ALIAmides Update: Palmitoylethanolamide and Its Formulations on Management of Peripheral Neuropathic Pain




Abstract
1. Introduction
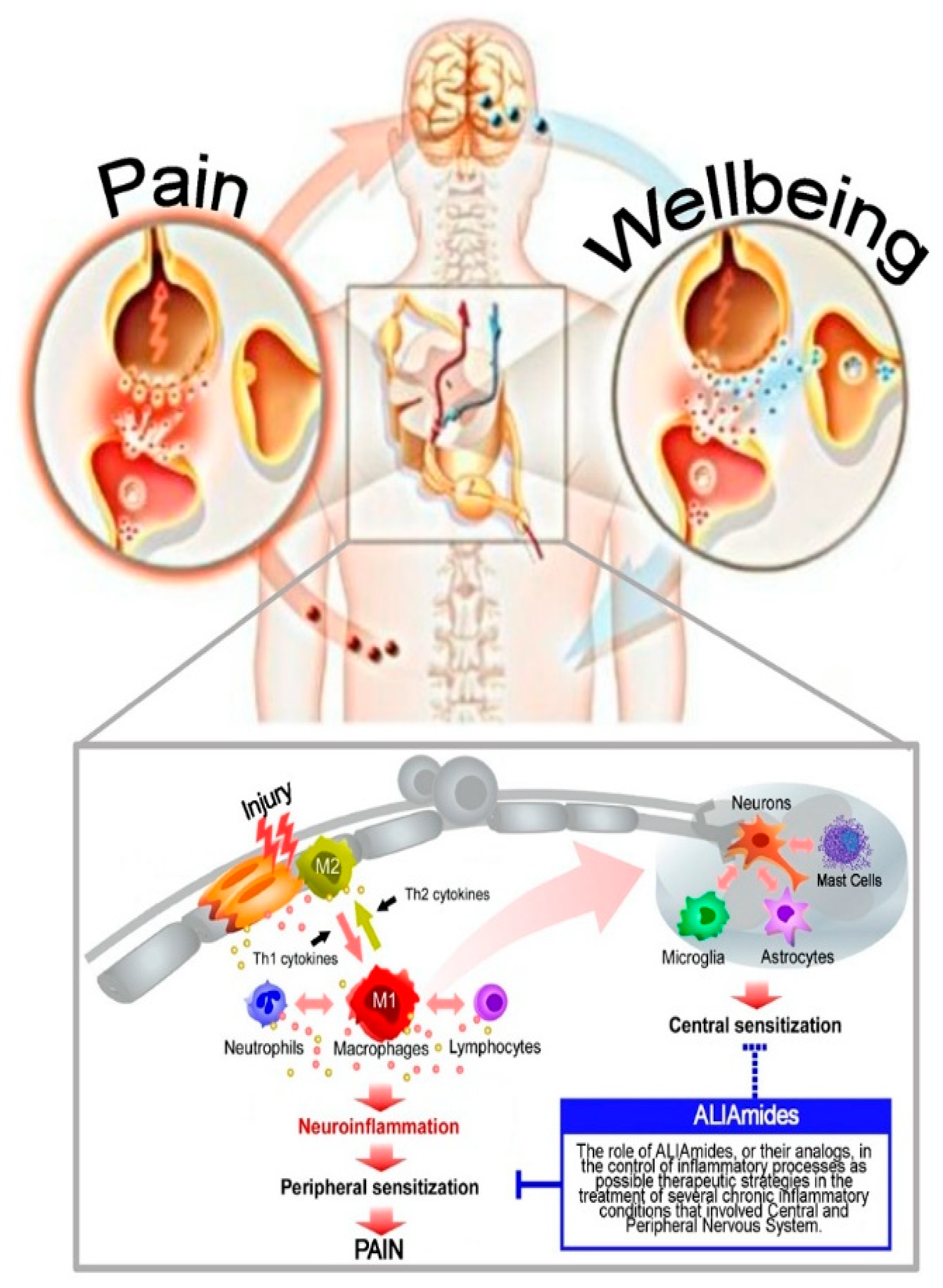
2. Neuroinflammation
3. Pharmacotherapy in Peripheral Neuropathic Pain
4. Overview on ALIAmides
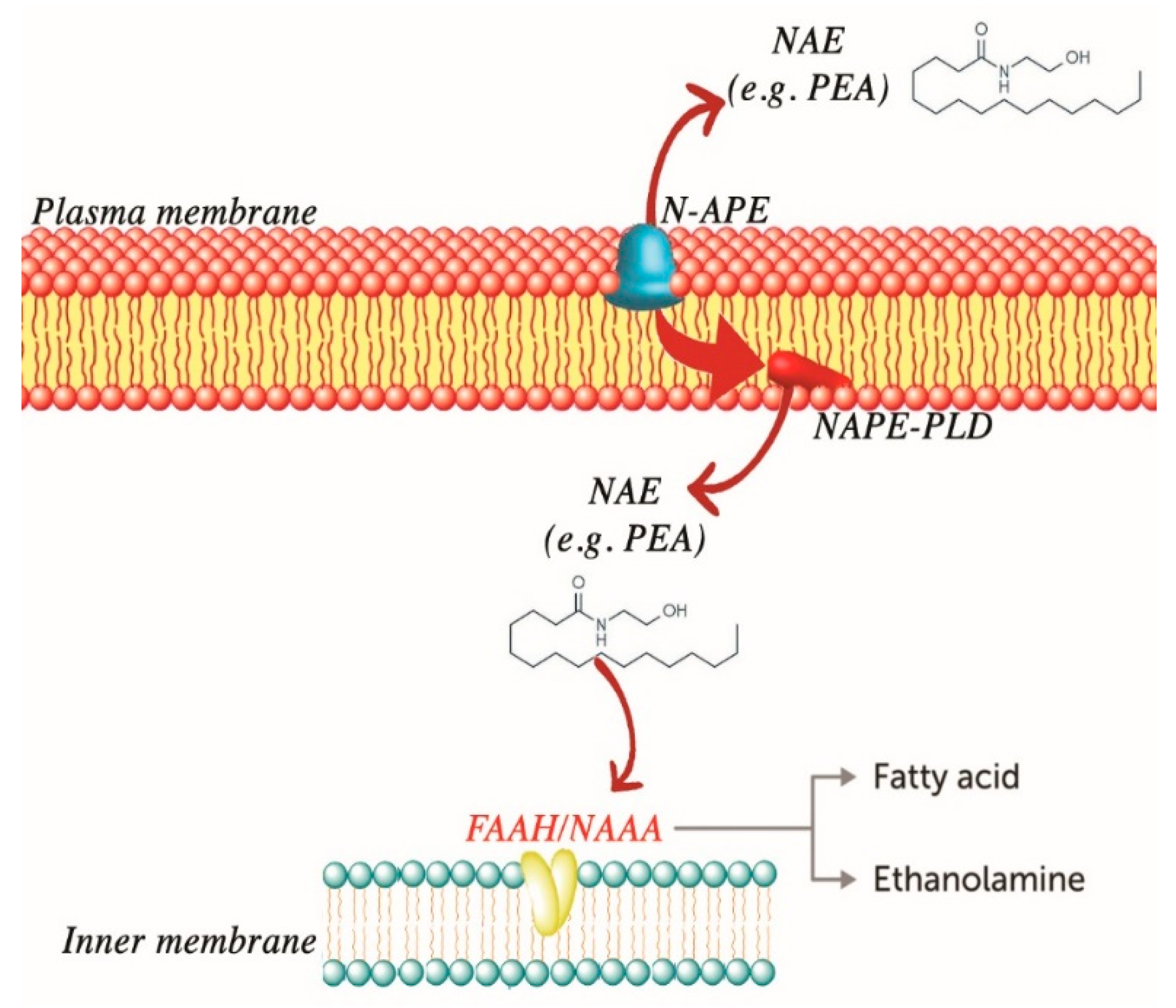
4.1. PEA
4.2. OEA and SEA
4.3. Adelmidrol
4.4. Glupamid
5. ALIAmides in Peripheral Neuropathic Pain
Clinical Studies
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALIAmides | Autacoid local injury antagonist amides |
| BBB | Blood–brain barrier |
| BSCB | Blood-spinal cord barrier |
| CCI | Chronic constriction injury |
| CIPN | Chemotherapy-induced peripheral neuropathy |
| CNS | Central nervous system |
| DPN | Diabetic painful neuropathy (DPN) |
| GPR | Receptor G-protein coupling |
| IASP | International Association for the Study of Pain |
| MAPK | Mitogen-activated protein kinase |
| MC | Mast cell |
| m-PEA | micronized-PEA |
| NAAA | N-acylethanolamine-hydrolyzing acid amidase |
| NAEs | N-acyl ethanolamines |
| OEA | Oleoyl ethanolamide |
| PEA | Palmitoyl ethanolamide |
| PEA-OXA | 2-pentadecyl-2-oxazoline |
| PGA | N-Palmitoyl-d-glucosamine |
| PNI | Peripheral nerve injury |
| PNS | Peripheral nervous system |
| PPAR-α | Peroxisome proliferator-activated receptor-α |
| ROS | Reactive oxygen species |
| SEA | Stearoyl ethanolamide |
| SNI | Spared nerve injury |
| SNRIs | Serotonin-norepinephrine reuptake inhibitors |
| STZ | Streptozotocin |
| TRPV1 | Transient receptor potential vanilloid receptor type 1 |
| TCAs | Tricyclic antidepressants |
| um-PEA | Ultramicronized-PEA |
References
- Basu, P.; Basu, A. In Vitro and In Vivo Effects of Flavonoids on Peripheral Neuropathic Pain. Molecules 2020, 25, 1171. [Google Scholar] [CrossRef] [PubMed]
- Price, T.J.; Gold, M.S. From Mechanism to Cure: Renewing the Goal to Eliminate the Disease of Pain. Pain Med. 2017, 19, 1525–1549. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.P.; Mao, J. Neuropathic pain: Mechanisms and their clinical implications. BMJ 2014, 348, f7656. [Google Scholar] [CrossRef] [PubMed]
- Szok, D.; Tajti, J.; Nyári, A.; Vecsei, L.; Trojano, L. Therapeutic Approaches for Peripheral and Central Neuropathic Pain. Behav. Neurol. 2019, 2019, 1–13. [Google Scholar] [CrossRef]
- Meacham, K.W.P.; Shepherd, A.J.; Mohapatra, D.P.; Haroutounian, S. Neuropathic Pain: Central vs. Peripheral Mechanisms. Curr. Pain Headache Rep. 2017, 21, 2204. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.C.; Sandroni, P. Central Neuropathic Pain Syndromes. Mayo Clin. Proc. 2016, 91, 372–385. [Google Scholar] [CrossRef]
- Parada, C.A.; Tambeli, C.H.; Green, P.; Cairns, B.E. Primary Afferent Nociceptor as a Target for the Relief of Pain. Pain Res. Treat. 2012, 2012, 1–2. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B.; et al. Neuropathic pain. Nat. Rev. Dis. Prim. 2017, 3, 17002. [Google Scholar] [CrossRef]
- Amir, R.; Argoff, C.E.; Bennett, G.J.; Cummins, T.R.; Durieux, M.E.; Gerner, P.; Gold, M.S.; Porreca, F.; Strichartz, G.R. The Role of Sodium Channels in Chronic Inflammatory and Neuropathic Pain. J. Pain 2006, 7, S1–S29. [Google Scholar] [CrossRef]
- Lai, J.; Hunter, J.C.; Porreca, F. The role of voltage-gated sodium channels in neuropathic pain. Curr. Opin. Neurobiol. 2003, 13, 291–297. [Google Scholar] [CrossRef]
- Reichling, D.B.; Green, P.G.; Levine, J.D. The fundamental unit of pain is the cell. Pain 2013, 154 (Suppl. 1), S2–S9. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.S.J. Advances in understanding nociception and neuropathic pain. J. Neurol. 2017, 265, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Zilliox, L.A. Neuropathic Pain. Contin. Lifelong Learn. Neurol. 2017, 23, 512–532. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, W.M.; Buglewicz, E.; Szikszay, T.M.; Luedtke, K.; Bąbel, P. Reward for Pain: Hyperalgesia and Allodynia Induced by Operant Conditioning: Systematic Review and Meta-Analysis. J. Pain 2019, 20, 861–875. [Google Scholar] [CrossRef]
- Smith, B.H.; Torrance, N.; Bennett, M.I.; Lee, A.J. Health and Quality of Life Associated With Chronic Pain of Predominantly Neuropathic Origin in the Community. Clin. J. Pain 2007, 23, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.S.; Finnerup, N. Allodynia and hyperalgesia in neuropathic pain: Clinical manifestations and mechanisms. Lancet Neurol. 2014, 13, 924–935. [Google Scholar] [CrossRef]
- Murray, G. Referred pain, allodynia and hyperalgesia. J. Am. Dent. Assoc. (1939) 2009, 140, 1122–1124. [Google Scholar] [CrossRef]
- Kalynovska, N.; Diallo, M.; Sotakova-Kasparova, D.; Palecek, J. Losartan attenuates neuroinflammation and neuropathic pain in paclitaxel-induced peripheral neuropathy. J. Cell. Mol. Med. 2020. [Google Scholar] [CrossRef]
- Penas, C.; Navarro, X. Epigenetic Modifications Associated to Neuroinflammation and Neuropathic Pain after Neural Trauma. Front. Cell. Neurosci. 2018, 12, 158. [Google Scholar] [CrossRef]
- Ellis, A.; Bennett, D.L.H. Neuroinflammation and the generation of neuropathic pain. Br. J. Anaesth. 2013, 111, 26–37. [Google Scholar] [CrossRef]
- Myers, R.R.; Campana, W.M.; Shubayev, V.I. The role of neuroinflammation in neuropathic pain: Mechanisms and therapeutic targets. Drug Discov. Today 2006, 11, 8–20. [Google Scholar] [CrossRef]
- Ji, R.-R.; Xu, Z.-Z.; Gao, Y.-J. Emerging targets in neuroinflammation-driven chronic pain. Nat. Rev. Drug Discov. 2014, 13, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.-R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhang, Y.-Q.; Qadri, Y.; Serhan, C.N.; Ji, R.-R. Microglia in Pain: Detrimental and Protective Roles in Pathogenesis and Resolution of Pain. Neuron 2018, 100, 1292–1311. [Google Scholar] [CrossRef] [PubMed]
- Breivik, H.; Gordh, T.E. Neuroinflammation explains aspects of chronic pain and opens new avenues for therapeutic interventions. Scand. J. Pain 2010, 1, 65–66. [Google Scholar] [CrossRef]
- Myers, R.R.; Shubayev, V.I. The ology of neuropathy: An integrative review of the role of neuroinflammation and TNF-α axonal transport in neuropathic pain. J. Peripher. Nerv. Syst. 2011, 16, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Amor, S.; Woodroofe, N. Innate and adaptive immune responses in neurodegeneration and repair. Immunology 2014, 141, 287–291. [Google Scholar] [CrossRef]
