. 2014, 37, 73–80. [Google Scholar] [CrossRef]
- Deslandes, L.; Olivier, J.; Peeters, N.; Feng, D.X.; Khounlotham, M.; Boucher, C.; E Somssich, I.; Genin, S.; Marco, Y. Physical interaction between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus. Proc. Natl. Acad. Sci. USA 2003, 100, 8024–8029. [Google Scholar] [CrossRef]
- Liu, X.; Inoue, H.; Hayashi, N.; Jiang, C.-J.; Takatsuji, H. CC-NBS-LRR-type R proteins for rice blast commonly interact with specific WRKY transcription factors. Plant Mol. Biol. Rep. 2015, 34, 533–537. [Google Scholar] [CrossRef]
- Yang, F.; Li, W.; Jørgensen, H.J.L. Transcriptional reprogramming of wheat and the hemibiotrophic pathogen Septoria tritici during two phases of the compatible interaction. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Rudd, J.J.; Kanyuka, K.; Hassani-Pak, K.; Derbyshire, M.C.; Andongabo, A.; Devonshire, J.; Lysenko, A.; Saqi, M.; Desai, N.M.; Powers, S.J.; et al. Transcriptome and metabolite profiling of the infection cycle of Zymoseptoria tritici on wheat reveals a biphasic interaction with plant immunity involving differential pathogen chromosomal contributions and a variation on the hemibiotrophic lifestyle definition. Plant Physiol. 2015, 167, 1158–1185. [Google Scholar] [CrossRef]
- Lee, J.; Orosa, B.; Millyard, L.; Edwards, M.; Kanyuka, K.; Gatehouse, A.; Rudd, J.J.; Hammond-Kosack, K.E.; Pain, N.; Sadanandom, A.; et al. Functional analysis of a Wheat Homeodomain protein, TaR1, reveals that host chromatin remodelling influences the dynamics of the switch to necrotrophic growth in the phytopathogenic fungus Zymoseptoria tritici. New Phytol. 2015, 206, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, R.; Jiang, S.-Y.; Kumar, N.; Venkatesh, P.N.; Ramachandran, S. A comprehensive transcriptional profiling of the WRKY gene family in rice under various abiotic and phytohormone treatments. Plant Cell Physiol. 2008, 49, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Vinod, K.; Zheng, Z.; Fan, B.; Chen, Z. Roles of Arabidopsis WRKY3 and WRKY4 transcription factors in plant responses to pathogens. BMC Plant Biol. 2008, 8, 68. [Google Scholar] [CrossRef]
- Gao, H.; Wang, Y.; Xu, P.; Zhang, Z. Overexpression of a WRKY transcription factor TaWRKY2 enhances drought stress tolerance in transgenic wheat. Front. Plant Sci. 2018, 9, 997. [Google Scholar] [CrossRef]
- Shi, W.-Y.; Du, Y.-T.; Ma, J.; Min, D.-H.; Jin, L.-G.; Chen, J.; Chen, M.; Zhou, Y.-B.; Ma, Y.; Xu, Z.-S.; et al. The WRKY transcription factor GmWRKY12 confers drought and salt tolerance in soybean. Int. J. Mol. Sci. 2018, 19, 4087. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, M.; Oelmüller, R. WRKY transcription factors: Jack of many trades in plants. Plant Signal. Behav. 2014, 9. [Google Scholar] [CrossRef]
- Weßling, R.; Panstruga, R. Rapid quantification of plant-powdery mildew interactions by qPCR and conidiospore counts. Plant Methods 2012, 8, 35. [Google Scholar] [CrossRef]
- Cunfer, B.M.; Ueng, P.P. Taxonomy and identification of Septoria and Stagonospora species on small-grain cereals. Annu. Rev. Phytopathol. 1999, 37, 267–284. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Kema, G.H.J.; Groenewald, J.; Verkley, G.; Seifbarghi, S.; Razavi, M.; Gohari, A.M.; Mehrabi, R.; Crous, P. Zymoseptoria gen. nov.: A new genus to accommodate Septoria-like species occurring on graminicolous hosts. Pers. Mol. Phylogeny Evol. Fungi 2011, 26, 57–69. [Google Scholar] [CrossRef]
- Howard, D.D.; Chambers, A.Y.; Logan, J. Nitrogen and fungicide effects on yield components and disease severity in wheat. J. Prod. Agric. 1994, 7, 448–454. [Google Scholar] [CrossRef]
- Bent, A.; Mackey, D. Elicitors, effectors, andRGenes: The new paradigm and a lifetime supply of questions. Annu. Rev. Phytopathol. 2007, 45, 399–436. [Google Scholar] [CrossRef]
- Ma, X.; Keller, B.; McDonald, B.A.; Palma-Guerrero, J.; Wicker, T. Comparative transcriptomics reveals how wheat responds to infection by Zymoseptoria tritici. Mol. Plant Microbe Interact. 2018, 31, 420–431. [Google Scholar] [CrossRef]
- Kim, Y.; Park, S.; Gilmour, S.J.; Deits, T. Roles of CAMTA transcription factors and salicylic acid in configuring the low-temperature transcriptome and freezing tolerance of Arabidopsis. Plant J. 2013, 75, 364–376. [Google Scholar] [CrossRef] [PubMed]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Jagodzik, P.; Tajdel-Zielinska, M.; Cieśla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- El Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Ahmad, M. Overexpression of AtWRKY30 transcription factor enhances heat and drought stress tolerance in wheat (Triticum aestivum L.). Genes 2019, 10, 163. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Saari, E.E.; Prescott, J.M. A scale for appraising the foliar intensity of wheat diseases. Plant Dis. Rep. 1975, 59, 4. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poll, A.A.; Lee, J.; Sanderson, R.A.; Byrne, E.; Gatehouse, J.A.; Sadanandom, A.; Gatehouse, A.M.R.; Edwards, M.G. Septoria Leaf Blotch and Reduced Nitrogen Availability Alter WRKY Transcription Factor Expression in a Codependent Manner. Int. J. Mol. Sci. 2020, 21, 4165. https://doi.org/10.3390/ijms21114165
Poll AA, Lee J, Sanderson RA, Byrne E, Gatehouse JA, Sadanandom A, Gatehouse AMR, Edwards MG. Septoria Leaf Blotch and Reduced Nitrogen Availability Alter WRKY Transcription Factor Expression in a Codependent Manner. International Journal of Molecular Sciences. 2020; 21(11):4165. https://doi.org/10.3390/ijms21114165
Chicago/Turabian StylePoll, Alistair A., Jack Lee, Roy A. Sanderson, Ed Byrne, John A. Gatehouse, Ari Sadanandom, Angharad M. R. Gatehouse, and Martin G. Edwards. 2020. "Septoria Leaf Blotch and Reduced Nitrogen Availability Alter WRKY Transcription Factor Expression in a Codependent Manner" International Journal of Molecular Sciences 21, no. 11: 4165. https://doi.org/10.3390/ijms21114165
APA StylePoll, A. A., Lee, J., Sanderson, R. A., Byrne, E., Gatehouse, J. A., Sadanandom, A., Gatehouse, A. M. R., & Edwards, M. G. (2020). Septoria Leaf Blotch and Reduced Nitrogen Availability Alter WRKY Transcription Factor Expression in a Codependent Manner. International Journal of Molecular Sciences, 21(11), 4165. https://doi.org/10.3390/ijms21114165