Functional Recognition by CD8+ T Cells of Epitopes with Amino Acid Variations Outside Known MHC Anchor or T Cell Receptor Recognition Residues



Abstract
:1. Introduction
2. Results
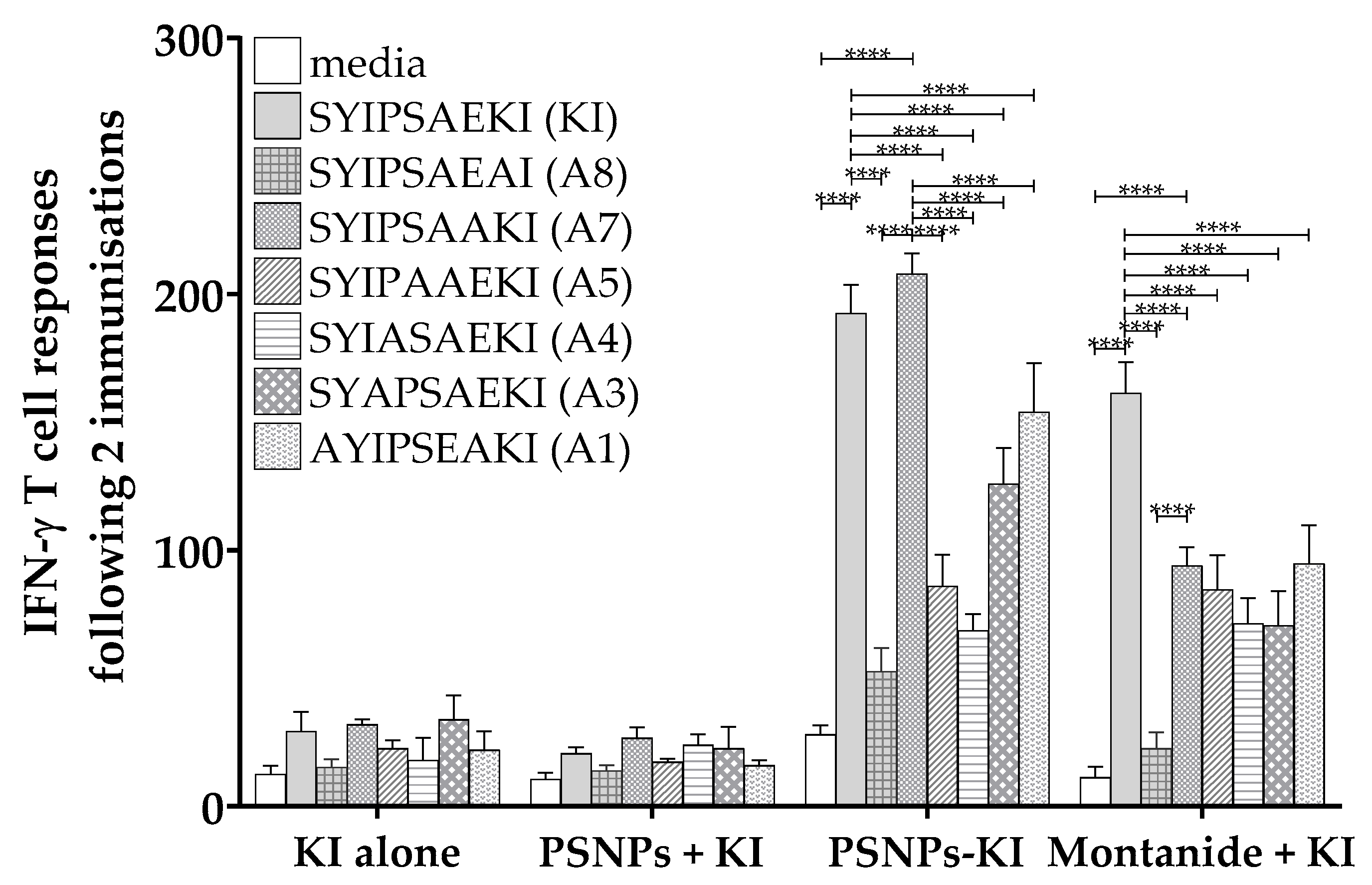
2.1. Peptide Antigen Delivery via Conjugation to Nanoparticles (PSNPs) Preserves the Moderate Cross-Reactivity to Alanine Altered Peptide Variants of SYIPSAEKI
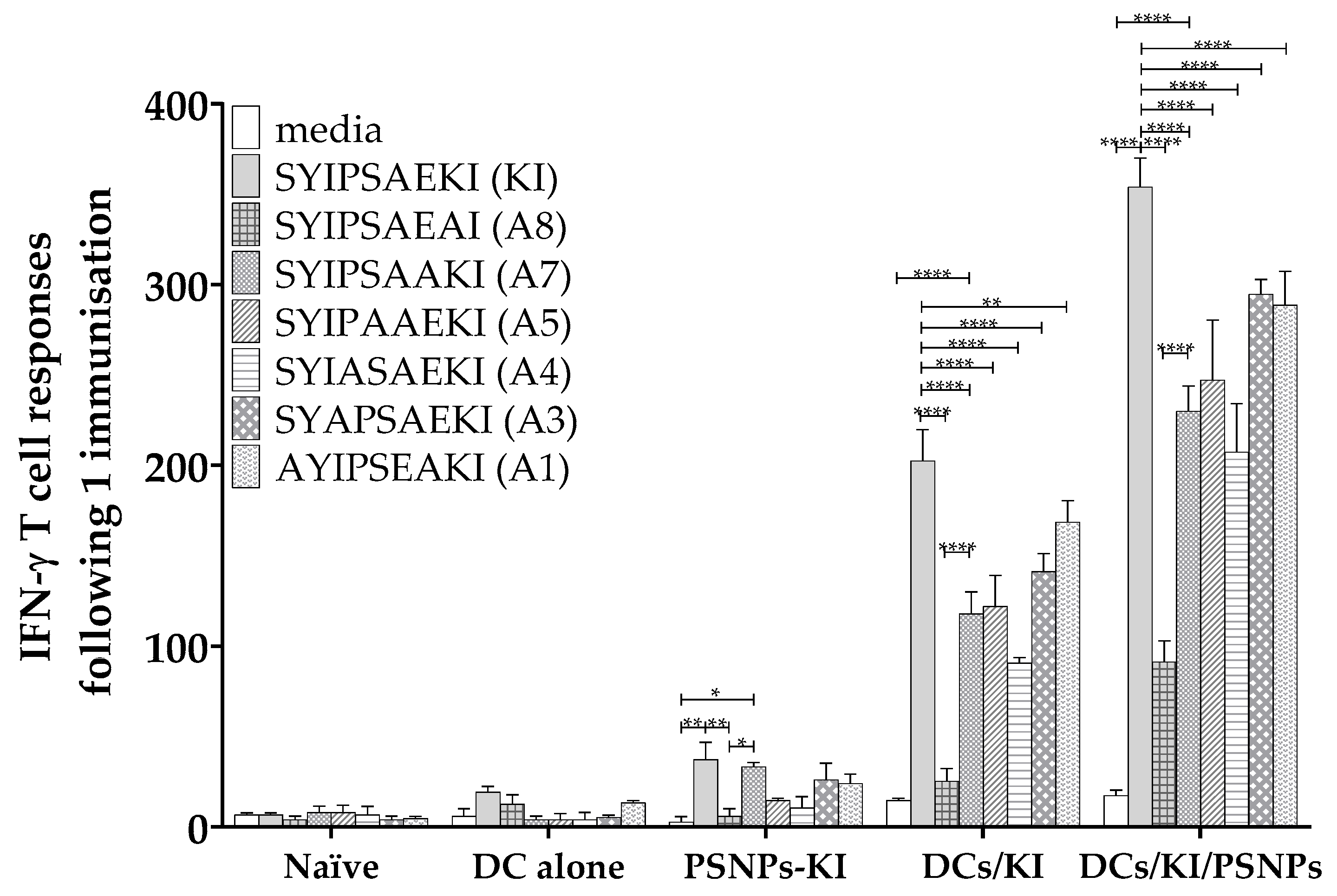
2.2. Ex Vivo Peptide Pulsed DCs Induce Potent T Cell Responses and Moderate Restimulated Cross-Reactive Responses to Alanine Screened Peptide Variants
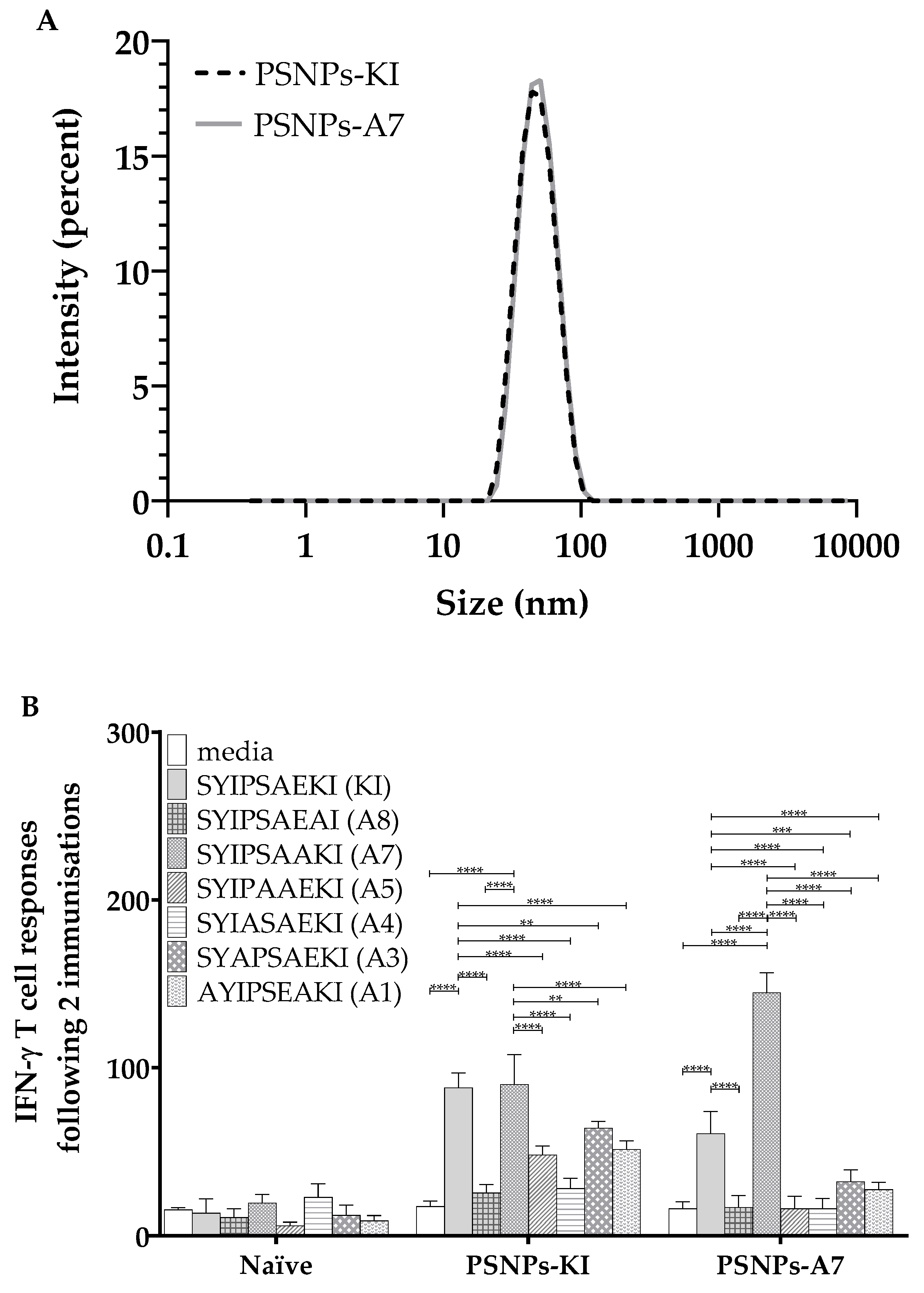
2.3. PSNPs Conjugated Peptide Variants (KI or A7) Elicit Peptide Specific IFN-γ T Cell Responses and Bi-Directional Cross-Reactivity
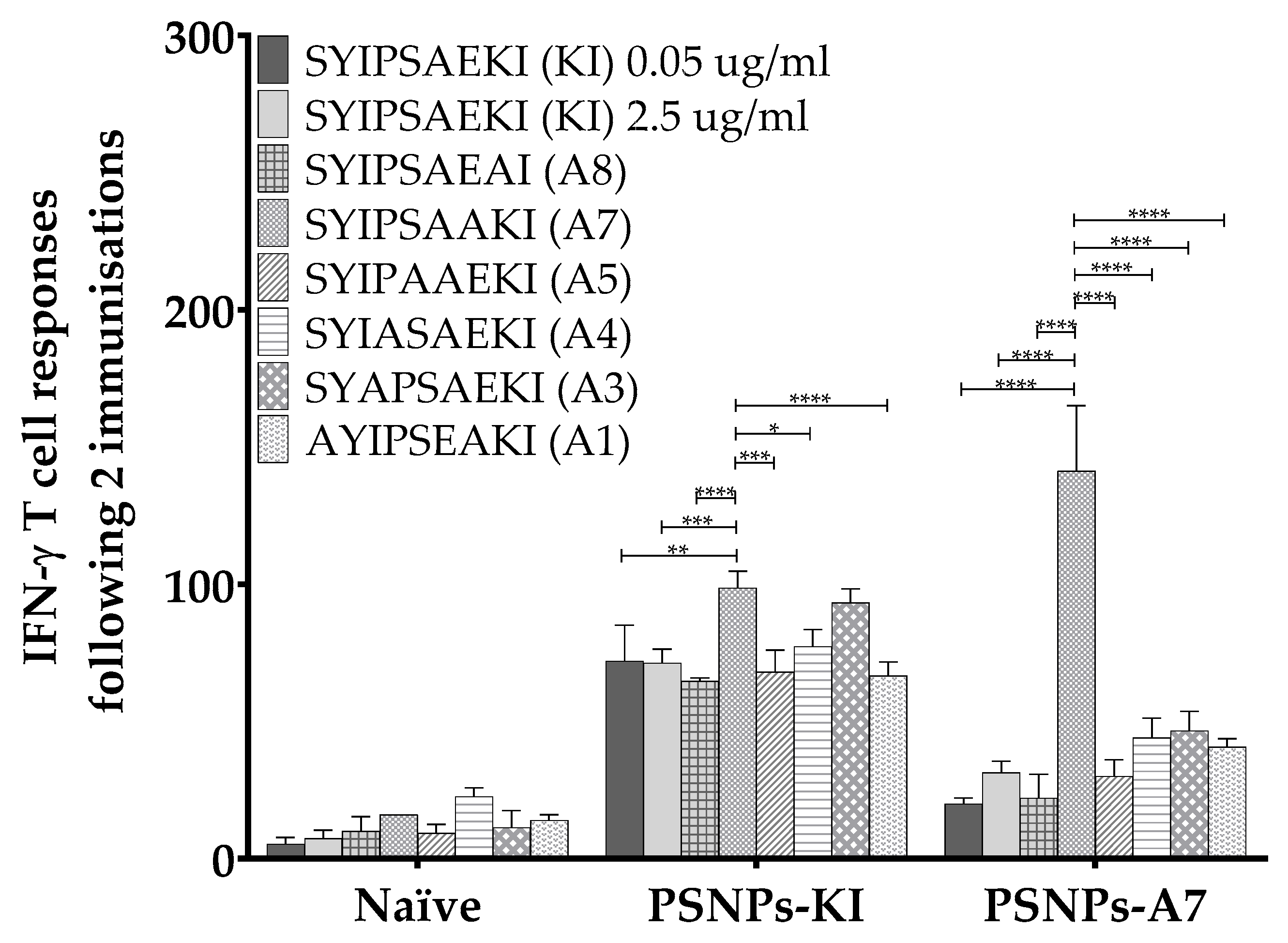
2.4. In Vitro Restimulation of Immunized Splenocytes with SYIPSAEKI co-Incubated with the Alanine Variants Together Shows no Evidence of Antagonistic Responses
3. Discussion
3.1. Influence of Conjugation on Peptide Responses
3.2. Differential Uptake of Conjugated vs. Soluble Peptides
3.3. Peptide Recognition by the TCR
3.4. Influence of Each Amino Acid Position along the Peptide
4. Materials and Methods
4.1. Animals
4.2. Peptides
4.3. Peptide Conjugation to PSNPs
4.4. Ex vivo DC Peptide Pulsing
4.5. Vaccines and Immunizations
4.6. ELISpot Assay
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| A1 | AYIPSAEKI peptide |
| A3 | SYAPSAEKI peptide |
| A4 | SYIASAEKI peptide |
| A5 | SYIPAAEKI peptide |
| A7 | SYIPSAAKI peptide |
| A8 | SYIPSAEAI peptide |
| AEC | Animal Ethics Committee |
| AMREP | Alfred Medical Research and Education Precinct |
| APCs | Antigen presenting cells |