- Muthuraju, S.; Zakaria, R.; Karuppan, M.K.M.; Al-Rahbi, B. The Role of Neuroinflammation in Cellular Damage in Neurodegenerative Diseases. Biomed. Res. Int. 2020, 2020, 9231452. [Google Scholar] [CrossRef]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-mediated neuroinflammation in neurodegenerative diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef]
- Najjar, S.; Pearlman, D.M.; Alper, K.; Najjar, A.; Devinsky, O. Neuroinflammation and psychiatric illness. J. Neuroinflamm. 2013, 10, 43. [Google Scholar] [CrossRef]
- Matta, S.M.; Hill-Yardin, E.L.; Crack, P.J. The influence of neuroinflammation in Autism Spectrum Disorder. Brain Behav. Immun. 2019, 79, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M. Microglia-Mediated Regulation of Neuropathic Pain: Molecular and Cellular Mechanisms. Biol. Pharm. Bull. 2019, 42, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Tsuda, M. Microglia in neuropathic pain: Cellular and molecular mechanisms and therapeutic potential. Nat. Rev. Neurosci. 2018, 19, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M. Microglia in the spinal cord and neuropathic pain. J. Diabetes Investig. 2015, 7, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Aldskogius, H.; Kozlova, E.N. Microglia and neuropathic pain. CNS Neurol. Disord. Drug Targets 2013, 12, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Davalos, D.; Grutzendler, J.; Yang, G.; Kim, J.V.; Zuo, Y.; Jung, S.; Littman, D.R.; Dustin, M.L.; Gan, W.-B. ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 2005, 8, 752–758. [Google Scholar] [CrossRef]
- Mika, J.; Osikowicz, M.; Rojewska, E.; Korostynski, M.; Wawrzczak-Bargieła, A.; Przewlocki, R.; Przewlocka, B. Differential activation of spinal microglial and astroglial cells in a mouse model of peripheral neuropathic pain. Eur. J. Pharmacol. 2009, 623, 65–72. [Google Scholar] [CrossRef]
- Carniglia, L.; Ramírez, D.; Durand, D.; Saba, J.; Turati, J.; Caruso, C.; Scimonelli, T.N.; Lasaga, M. Neuropeptides and Microglial Activation in Inflammation, Pain, and Neurodegenerative Diseases. Mediat. Inflamm. 2017, 2017, 1–23. [Google Scholar] [CrossRef]
- Wen, Y.R.; Tan, P.-H.; Cheng, J.-K.; Liu, Y.-C.; Ji, R.-R. Microglia: A Promising Target for Treating Neuropathic and Postoperative Pain, and Morphine Tolerance. J. Formos. Med. Assoc. 2011, 110, 487–494. [Google Scholar] [CrossRef]
- Suter, M.R.; Wen, Y.R.; Decosterd, I.; Ji, R.-R. Do glial cells control pain? Neuron Glia Biol. 2007, 3, 255–268. [Google Scholar] [CrossRef]
- Echeverry, S.; Shi, X.Q.; Yang, M.; Huang, H.; Wu, Y.; Lorenzo, L.-E.; Perez-Sanchez, J.; Bonin, R.; De Koninck, Y.; Zhang, J. Spinal microglia are required for long-term maintenance of neuropathic pain. Pain 2017, 158, 1792–1801. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.K.; Malcangio, M. Fractalkine/CX3CR1 signaling during neuropathic pain. Front. Cell. Neurosci. 2014, 8, 121. [Google Scholar] [CrossRef]
- Clark, A.; Staniland, A.A.; Malcangio, M. Fractalkine/CX3CR1 signalling in chronic pain and inflammation. Curr. Pharm. Biotechnol. 2011, 12, 1707–1714. [Google Scholar] [CrossRef]
- Inoue, K. Role of the P2X4 receptor in neuropathic pain. Curr. Opin. Pharmacol. 2019, 47, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Zhu, Z.M.; Liu, Z.X. The role of P2X4 receptor in neuropathic pain and its pharmacological properties. Pharmacol. Res. 2020, 158, 104875. [Google Scholar] [CrossRef] [PubMed]
- Jurga, A.M.; Piotrowska, A.; Makuch, W.; Przewlocka, B.; Mika, J. Blockade of P2X4 Receptors Inhibits Neuropathic Pain-Related Behavior by Preventing MMP-9 Activation and, Consequently, Pronociceptive Interleukin Release in a Rat Model. Front. Pharmacol. 2017, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.-J.; Jia, L.; Zhang, X.; Wei, H.; Yue, S.-W. MAPK Pathways Are Involved in Neuropathic Pain in Rats with Chronic Compression of the Dorsal Root Ganglion. Evid.-Based Complement. Altern. Med. 2016, 2016, 1–8. [Google Scholar] [CrossRef]
- Obata, K.; Noguchi, K. MAPK activation in nociceptive neurons and pain hypersensitivity. Life Sci. 2004, 74, 2643–2653. [Google Scholar] [CrossRef]
- Lin, X.; Wang, M.; Zhang, J.; Xu, R. p38 MAPK: A potential target of chronic pain. Curr. Med. Chem. 2014, 21, 4405–4418. [Google Scholar] [CrossRef]
- Wu, J.; Xu, Y.; Pu, S.; Jiang, W.; Du, D. p38/MAPK inhibitor modulates the expression of dorsal horn GABA(B) receptors in the spinal nerve ligation model of neuropathic pain. Neuroimmunomodulation 2011, 18, 150–155. [Google Scholar] [CrossRef]
- Suzuki, N.; Hasegawa-Moriyama, M.; Takahashi, Y.; Kamikubo, Y.; Sakurai, T.; Inada, E. Lidocaine Attenuates the Development of Diabetic-Induced Tactile Allodynia by Inhibiting Microglial Activation. Anesthesia Analg. 2011, 113, 941–946. [Google Scholar] [CrossRef]
- Choi, Y.-S.; Jun, I.-G.; Kim, S.-H.; Park, J.Y. Intrathecal Lamotrigine Attenuates Mechanical Allodynia and Suppresses Microglial and Astrocytic Activation in a Rat Model of Spinal Nerve Ligation. Yonsei Med. J. 2013, 54, 321–329. [Google Scholar] [CrossRef][Green Version]
- Han, S.R.; Yang, G.Y.; Ahn, M.H.; Kim, M.J.; Ju, J.S.; Bae, Y.C.; Ahn, D.-K. Blockade of microglial activation reduces mechanical allodynia in rats with compression of the trigeminal ganglion. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 36, 52–59. [Google Scholar] [CrossRef]
- Skaper, S.D. Mast Cell—Glia Dialogue in Chronic Pain and Neuropathic Pain: Blood-Brain Barrier Implications. CNS Neurol. Disord. Drug Targets 2016, 15, 1072–1078. [Google Scholar] [CrossRef]
- Cordaro, M.; Cuzzocrea, S.; Crupi, R. An Update of Palmitoylethanolamide and Luteolin Effects in Preclinical and Clinical Studies of Neuroinflammatory Events. Antioxidants 2020, 9, 216. [Google Scholar] [CrossRef]
- Green, D.P.; Limjunyawong, N.; Gour, N.; Pundir, P.; Dong, X. A Mast-Cell-Specific Receptor Mediates Neurogenic Inflammation and Pain. Neuron 2019, 101, 412–420. [Google Scholar] [CrossRef]
- Ji, R.-R.; Chamessian, A.; Zhang, Y.-Q. Pain regulation by non-neuronal cells and inflammation. Science 2016, 354, 572–577. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L.; Giusti, P. Mast cells, glia and neuroinflammation: Partners in crime? Immunology 2014, 141, 314–327. [Google Scholar] [CrossRef]
- Kles, K.; Vinik, A. Pathophysiology and Treatment of Diabetic Peripheral Neuropathy: The Case for Diabetic Neurovascular Function as an Essential Component. Curr. Diabetes Rev. 2006, 2, 131–145. [Google Scholar] [CrossRef]
- Sommer, C.; Leinders, M.; Üçeyler, N. Inflammation in the pathophysiology of neuropathic pain. Pain 2018, 159, 595–602. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Nedergaard, M. The homeostatic astroglia emerges from evolutionary specialization of neural cells. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150428. [Google Scholar] [CrossRef]
- Tsai, H.-H.; Li, H.; Fuentealba, L.C.; Molofsky, A.V.; Taveira-Marques, R.; Zhuang, H.; Tenney, A.; Murnen, A.T.; Fancy, S.P.J.; Merkle, F.; et al. Regional Astrocyte Allocation Regulates CNS Synaptogenesis and Repair. Science 2012, 337, 358–362. [Google Scholar] [CrossRef]
- Chung, W.-S.; Allen, N.J.; Eroglu, C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020370. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Taveggia, C.; Melendez-Vasquez, C.V.; Einheber, S.; Raine, C.S.; Salzer, J.L.; Brosnan, C.F.; John, G.R. Interleukin-11 Potentiates Oligodendrocyte Survival and Maturation, and Myelin Formation. J. Neurosci. 2006, 26, 12174–12185. [Google Scholar] [CrossRef] [PubMed]
- Singhal, G.; Jaehne, E.J.; Corrigan, F.; Toben, C.; Baune, B.T. Inflammasomes in neuroinflammation and changes in brain function: A focused review. Front. Mol. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Voet, S.; Srinivasan, S.; Lamkanfi, M.; Van Loo, G. Inflammasomes in neuroinflammatory and neurodegenerative diseases. EMBO Mol. Med. 2019, 11, e10248. [Google Scholar] [CrossRef]
- Ramesh, G.; MacLean, A.G.; Philipp, M.T. Cytokines and Chemokines at the Crossroads of Neuroinflammation, Neurodegeneration, and Neuropathic Pain. Mediat. Inflamm. 2013, 2013, 1–20. [Google Scholar] [CrossRef]
- Becher, B.; Spath, S.; Goverman, J. Cytokine networks in neuroinflammation. Nat. Rev. Immunol. 2016, 17, 49–59. [Google Scholar] [CrossRef]
- Mills, S.; Nicolson, K.P.; Smith, B.H. Chronic pain: A review of its epidemiology and associated factors in population-based studies. Br. J. Anaesth. 2019, 123, e273–e283. [Google Scholar] [CrossRef]
- Reid, K.; Harker, J.; Bala, M.M.; Truyers, C.; Kellen, E.; E Bekkering, G.; Kleijnen, J. Epidemiology of chronic non-cancer pain in Europe: Narrative review of prevalence, pain treatments and pain impact. Curr. Med. Res. Opin. 2011, 27, 449–462. [Google Scholar] [CrossRef]
- Baron, R.; Binder, A.; Wasner, G. Neuropathic pain: Diagnosis, pathophysiological mechanisms, and treatment. Lancet Neurol. 2010, 9, 807–819. [Google Scholar] [CrossRef]
- Shaygan, M.; Böger, A.; Kröner-Herwig, B. Neuropathic sensory symptoms: Association with pain and psychological factors. Neuropsychiatr. Dis. Treat. 2014, 10, 897–906. [Google Scholar] [CrossRef] [PubMed][Green Version]
- A Shipton, E. The pain experience and sociocultural factors. N. Z. Med. J. 2013, 126, 7–9. [Google Scholar] [PubMed]
- Haythornthwaite, J.A.; Benrud-Larson, L.M. Psychological aspects of neuropathic pain. Clin. J. Pain 2000, 16, S101–S105. [Google Scholar] [CrossRef] [PubMed]
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpää, M.; Hansson, P.; Jensen, T.S.; et al. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol. 2015, 14, 162–173. [Google Scholar] [CrossRef]
- Fornasari, D. Pharmacotherapy for Neuropathic Pain: A Review. Pain Ther. 2017, 6, 25–33. [Google Scholar] [CrossRef]
- Kremer, M.; Salvat, E.; Muller, A.; Yalcin, I.; Barrot, M. Antidepressants and gabapentinoids in neuropathic pain: Mechanistic insights. Neuroscience 2016, 338, 183–206. [Google Scholar] [CrossRef]
- Ruiz-Negron, N.; Menon, J.; King, J.B.; Ma, J.; Bellows, B.K. Cost-Effectiveness of Treatment Options for Neuropathic Pain: A Systematic Review. Pharmacoeconomics 2019, 37, 669–688. [Google Scholar] [CrossRef]
- Stahl, S.M.; Porreca, F.; Taylor, C.P.; Cheung, R.; Thorpe, A.J.; Clair, A. The diverse therapeutic actions of pregabalin: Is a single mechanism responsible for several pharmacological activities? Trends Pharmacol. Sci. 2013, 34, 332–339. [Google Scholar] [CrossRef]
- Bockbrader, H.N.; Wesche, D.; Miller, R.; Chapel, S.; Janiczek, N.; Burger, P.; Bockbrader, H.N. A Comparison of the Pharmacokinetics and Pharmacodynamics of Pregabalin and Gabapentin. Clin. Pharmacokinet. 2010, 49, 661–669. [Google Scholar] [CrossRef]
- Baftiu, A.; Landmark, C.J.; Rusten, I.R.; Feet, S.A.; Johannessen, S.I.; Larsson, P.G. Changes in utilisation of antiepileptic drugs in epilepsy and non-epilepsy disorders—A pharmacoepidemiological study and clinical implications. Eur. J. Clin. Pharmacol. 2016, 72, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Landmark, C.J. Antiepileptic Drugs in Non-Epilepsy Disorders. CNS Drugs 2008, 22, 27–47. [Google Scholar] [CrossRef] [PubMed]
- Dworkin, R.H.; O’Connor, A.; Audette, J.; Baron, R.; Gourlay, G.K.; Haanpää, M.L.; Kent, J.L.; Krane, E.J.; Lebel, A.A.; Levy, R.M.; et al. Recommendations for the Pharmacological Management of Neuropathic Pain: An Overview and Literature Update. Mayo Clin. Proc. 2010, 85, S3–S14. [Google Scholar] [CrossRef]
- Baron, R.; Allegri, M.; Correa-Illanes, G.; Hans, G.; Serpell, M.; Mick, G.; Mayoral, V. The 5% Lidocaine-Medicated Plaster: Its Inclusion in International Treatment Guidelines for Treating Localized Neuropathic Pain, and Clinical Evidence Supporting its Use. Pain Ther. 2016, 5, 149–169. [Google Scholar] [CrossRef] [PubMed]
- Crawford, P.; Xu, Y. Topical Capsaicin for Treatment of Chronic Neuropathic Pain in Adults. Am. Fam. Physician 2017, 96. [Google Scholar] [CrossRef]
- Schumacher, M.; Pasvankas, G. Topical capsaicin formulations in the management of neuropathic pain. Prog. Drug Res. 2014, 68, 105–128. [Google Scholar] [CrossRef]
- Deng, Y.; Luo, L.; Hu, Y.; Fang, K.; Liu, J. Clinical practice guidelines for the management of neuropathic pain: A systematic review. BMC Anesthesiol. 2016, 16, 12. [Google Scholar] [CrossRef]
- Fornasari, D. Pain pharmacology: Focus on opioids. Clin. Cases Miner. Bone Metab. 2014, 11, 165–168. [Google Scholar] [CrossRef]
- Leonardi, C.; Vellucci, R.; Mammucari, M.; Fanelli, G. Opioid risk addiction in the management of chronic pain in primary care: The addition risk questionnaire. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4898–4905. [Google Scholar]
- Mensah-Nyagan, A.G.; Meyer, L.; Schaeffer, V.; Kibaly, C.; Patte-Mensah, C. Evidence for a key role of steroids in the modulation of pain. Psychoneuroendocrinology 2009, 34, S169–S177. [Google Scholar] [CrossRef]
- Vyvey, M. Steroids as pain relief adjuvants. Can. Fam. Physician Med. Fam. Can. 2010, 56, 1295–1297. [Google Scholar]