| CD | Cluster of differentiation |
| ChAd | Chimpanzee adenoviral |
| CO2 | Carbon dioxide |
| DCs | Dendritic cells |
| EDC | 1-Ethyl-3-(3-dimethylaminopropyl)carbodiimide |
| FBS | Fetal bovine serum |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| IFN-y | Interferon gamma |
| IL | Interleukin |
| ISCOMs | Immune stimulating complexes |
| kDa | Kilodalton |
| KI | SYIPSAEKI peptide |
| MDSCs | Myeloid derived suppressor cells |
| MES | 2-(N-morpholino) ethanesulfonic acid |
| MHC | Major histocompatibility complex |
| MVA | Modified vaccinia Ankara |
| NP | Nucleoprotein |
| PBS | Phosphate buffered saline |
| PSNPs | Polystyrene nanoparticles |
| RT | Room temperature |
| SD | Standard deviation |
| SFU | Spot forming unit |
| TCR | T cell receptor |
| TNFR2 | Tumour necrosis factor receptor 2 |
| VLP | Virus like particle |
References
- Ko, E.J.; Kang, S.M. Immunology and efficacy of mf59-adjuvanted vaccines. Hum. Vaccines Immunother. 2018, 14, 3041–3045. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.B.; Haensler, J. An update on safety and immunogenicity of vaccines containing emulsion-based adjuvants. Expert Rev. Vaccines 2013, 12, 747–758. [Google Scholar] [CrossRef]
- Ewer, K.J.; Lambe, T.; Rollier, C.S.; Spencer, A.J.; Hill, A.V.; Dorrell, L. Viral vectors as vaccine platforms: From immunogenicity to impact. Curr. Opin. Immunol. 2016, 41, 47–54. [Google Scholar] [CrossRef]
- Huber, A.; Dammeijer, F.; Aerts, J.; Vroman, H. Current state of dendritic cell-based immunotherapy: Opportunities for in vitro antigen loading of different dc subsets? Front. Immunol. 2018, 9, 2804. [Google Scholar] [CrossRef]