- Otis, J.A.D.; Macone, A. Neuropathic Pain. Semin. Neurol. 2018, 38, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Moisset, X.; De Andrade, D.C.; Bouhassira, D. From pulses to pain relief: An update on the mechanisms of rTMS-induced analgesic effects. Eur. J. Pain 2015, 20, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Penson, P.E. Autonomic & Autacoid Pharmacology 2016: The year in review. Auton. Autacoid Pharmacol. 2016, 36, 27. [Google Scholar] [CrossRef] [PubMed]
- Hesselink, J.M.K. Fundamentals of and Critical Issues in Lipid Autacoid Medicine: A Review. Pain Ther. 2017, 6, 153–164. [Google Scholar] [CrossRef]
- Penson, P.E. Autonomic & Autacoid Pharmacology: Past, present and future. Auton. Autacoid Pharmacol. 2015, 35, 45. [Google Scholar] [CrossRef]
- Aloe, L.; Leon, A.; Levi-Montalcini, R. A proposed autacoid mechanism controlling mastocyte behaviour. Inflamm. Res. 1993, 39, C145–C147. [Google Scholar] [CrossRef]
- Hansen, H.S.; Diep, T.A. N-acylethanolamines, anandamide and food intake. Biochem. Pharmacol. 2009, 78, 553–560. [Google Scholar] [CrossRef]
- De Filippis, D.; Negro, L.; Vaia, M.; Cinelli, M.; Iuvone, T. New insights in mast cell modulation by palmitoylethanolamide. CNS Neurol. Disord. Drug Targets 2013, 12, 78–83. [Google Scholar] [CrossRef]
- Cantarella, G.; Scollo, M.; Lempereur, L.; Saccani-Jotti, G.; Basile, F.; Bernardini, R. Endocannabinoids inhibit release of nerve growth factor by inflammation-activated mast cells. Biochem. Pharmacol. 2011, 82, 380–388. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L.; Barbierato, M.; Zusso, M.; Bruschetta, G.; Impellizzeri, D.; Cuzzocrea, S.; Giusti, P. N-Palmitoylethanolamine and Neuroinflammation: A Novel Therapeutic Strategy of Resolution. Mol. Neurobiol. 2015, 52, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Cuzzocrea, S. Palmitoylethanolamide is a new possible pharmacological treatment for the inflammation associated with trauma. Mini Rev. Med. Chem. 2013, 13, 237–255. [Google Scholar] [PubMed]
- Wellner, N.; Diep, T.A.; Janfelt, C.; Hansen, H.S. N-acylation of phosphatidylethanolamine and its biological functions in mammals. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2013, 1831, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiù, V.; Leuti, A.; Smoum, R.; Mechoulam, R.; Maccarrone, M. Bioactive lipids ALIAmides differentially modulate inflammatory responses of distinct subsets of primary human T lymphocytes. FASEB J. 2018, 32, 5716–5723. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Di Paola, R.; Cordaro, M.; Gugliandolo, E.; Casili, G.; Morittu, V.; Britti, D.; Esposito, E.; Cuzzocrea, S. Adelmidrol, a palmitoylethanolamide analogue, as a new pharmacological treatment for the management of acute and chronic inflammation. Biochem. Pharmacol. 2016, 119, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Crupi, R.; Siracusa, R.; Esposito, E.; Cuzzocrea, S. Micronized/ultramicronized palmitoylethanolamide displays superior oral efficacy compared to nonmicronized palmitoylethanolamide in a rat model of inflammatory pain. J. Neuroinflamm. 2014, 11, 1–9. [Google Scholar] [CrossRef]
- Alhouayek, M.; Muccioli, G. Harnessing the anti-inflammatory potential of palmitoylethanolamide. Drug Discov. Today 2014, 19, 1632–1639. [Google Scholar] [CrossRef]
- Ganley, O.H.; E Graessle, O.; Robinson, H.J. Anti-inflammatory activity on compounds obtained from egg yolk, peanut oil, and soybean lecithin. J. Lab. Clin. Med. 1958, 51, 709–714. [Google Scholar]
- Beggiato, S.; Tomasini, M.C.; Ferraro, L. Palmitoylethanolamide (PEA) as a Potential Therapeutic Agent in Alzheimer’s Disease. Front. Pharmacol. 2019, 10, 821. [Google Scholar] [CrossRef]
- Levi-Montalcini, R.; Skaper, S.D.; Toso, R.D.; Petrelli, L.; Leon, A. Nerve growth factor: From neurotrophin to neurokine. Trends Neurosci. 1996, 19, 514–520. [Google Scholar] [CrossRef]
- Peritore, A.F.; Siracusa, R.; Crupi, R.; Cuzzocrea, S. Therapeutic Efficacy of Palmitoylethanolamide and Its New Formulations in Synergy with Different Antioxidant Molecules Present in Diets. Nutrients 2019, 11, 2175. [Google Scholar] [CrossRef]
- Schuel, H.; Burkman, L.J.; Lippes, J.; Crickard, K.; Forester, E.; Piomelli, D.; Giuffrida, A. N-Acylethanolamines in human reproductive fluids. Chem. Phys. Lipids 2002, 121, 211–227. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Peritore, A.F.; Piras, C.; Cuzzocrea, S.; Crupi, R. Palmitoylethanolamide and Related ALIAmides: Prohomeostatic Lipid Compounds for Animal Health and Wellbeing. Veter. Sci. 2020, 7, 78. [Google Scholar] [CrossRef]
- Gabrielsson, L.; Mattsson, S.; Fowler, C.J. Palmitoylethanolamide for the treatment of pain: Pharmacokinetics, safety and efficacy. Br. J. Clin. Pharmacol. 2016, 82, 932–942. [Google Scholar] [CrossRef]
- Nestmann, E.R. Safety of micronized palmitoylethanolamide (microPEA): Lack of toxicity and genotoxic potential. Food Sci. Nutr. 2016, 5, 292–309. [Google Scholar] [CrossRef]
- Petrosino, S.; Di Marzo, V. The pharmacology of palmitoylethanolamide and first data on the therapeutic efficacy of some of its new formulations. Br. J. Pharmacol. 2016, 174, 1349–1365. [Google Scholar] [CrossRef]
- Tsuboi, K.; Uyama, T.; Okamoto, Y.; Ueda, N. Endocannabinoids and related N-acylethanolamines: Biological activities and metabolism. Inflamm. Regen. 2018, 38, 28. [Google Scholar] [CrossRef]
- Petrosino, S.; Iuvone, T.; Di Marzo, V. N-palmitoyl-ethanolamine: Biochemistry and new therapeutic opportunities. Biochimie 2010, 92, 724–727. [Google Scholar] [CrossRef]
- Okamoto, Y.; Morishita, J.; Tsuboi, K.; Tonai, T.; Ueda, N. Molecular Characterization of a Phospholipase D Generating Anandamide and Its Congeners. J. Biol. Chem. 2003, 279, 5298–5305. [Google Scholar] [CrossRef]
- Ueda, N.; Tsuboi, K.; Uyama, T. Enzymological studies on the biosynthesis of N-acylethanolamines. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2010, 1801, 1274–1285. [Google Scholar] [CrossRef]
- Verme, J.L.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The Nuclear Receptor Peroxisome Proliferator-Activated Receptor-α Mediates the Anti-Inflammatory Actions of Palmitoylethanolamide. Mol. Pharmacol. 2004, 67, 15–19. [Google Scholar] [CrossRef]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.-O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol. 2009, 152, 1092–1101. [Google Scholar] [CrossRef]
- De Novellis, V.; Luongo, L.; Guida, F.; Cristino, L.; Palazzo, E.; Russo, R.; Marabese, I.; D’Agostino, G.; Calignano, A.; Rossi, F.; et al. Effects of intra-ventrolateral periaqueductal grey palmitoylethanolamide on thermoceptive threshold and rostral ventromedial medulla cell activity. Eur. J. Pharmacol. 2012, 676, 41–50. [Google Scholar] [CrossRef]
- Ho, W.-S.V.; A Barrett, D.; Randall, M.D. ‘Entourage’ effects ofN-palmitoylethanolamide andN-oleoylethanolamide on vasorelaxation to anandamide occur through TRPV1 receptors. Br. J. Pharmacol. 2008, 155, 837–846. [Google Scholar] [CrossRef]
- Di Marzo, V.; Melck, M.; Orlando, P.; Bisogno, T.; Zagoory, O.; Bifulco, M.; Vogel, Z.; De Petrocellis, L. Palmitoylethanolamide inhibits the expression of fatty acid amide hydrolase and enhances the anti-proliferative effect of anandamide in human breast cancer cells. Biochem. J. 2001, 358, 249–255. [Google Scholar] [CrossRef]
- Ueda, N.; Yamanaka, K.; Yamamoto, S. Purification and Characterization of an Acid Amidase Selective forN-Palmitoylethanolamine, a Putative Endogenous Anti-inflammatory Substance. J. Biol. Chem. 2001, 276, 35552–35557. [Google Scholar] [CrossRef]
- Rasenack, N.; Müller, B.W. Micron-Size Drug Particles: Common and Novel Micronization Techniques. Pharm. Dev. Technol. 2004, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Cordaro, M.; Crupi, R.; Siracusa, R.; Campolo, M.; Bruschetta, G.; Fusco, R.; Pugliatti, P.; Esposito, E.; Cuzzocrea, S. Protective Effects of Ultramicronized Palmitoylethanolamide (PEA-um) in Myocardial Ischaemia and Reperfusion Injury in VIVO. Shock 2016, 46, 202–213. [Google Scholar] [CrossRef]
- Di Paola, R.; Impellizzeri, D.; Fusco, R.; Cordaro, M.; Siracusa, R.; Crupi, R.; Esposito, E.; Cuzzocrea, S. Ultramicronized palmitoylethanolamide (PEA-um ®) in the treatment of idiopathic pulmonary fibrosis. Pharmacol. Res. 2016, 111, 405–412. [Google Scholar] [CrossRef]
- Crupi, R.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Casili, G.; Evangelista, M.; Cuzzocrea, S. N-palmitoylethanolamide Prevents Parkinsonian Phenotypes in Aged Mice. Mol. Neurobiol. 2018, 55, 8455–8472. [Google Scholar] [CrossRef]
- Di Paola, R.; Impellizzeri, D.; Torre, A.; Mazzon, E.; Cappellani, A.; Faggio, C.; Esposito, E.; Trischitta, F.; Cuzzocrea, S. Effects of palmitoylethanolamide on intestinal injury and inflammation caused by ischemia-reperfusion in mice. J. Leukoc. Biol. 2012, 91, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Ahmad, A.; Bruschetta, G.; Di Paola, R.; Crupi, R.; Paterniti, I.; Esposito, E.; Cuzzocrea, S. The anti-inflammatory effects of palmitoylethanolamide (PEA) on endotoxin-induced uveitis in rats. Eur. J. Pharmacol. 2015, 761, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Campolo, M.; Impellizzeri, D.; Paterniti, I.; Allarà, M.; Gugliandolo, E.; D’Amico, R.; Siracusa, R.; Cordaro, M.; Esposito, E.; et al. 2-Pentadecyl-2-Oxazoline, the Oxazoline of Pea, Modulates Carrageenan-Induced Acute Inflammation. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Scuto, M.; Siracusa, R.; D’Amico, R.; Peritore, A.F.; Gugliandolo, E.; Fusco, R.; Crupi, R.; Impellizzeri, D.; Pozzebon, M.; et al. Effect of N-palmitoylethanolamine-oxazoline on comorbid neuropsychiatric disturbance associated with inflammatory bowel disease. FASEB J. 2020, 34, 4085–4106. [Google Scholar] [CrossRef]
- Fusco, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Gugliandolo, E.; Siracusa, R.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. N-Palmitoylethanolamide-Oxazoline Protects against Middle Cerebral Artery Occlusion Injury in Diabetic Rats by Regulating the SIRT1 Pathway. Int. J. Mol. Sci. 2019, 20, 4845. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Crupi, R.; Impellizzeri, D.; Peritore, A.F.; D’Amico, R.; Gugliandolo, E.; Di Paola, R.; Cuzzocrea, S. 2-Pentadecyl-2-Oxazoline Reduces Neuroinflammatory Environment in the MPTP Model of Parkinson Disease. Mol. Neurobiol. 2018, 55, 9251–9266. [Google Scholar] [CrossRef]
- Lunardelli, M.L.; Crupi, R.; Siracusa, R.; Cocuzza, G.; Cordaro, M.; Martini, E.; Impellizzeri, D.; Di Paola, R.; Cuzzocrea, S. Co-ultraPEALut: Role in Preclinical and Clinical Delirium Manifestations. CNS Neurol. Disord. Drug Targets 2019, 18, 530–554. [Google Scholar] [CrossRef]
- Siracusa, R.; Impellizzeri, D.; Cordaro, M.; Crupi, R.; Esposito, E.; Petrosino, S.; Cuzzocrea, S. Anti-Inflammatory and Neuroprotective Effects of Co-UltraPEALut in a Mouse Model of Vascular Dementia. Front. Neurol. 2017, 8, 233. [Google Scholar] [CrossRef]
- Cordaro, M.; Impellizzeri, D.; Siracusa, R.; Gugliandolo, E.; Fusco, R.; Inferrera, A.; Esposito, E.; Di Paola, R.; Cuzzocrea, S. Effects of a co-micronized composite containing palmitoylethanolamide and polydatin in an experimental model of benign prostatic hyperplasia. Toxicol. Appl. Pharmacol. 2017, 329, 231–240. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Bruschetta, G.; Ahmad, A.; Crupi, R.; Siracusa, R.; Di Paola, R.; Paterniti, I.; Prosdocimi, M.; Esposito, E.; Cuzzocrea, S. Effects of palmitoylethanolamide and silymarin combination treatment in an animal model of kidney ischemia and reperfusion. Eur. J. Pharmacol. 2015, 762, 136–149. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Fusco, R.; Biundo, F.; D’Amico, R.; Benedetto, F.; Di Paola, R.; Cuzzocrea, S. Palmitoylethanolamide and Polydatin combination reduces inflammation and oxidative stress in vascular injury. Pharmacol. Res. 2017, 123, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Fusco, R.; Gugliandolo, E.; Crupi, R.; Evangelista, M.; Granese, R.; Cuzzocrea, S. Co-micronized Palmitoylethanolamide/Polydatin Treatment Causes Endometriotic Lesion Regression in a Rodent Model of Surgically Induced Endometriosis. Front. Pharmacol. 2016, 7, 382. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, R.; Fusco, R.; Gugliandolo, E.; Cordaro, M.; Siracusa, R.; Impellizzeri, D.; Peritore, A.F.; Crupi, R.; Cuzzocrea, S.; Di Paola, R. Effects of a new compound containing Palmitoylethanolamide and Baicalein in myocardial ischaemia/reperfusion injury in vivo. Phytomedicine 2018, 54, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Britti, D.; Crupi, R.; Impellizzeri, D.; Gugliandolo, E.; Fusco, R.; Schievano, C.; Morittu, V.; Evangelista, M.; Di Paola, R.; Cuzzocrea, S. A novel composite formulation of palmitoylethanolamide and quercetin decreases inflammation and relieves pain in inflammatory and osteoarthritic pain models. BMC Veter. Res. 2017, 13, 1–13. [Google Scholar] [CrossRef]
- Sayd, A.; Anton, M.; Alén, F.; Caso, J.R.; Pavon, J.; Leza, J.C.; De Fonseca, F.R.; Garcia-Bueno, B.; Orio, L. Systemic Administration of Oleoylethanolamide Protects from Neuroinflammation and Anhedonia Induced by LPS in Rats. Int. J. Neuropsychopharmacol. 2015, 18, 18. [Google Scholar] [CrossRef]