- Aucouturier, J.; Dupuis, L.; Deville, S.; Ascarateil, S.; Ganne, V. Montanide isa 720 and 51: A new generation of water in oil emulsions as adjuvants for human vaccines. Expert Rev. Vaccines 2002, 1, 111–118. [Google Scholar] [CrossRef]
- Herrera, S.; Fernandez, O.L.; Vera, O.; Cardenas, W.; Ramirez, O.; Palacios, R.; Chen-Mok, M.; Corradin, G.; Arevalo-Herrera, M. Phase i safety and immunogenicity trial of plasmodium vivax cs derived long synthetic peptides adjuvanted with montanide isa 720 or montanide isa 51. Am. J. Trop. Med. Hyg. 2011, 84, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arevalo-Herrera, M.; Vera, O.; Castellanos, A.; Cespedes, N.; Soto, L.; Corradin, G.; Herrera, S. Preclinical vaccine study of plasmodium vivax circumsporozoite protein derived-synthetic polypeptides formulated in montanide isa 720 and montanide isa 51 adjuvants. Am. J. Trop. Med. Hyg. 2011, 84, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy-Thompson, E.; King, L.K.; Nunnley, L.D.; Mullins, I.M.; Slingluff, C.L., Jr.; Mullins, D.W. Peptide vaccination in montanide adjuvant induces and gm-csf increases cxcr3 and cutaneous lymphocyte antigen expression by tumor antigen-specific cd8 t cells. Cancer Immunol. Res. 2013, 1, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zha, Y.; Choudhury, N.; Malnassy, G.; Fulton, N.; Green, M.; Park, J.H.; Nakamura, Y.; Larson, R.A.; Salazar, A.M.; et al. Wt1 peptide vaccine in montanide in contrast to poly iclc, is able to induce wt1-specific immune response with tcr clonal enrichment in myeloid leukemia. Exp. Hematol. Oncol. 