- Orio, L.; Alen, F.; Pavón, F.J.; Serrano, A.; García-Bueno, B. Oleoylethanolamide, Neuroinflammation, and Alcohol Abuse. Front. Mol. Neurosci. 2019, 11, 490. [Google Scholar] [CrossRef]
- De Fonseca, F.R.; Navarro, M.; Gomez, R.; Escuredo, L.; Nava, F.; Fu, J.; Murillo-Rodríguez, E.; Giuffrida, A.; LoVerme, J.; Gaetani, S.; et al. An anorexic lipid mediator regulated by feeding. Nature 2001, 414, 209–212. [Google Scholar] [CrossRef]
- Berdyshev, A.; Kosyakova, H.; Onopchenko, O.V.; Panchuk, R.; Stoika, R.S.; Hula, N.M. N-Stearoylethanolamine suppresses the pro-inflammatory cytokines production by inhibition of NF-κB translocation. Prostaglandins Other Lipid Mediat. 2015, 121, 91–96. [Google Scholar] [CrossRef]
- Brown, J.; Azari, E.K.; Ayala, J.E. Oleoylethanolamide: A fat ally in the fight against obesity. Physiol. Behav. 2017, 176, 50–58. [Google Scholar] [CrossRef]
- Onopchenko, O.V.; Kosiakova, G.V.; Oz, M.; Klimashevsky, V.M.; Gula, N.M. N-Stearoylethanolamine Restores Pancreas Lipid Composition in Obesity-Induced Insulin Resistant Rats. Lipids 2014, 50, 13–21. [Google Scholar] [CrossRef]
- Malan, T.P.; Porreca, F.; Malan, T.P. Lipid mediators regulating pain sensitivity. Prostaglandins Other Lipid Mediat. 2005, 77, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Suardíaz, M.; Torrús, G.E.; Goicoechea, C.; Bilbao, A.; De Fonseca, F.R. Analgesic properties of oleoylethanolamide (OEA) in visceral and inflammatory pain. Pain 2007, 133, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Rovito, D.; Giordano, C.; Plastina, P.; Barone, I.; De Amicis, F.; Mauro, L.; Rizza, P.; Lanzino, M.; Catalano, S.; Bonofiglio, D.; et al. Omega-3 DHA- and EPA–dopamine conjugates induce PPARγ-dependent breast cancer cell death through autophagy and apoptosis. Biochim. Biophys. Acta (BBA) Gen. Subj. 2015, 1850, 2185–2195. [Google Scholar] [CrossRef]
- De Filippis, D.; D’Amico, A.; Cinelli, M.P.; Esposito, G.; Di Marzo, V.; Iuvone, T. Adelmidrol, a palmitoylethanolamide analogue, reduces chronic inflammation in a carrageenin-granuloma model in rats. J. Cell. Mol. Med. 2009, 13, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Pulvirenti, N.; Nasca, M.R.; Micali, G. Topical adelmidrol 2% emulsion, a novel aliamide, in the treatment of mild atopic dermatitis in pediatric subjects: A pilot study. Acta Dermatovenerol. Croat. 2007, 15, 80–83. [Google Scholar] [PubMed]
- Cerrato, S.; Brazis, P.; Della Valle, M.F.; Miolo, A.; Puigdemont, A. Inhibitory effect of topical Adelmidrol on antigen-induced skin wheal and mast cell behavior in a canine model of allergic dermatitis. BMC Veter. Res. 2012, 8, 230. [Google Scholar] [CrossRef]
- Russo, A.; Russo, G.; Peticca, M.; Pietropaolo, C.; Di Rosa, M.; Iuvone, T. Inhibition of granuloma-associated angiogenesis by controlling mast cell mediator release: Role of mast cell protease-5. Br. J. Pharmacol. 2005, 145, 24–33. [Google Scholar] [CrossRef]
- Borrelli, F.; Romano, B.; Petrosino, S.; Pagano, E.; Capasso, R.; Coppola, D.; Battista, G.; Orlando, P.; Di Marzo, V.; Izzo, A.A. Palmitoylethanolamide, a naturally occurring lipid, is an orally effective intestinal anti-inflammatory agent. Br. J. Pharmacol. 2014, 172, 142–158. [Google Scholar] [CrossRef]
- Cordaro, M.; Impellizzeri, D.; Gugliandolo, E.; Siracusa, R.; Crupi, R.; Esposito, E.; Cuzzocrea, S. Adelmidrol, a Palmitoylethanolamide Analogue, as a New Pharmacological Treatment for the Management of Inflammatory Bowel Disease. Mol. Pharmacol. 2016, 90, 549–561. [Google Scholar] [CrossRef]
- Ostardo, E.; Impellizzeri, D.; Cervigni, M.; Porru, D.; Sommariva, M.; Cordaro, M.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; Crupi, R.; et al. Adelmidrol + sodium hyaluronate in IC/BPS or conditions associated to chronic urothelial inflammation. A translational study. Pharmacol. Res. 2018, 134, 16–30. [Google Scholar] [CrossRef]
- Petrosino, S.; Puigdemont, A.; Della Valle, M.; Fusco, M.; Verde, R.; Allarà, M.; Aveta, T.; Orlando, P.; Di Marzo, V. Adelmidrol increases the endogenous concentrations of palmitoylethanolamide in canine keratinocytes and down-regulates an inflammatory reaction in an in vitro model of contact allergic dermatitis. Veter. J. 2016, 207, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Impellizzeri, D.; Cordaro, M.; Gugliandolo, E.; Peritore, A.F.; Di Paola, R.; Cuzzocrea, S. Topical Application of Adelmidrol + Trans-Traumatic Acid Enhances Skin Wound Healing in a Streptozotocin-Induced Diabetic Mouse Model. Front. Pharmacol. 2018, 9, 871. [Google Scholar] [CrossRef]
- Nageeb, M.M.; Khatab, M.I.; Abdel-Sameea, A.A.; Teleb, N.A. Adelmidrol protects against non-alcoholic steatohepatitis in mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 393, 777–784. [Google Scholar] [CrossRef]
- Di Paola, R.; Fusco, R.; Impellizzeri, D.; Cordaro, M.; Britti, D.; Morittu, V.; Evangelista, M.; Cuzzocrea, S. Adelmidrol, in combination with hyaluronic acid, displays increased anti-inflammatory and analgesic effects against monosodium iodoacetate-induced osteoarthritis in rats. Arthritis Res. 2016, 18, 291. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Cordaro, M.; Genovese, T.; Impellizzeri, D.; Siracusa, R.; Gugliandolo, E.; Peritore, A.F.; D’Amico, R.; Crupi, R.; Cuzzocrea, S.; et al. Adelmidrol: A New Promising Antioxidant and Anti-Inflammatory Therapeutic Tool in Pulmonary Fibrosis. Antioxidants 2020, 9, 601. [Google Scholar] [CrossRef]
- Campolo, M.; Siracusa, R.; Cordaro, M.; Filippone, A.; Gugliandolo, E.; Peritore, A.F.; Impellizzeri, D.; Crupi, R.; Paterniti, I.; Cuzzocrea, S. The association of adelmidrol with sodium hyaluronate displays beneficial properties against bladder changes following spinal cord injury in mice. PLoS ONE 2019, 14, e0208730. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Siracusa, R.; Impellizzeri, D.; Amico, R.D.; Peritore, A.F.; Crupi, R.; Gugliandolo, E.; Fusco, R.; Di Paola, R.; Schievano, C.; et al. Safety and efficacy of a new micronized formulation of the ALIAmide palmitoylglucosamine in preclinical models of inflammation and osteoarthritis pain. Arthritis Res. 2019, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Martini, F.M.; De Bellesini, A.B.; Miolo, A.; Del Coco, L.; Fanizzi, F.P.; Crovace, A. Combining a joint health supplement with tibial plateau leveling osteotomy in dogs with cranial cruciate ligament rupture. An exploratory controlled trial. Int. J. Veter. Sci. Med. 2017, 5, 105–112. [Google Scholar] [CrossRef]
- Deutsch, D.G.; Ueda, N.; Yamamoto, S. The fatty acid amide hydrolase (FAAH). Prostaglandins Leukot. Essent. Fat. Acids 2002, 66, 201–210. [Google Scholar] [CrossRef]
- Waly, N.E.; Refaiy, A.; Aborehab, N.M. IL-10 and TGF-β: Roles in chondroprotective effects of Glucosamine in experimental Osteoarthritis? Pathophysiology 2017, 24, 45–49. [Google Scholar] [CrossRef]
- Henrotin, Y.; Marty, M.; Mobasheri, A. What is the current status of chondroitin sulfate and glucosamine for the treatment of knee osteoarthritis? Maturitas 2014, 78, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Haywood, A.R.; Hathway, G.; Chapman, V. Differential contributions of peripheral and central mechanisms to pain in a rodent model of osteoarthritis. Sci. Rep. 2018, 8, 7122. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Dickenson, A.H.; Baron, R. Osteoarthritis pain: Nociceptive or neuropathic? Nat. Rev. Rheumatol. 2014, 10, 374–380. [Google Scholar] [CrossRef]
- Orita, S.; Ishikawa, T.; Miyagi, M.; Ochiai, N.; Inoue, G.; Eguchi, Y.; Kamoda, H.; Arai, G.; Toyone, T.; Aoki, Y.; et al. Pain-related sensory innervation in monoiodoacetate-induced osteoarthritis in rat knees that gradually develops neuronal injury in addition to inflammatory pain. BMC Musculoskelet. Disord. 2011, 12, 134. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Crupi, R.; Siracusa, R.; Esposito, E.; Cuzzocrea, S. Erratum to: Micronized/ultramicronized palmitoylethanolamide displays superior oral efficacy compared to nonmicronized palmitoylethanolamide in a rat model of inflammatory pain. J. Neuroinflamm. 2016, 13, 129. [Google Scholar] [CrossRef]
- Petrosino, S.; Cordaro, M.; Verde, R.; Moriello, A.S.; Marcolongo, G.; Schievano, C.; Siracusa, R.; Piscitelli, F.; Peritore, A.F.; Crupi, R.; et al. Oral Ultramicronized Palmitoylethanolamide: Plasma and Tissue Levels and Spinal Anti-hyperalgesic Effect. Front. Pharmacol. 2018, 9, 249. [Google Scholar] [CrossRef]
- Koeppen, A.H. Wallerian degeneration: History and clinical significance. J. Neurol. Sci. 2004, 220, 115–117. [Google Scholar] [CrossRef]
- Costa, B.; Comelli, F.; Bettoni, I.; Colleoni, M.; Giagnoni, G. The endogenous fatty acid amide, palmitoylethanolamide, has anti-allodynic and anti-hyperalgesic effects in a murine model of neuropathic pain: Involvement of CB1, TRPV1 and PPARγ receptors and neurotrophic factors. Pain 2008, 139, 541–550. [Google Scholar] [CrossRef]
- Bettoni, I.; Comelli, F.; Colombo, A.; Bonfanti, P.; Costa, B. Non-neuronal cell modulation relieves neuropathic pain: Efficacy of the endogenous lipid palmitoylethanolamide. CNS Neurol. Disord. Drug Targets 2013, 12, 34–44. [Google Scholar] [CrossRef]
- Zuo, Y.; Perkins, N.M.; Tracey, D.; Geczy, C.L. Inflammation and hyperalgesia induced by nerve injury in the rat: A key role of mast cells. Pain 2003, 105, 467–479. [Google Scholar] [CrossRef]
- Petrosino, S.; Palazzo, E.; De Novellis, V.; Bisogno, T.; Rossi, F.; Maione, S.; Di Marzo, V. Changes in spinal and supraspinal endocannabinoid levels in neuropathic rats. Neuropharmacology 2007, 52, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Guida, F.; Luongo, L.; Marmo, F.; Romano, R.; Iannotta, M.; Napolitano, F.; Belardo, C.; Marabese, I.; D’Aniello, A.; De Gregorio, D.; et al. Palmitoylethanolamide reduces pain-related behaviors and restores glutamatergic synapses homeostasis in the medial prefrontal cortex of neuropathic mice. Mol. Brain 2015, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Rahn, E.J.; Guzman-Karlsson, M.C.; Sweatt, J.D. Cellular, molecular, and epigenetic mechanisms in non-associative conditioning: Implications for pain and memory. Neurobiol. Learn. Mem. 2013, 105, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Boccella, S.; Cristiano, C.; Romano, R.; Iannotta, M.; Belardo, C.; Farina, A.; Guida, F.; Piscitelli, F.; Palazzo, E.; Mazzitelli, M.; et al. Ultra-micronized palmitoylethanolamide rescues the cognitive decline-associated loss of neural plasticity in the neuropathic mouse entorhinal cortex-dentate gyrus pathway. Neurobiol. Dis. 2019, 121, 106–119. [Google Scholar] [CrossRef]
- Gugliandolo, E.; D’Amico, R.; Cordaro, M.; Fusco, R.; Siracusa, R.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. Effect of PEA-OXA on neuropathic pain and functional recovery after sciatic nerve crush. J. Neuroinflamm. 2018, 15, 264. [Google Scholar] [CrossRef]
- Peritore, A.F.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; D’Amico, R.; Cordaro, M.; Crupi, R.; Genovese, T.; Impellizzeri, D.; Cuzzocrea, S.; et al. Ultramicronized Palmitoylethanolamide and Paracetamol, a New Association to Relieve Hyperalgesia and Pain in a Sciatic Nerve Injury Model in Rat. Int. J. Mol. Sci. 2020, 21, 3509. [Google Scholar] [CrossRef] [PubMed]
- Pieralice, S.; Vari, R.; Minutolo, A.; Maurizi, A.R.; Fioriti, E.; Napoli, N.; Pozzilli, P.; Manfrini, S.; Maddaloni, E. Biomarkers of response to alpha-lipoic acid ± palmitoiletanolamide treatment in patients with diabetes and symptoms of peripheral neuropathy. Endocrine 2019, 66, 178–184. [Google Scholar] [CrossRef]
- Greene, D.A.; Sima, A.A.F.; Stevens, M.J.; Feldman, E.L.; A Lattimer, S. Complications: Neuropathy, Pathogenetic Considerations. Diabetes Care 1992, 15, 1902–1925. [Google Scholar] [CrossRef]
- Kiyani, M.; Yang, Z.; Charalambous, L.T.; Adil, S.M.; Lee, H.-J.; Yang, S.; Pagadala, P.; Parente, B.; Spratt, S.E.; Lad, S.P. Painful diabetic peripheral neuropathy: Health care costs and complications from 2010 to 2015. Neurol. Clin. Pr. 2020, 10, 47–57. [Google Scholar] [CrossRef]
- Bodman, M.A.; Varacallo, M. Peripheral Diabetic Neuropathy; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Pham, V.M.; Matsumura, S.; Katano, T.; Funatsu, N.; Ito, S. Diabetic neuropathy research: From mouse models to targets for treatment. Neural Regen. Res. 2019, 14, 1870–1879. [Google Scholar] [CrossRef]
- O’Brien, P.D.; Sakowski, S.A.; Feldman, E.L. Mouse models of diabetic neuropathy. ILAR J. 2014, 54, 259–272. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Peritore, A.F.; Cordaro, M.; Gugliandolo, E.; Siracusa, R.; Crupi, R.; D’Amico, R.; Fusco, R.; Evangelista, M.; Cuzzocrea, S.; et al. The neuroprotective effects of micronized PEA (PEA-m) formulation on diabetic peripheral neuropathy in mice. FASEB J. 2019, 33, 11364–11380. [Google Scholar] [CrossRef]