2018, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, E.; Liu, H.; Huckriede, A.; Hak, E. Safety and tolerability evaluation of the use of montanide isa51 as vaccine adjuvant: A systematic review. Hum. Vaccines Immunother. 2016, 12, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Chen, S.; Zhang, J. Bioinspired and biomimetic nanotherapies for the treatment of infectious diseases. Front. Pharmacol. 2019, 10, 751. [Google Scholar] [CrossRef]
- Powles, L.; Xiang, S.D.; Selomulya, C.; Plebanski, M. The use of synthetic carriers in malaria vaccine design. Vaccines 2015, 3, 894–929. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.D.; Gao, Q.; Wilson, K.L.; Heyerick, A.; Plebanski, M. A nanoparticle based sp17 peptide vaccine exposes new immuno-dominant and species cross-reactive b cell epitopes. Vaccines 2015, 3, 875–893. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.D.; Wilson, K.L.; Goubier, A.; Heyerick, A.; Plebanski, M. Design of peptide-based nanovaccines targeting leading antigens from gynecological cancers to induce hla-a2.1 restricted cd8(+) t cell responses. Front. Immunol. 2018, 9, 2968. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.D.; Wilson, K.; Day, S.; Fuchsberger, M.; Plebanski, M. Methods of effective conjugation of antigens to nanoparticles as non-inflammatory vaccine carriers. Methods 2013, 60, 232–241. [Google Scholar] [CrossRef]
- Wilson, K.L.; Pouniotis, D.; Hanley, J.; Xiang, S.D.; Ma, C.; Coppel, R.L.; Plebanski, M. A synthetic nanoparticle based vaccine approach targeting msp4/5 is immunogenic and induces moderate protection against murine blood-stage malaria. Front. Immunol. 2019, 10, 331. [Google Scholar] [CrossRef] [Green Version]
- Eberl, G.; Sabbatini, A.; Servis, C.; Romero, P.; Maryanski, J.L.; Corradin, G. Mhc class i h-2kd-restricted antigenic peptides: Additional constraints for the binding motif. Int. Immunol. 1993, 5, 1489–1492. [Google Scholar] [CrossRef]
- Katsara, M.; Minigo, G.; Plebanski, M.; Apostolopoulos, V. The good, the bad and the ugly: How altered peptide ligands modulate immunity. Expert Opin. Biol. Ther. 2008, 8, 1873–1884. [Google Scholar] [CrossRef]
- Plebanski, M.; Lee, E.A.; Hill, A.V. Immune evasion in malaria: Altered peptide ligands of the circumsporozoite protein. Parasitology 1997, 115, S55–S66. [Google Scholar] [CrossRef]
- Flanagan, K.L.; Wilson, K.L.; Plebanski, M. Polymorphism in liver-stage malaria vaccine candidate proteins: Immune evasion and implications for vaccine design. Expert Rev. Vaccines 2016, 15, 389–399. [Google Scholar] [CrossRef]
- Minigo, G.; Flanagan, K.L.; Slattery, R.M.; Plebanski, M. Vaccination with altered peptide ligands of a plasmodium berghei circumsporozoite protein cd8 t-cell epitope: A model to generate t cells resistant to immune interference by polymorphic epitopes. Front. Immunol. 2017, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Plebanski, M.; Lee, E.A.; Hannan, C.M.; Flanagan, K.L.; Gilbert, S.C.; Gravenor, M.B.; Hill, A.V. Altered peptide ligands narrow the repertoire of cellular immune responses by interfering with t-cell priming. Nat. Med. 1999, 5, 565–571. [Google Scholar] [CrossRef]
- Wilson, K.L.; Xiang, S.D.; Plebanski, M. Montanide, poly i:C and nanoparticle based vaccines promote differential suppressor and effector cell expansion: A study of induction of cd8 t cells to a minimal plasmodium berghei epitope. Front. Microbiol. 2015, 6, 29. [Google Scholar] [CrossRef]
- Wilson, K.L.; Xiang, S.D.; Plebanski, M. A model to study the impact of polymorphism driven liver-stage immune evasion by malaria parasites, to help design effective cross-reactive vaccines. Front. Microbiol. 2016, 7, 303. [Google Scholar] [CrossRef]
- Kessler, B.M.; Bassanini, P.; Cerottini, J.C.; Luescher, I.F. Effects of epitope modification on t cell receptor-ligand binding and antigen recognition by seven h-2kd-restricted cytotoxic t lymphocyte clones specific for a photoreactive peptide derivative. J. Exp. Med. 1997, 185, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Romero, P.; Corradin, G.; Luescher, I.F.; Maryanski, J.L. H-2kd-restricted antigenic peptides share a simple binding motif. J. Exp. Med. 1991, 174, 603–612. [Google Scholar] [CrossRef]
- Fifis, T.; Mottram, P.; Bogdanoska, V.; Hanley, J.; Plebanski, M. Short peptide sequences containing mhc class i and/or class ii epitopes linked to nano-beads induce strong immunity and inhibition of growth of antigen-specific tumour challenge in mice. Vaccine 2004, 23, 258–266. [Google Scholar] [CrossRef]
- Mottram, P.L.; Leong, D.; Crimeen-Irwin, B.; Gloster, S.; Xiang, S.D.; Meanger, J.; Ghildyal, R.; Vardaxis, N.; Plebanski, M. Type 1 and 2 immunity following vaccination is influenced by nanoparticle size: Formulation of a model vaccine for respiratory syncytial virus. Mol. Pharm. 2007, 4, 73–84. [Google Scholar] [CrossRef]
- Chen, W.; Mccluskey, J.; Rodda, S.; Carbone, F.R. Changes at peptide residues buried in the major histocompatibility complex (mhc) class-i binding cleft influence t-cell recognition—A possible role for indirect conformational alterations in the mhc class-i or bound peptide in determining t-cell recognition. J. Exp. Med. 1993, 177, 869–873. [Google Scholar]
- Kim, C.G.; Kye, Y.C.; Yun, C.H. The role of nanovaccine in cross-presentation of antigen-presenting cells for the activation of cd8(+) t cell responses. Pharmaceutics 2019, 11, 612. [Google Scholar] [CrossRef] [Green Version]
- Mitaksov, V.; Fremont, D.H. Structural definition of the h-2kd peptide-binding motif. J. Biol. Chem. 2006, 281, 10618–10625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karapetyan, A.R.; Chaipan, C.; Winkelbach, K.; Wimberger, S.; Jeong, J.S.; Joshi, B.; Stein, R.B.; Underwood, D.; Castle, J.C.; van Dijk, M.; et al. Tcr fingerprinting and off-target peptide identification. Front. Immunol. 2019, 10, 2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calis, J.J.; Maybeno, M.; Greenbaum, J.A.; Weiskopf, D.; De Silva, A.D.; Sette, A.; Kesmir, C.; Peters, B. Properties of mhc class i presented peptides that enhance immunogenicity. PLoS Comput. Biol. 2013, 9, e1003266. [Google Scholar] [CrossRef] [Green Version]
- Chowell, D.; Krishna, S.; Becker, P.D.; Cocita, C.; Shu, J.; Tan, X.F.; Greenberg, P.D.; Klavinskis, L.S.; Blattman, J.N.; Anderson, K.S. Tcr contact residue hydrophobicity is a hallmark of immunogenic cd8(+) t cell epitopes. Proc. Natl. Acad. Sci. USA 2015, 112, E1754–E1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designated Name | Peptide Sequence 1 | PI2 | Net Charge at pH 7 |
|---|---|---|---|
| KI | SYIPSAEKI | 6.58 | 0 |
| A8 | SYIPSAEAI | 0.94 | −1 |
| A7 | SYIPSAAKI | 9.5 | 1 |
| A5 | SYIPAAEKI | 6.58 | 0 |
| A4 | SYIASAEKI | 6.58 | 0 |
| A3 | SYAPSAEKI | 6.58 | 0 |
| A1 | AYIPSAEKI | 6.84 | 0 |
| Peptide Purity 1 | Concentration of Conjugated Peptide 2 | # of Peptide Molecules per Particle 3 | Size of PSNPs after Conjugation 4 | PdI 5 | % PSNPs Solids Injected 6 | µg Peptide Injected per Mouse | |
|---|---|---|---|---|---|---|---|
| PSNPs-KI | >95% | 0.21 mg/mL | 1047.77 | 45.29 ± 1.3 nm | 0.077 | 0.92% | 25 ug |
| PSNPs-A7 | >95% | 0.29 mg/mL | 1390.92 | 46.59 ± 1.4 nm | 0.075 | 0.72% | 25 ug |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson, K.L.; Xiang, S.D.; Plebanski, M. Functional Recognition by CD8+ T Cells of Epitopes with Amino Acid Variations Outside Known MHC Anchor or T Cell Receptor Recognition Residues. Int. J. Mol. Sci. 2020, 21, 4700. https://doi.org/10.3390/ijms21134700
Wilson KL, Xiang SD, Plebanski M. Functional Recognition by CD8+ T Cells of Epitopes with Amino Acid Variations Outside Known MHC Anchor or T Cell Receptor Recognition Residues. International Journal of Molecular Sciences. 2020; 21(13):4700. https://doi.org/10.3390/ijms21134700
Chicago/Turabian StyleWilson, Kirsty L., Sue D. Xiang, and Magdalena Plebanski. 2020. "Functional Recognition by CD8+ T Cells of Epitopes with Amino Acid Variations Outside Known MHC Anchor or T Cell Receptor Recognition Residues" International Journal of Molecular Sciences 21, no. 13: 4700. https://doi.org/10.3390/ijms21134700
APA StyleWilson, K. L., Xiang, S. D., & Plebanski, M. (2020). Functional Recognition by CD8+ T Cells of Epitopes with Amino Acid Variations Outside Known MHC Anchor or T Cell Receptor Recognition Residues. International Journal of Molecular Sciences, 21(13), 4700. https://doi.org/10.3390/ijms21134700