- Darmani, N.A.; Izzo, A.A.; Degenhardt, B.; Valenti, M.; Scaglione, G.; Capasso, R.; Sorrentini, I.; Di Marzo, V. Involvement of the cannabimimetic compound, N-palmitoyl-ethanolamine, in inflammatory and neuropathic conditions: Review of the available pre-clinical data, and first human studies. Neuropharmacology 2005, 48, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, L.D.C.; Pacini, A.; Corti, F.; Boccella, S.; Luongo, L.; Esposito, E.; Cuzzocrea, S.; Maione, S.; Calignano, A.; Ghelardini, C. Antineuropathic Profile of N-Palmitoylethanolamine in a Rat Model of Oxaliplatin-Induced Neurotoxicity. PLoS ONE 2015, 10, e0128080. [Google Scholar] [CrossRef]
- Beijers, A.J.M.; Mols, F.; Vreugdenhil, G. A systematic review on chronic oxaliplatin-induced peripheral neuropathy and the relation with oxaliplatin administration. Support. Care Cancer 2014, 22, 1999–2007. [Google Scholar] [CrossRef] [PubMed]
- Cavaletti, G.; Marmiroli, P. Management of Oxaliplatin-Induced Peripheral Sensory Neuropathy. Cancers 2020, 12, 1370. [Google Scholar] [CrossRef] [PubMed]
- Grothey, A. Oxaliplatin-safety profile: Neurotoxicity. Semin. Oncol. 2003, 30, 5–13. [Google Scholar] [CrossRef]
- Donvito, G.; Wilkerson, J.L.; Damaj, M.I.; Lichtman, A.H. Palmitoylethanolamide Reverses Paclitaxel-Induced Allodynia in Mice. J. Pharmacol. Exp. Ther. 2016, 359, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Maini, K.; Dua, A. Temporomandibular Joint Syndrome; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Rodriguez-Lopez, M.J.; Fernandez-Baena, M.; Aldaya-Valverde, C. Management of pain secondary to temporomandibular joint syndrome with peripheral nerve stimulation. Pain Physician 2015, 18, 229–236. [Google Scholar]
- Joshi, I.; Mattoo, B.; Mohan, V.K.; Bhatia, R. Aberrant pain modulation in trigeminal neuralgia patients. J. Basic Clin. Physiol. Pharmacol. 2020. [Google Scholar] [CrossRef]
- E Cairns, B.; Laursen, J.C.; Dong, X.-D.; Gazerani, P. Intraganglionic injection of a nitric oxide donator induces afferent mechanical sensitization that is attenuated by palmitoylethanolamide. Cephalalgia 2014, 34, 686–694. [Google Scholar] [CrossRef]
- Bartolucci, M.L.; Marini, I.; Bortolotti, F.; Impellizzeri, D.; Di Paola, R.; Bruschetta, G.; Crupi, R.; Portelli, M.; Militi, A.; Oteri, G.; et al. Micronized palmitoylethanolamide reduces joint pain and glial cell activation. Inflamm. Res. 2018, 67, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Alkali, N.H.; Al-Tahan, A.M.; Al-Majed, M.; Al-Tahan, H. Complex Regional Pain Syndrome: A Case Report and Review of the Literature. Ann. Afr. Med. 2020, 19, 68–70. [Google Scholar] [CrossRef] [PubMed]
- Birklein, F.; Ibrahim, A.; Schlereth, T.; Kingery, W.S. The Rodent Tibia Fracture Model: A Critical Review and Comparison with the Complex Regional Pain Syndrome Literature. J. Pain 2018, 19, 1102.e1–1102.e19. [Google Scholar] [CrossRef] [PubMed]
- De Mos, M.; De Bruijn, A.G.; Huygen, F.J.; Dieleman, J.P.; Stricker, C.B.; Sturkenboom, M.C. The incidence of complex regional pain syndrome: A population-based study. Pain 2007, 129, 12–20. [Google Scholar] [CrossRef]
- Wei, T.; Guo, T.-Z.; Li, W.-W.; Kingery, W.S.; Clark, J.D. Acute versus chronic phase mechanisms in a rat model of CRPS. J. Neuroinflamm. 2016, 13, 14. [Google Scholar] [CrossRef]
- Fusco, R.; Gugliandolo, E.; Campolo, M.; Evangelista, M.; Di Paola, R.; Cuzzocrea, S. Correction: Effect of a new formulation of micronized and ultramicronized N-palmitoylethanolamine in a tibia fracture mouse model of complex regional pain syndrome. PLoS ONE 2018, 13, e0201501. [Google Scholar] [CrossRef]
- Mulleman, D.; Mammou, S.; Griffoul, I.; Watier, H.; Goupille, P. Pathophysiology of disk-related sciatica. I.—Evidence supporting a chemical component. Jt. Bone Spine 2006, 73, 151–158. [Google Scholar] [CrossRef]
- Faig-Martí, J.; Martínez-Catassús, A. Use of palmitoylethanolamide in carpal tunnel syndrome: A prospective randomized study. J. Orthop. Traumatol. 2017, 18, 451–455. [Google Scholar] [CrossRef]
- Burton, C.; Chesterton, L.; Davenport, G. Diagnosing and managing carpal tunnel syndrome in primary care. Br. J. Gen. Pr. 2014, 64, 262–263. [Google Scholar] [CrossRef]
- Park, T.; Kuo, A.; Smith, M.T. Chronic low back pain: A mini-review on pharmacological management and pathophysiological insights from clinical and pre-clinical data. Inflammopharmacology 2018, 26, 881–898. [Google Scholar] [CrossRef]
- Kostadinović, S.; Milovanović, N.; Jovanović, J.; Tomašević-Todorović, S. Efficacy of the lumbar stabilization and thoracic mobilization exercise program on pain intensity and functional disability reduction in chronic low back pain patients with lumbar radiculopathy: A randomized controlled trial. J. Back Musculoskelet. Rehabilit. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.A.; Elia, C.; Saini, H.S.; E Miulli, D. A Review of Lumbar Radiculopathy, Diagnosis, and Treatment. Cureus 2019, 11, e5934. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, C.M.; Martín, A.D.; Ferrer, F.G.; Puertas, M.I.; Muro, A.L.; González, J.M.; Prieto, J.P.; Taberna, I.R. N-palmitoylethanolamide in the treatment of neuropathic pain associated with lumbosciatica. Pain Manag. 2012, 2, 119–124. [Google Scholar] [CrossRef]
- Morera, C.; Sabatés, S.; Jaén, Á. Sex differences in N-palmitoylethanolamide effectiveness in neuropathic pain associated with lumbosciatalgia. Pain Manag. 2015, 5, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Chirchiglia, D.; Paventi, S.; Seminara, P.; Cione, E.; Gallelli, L. N-Palmitoyl Ethanol Amide Pharmacological Treatment in Patients With Nonsurgical Lumbar Radiculopathy. J. Clin. Pharmacol. 2018, 58, 733–739. [Google Scholar] [CrossRef]
- Conigliaro, R.; Drago, V.; Foster, P.S.; Schievano, C.; Di Marzo, V. Use of palmitoylethanolamide in the entrapment neuropathy of the median in the wrist. Minerva Med. 2011, 102, 141–147. [Google Scholar]
- Evangelista, M.; Cilli, V.; De Vitis, R.; Militerno, A.; Fanfani, F. Ultra-micronized Palmitoylethanolamide Effects on Sleep-wake Rhythm and Neuropathic Pain Phenotypes in Patients with Carpal Tunnel Syndrome: An Open-label, Randomized Controlled Study. CNS Neurol. Disord. Drug Targets 2018, 17, 291–298. [Google Scholar] [CrossRef]
- Graziani, F.; D’Aiuto, F.; Arduino, P.G.; Tonelli, M.; Gabriele, M. Perioperative dexamethasone reduces post-surgical sequelae of wisdom tooth removal. A split-mouth randomized double-masked clinical trial. Int. J. Oral Maxillofac. Surg. 2006, 35, 241–246. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L.; Fusco, M.; Della Valle, M.F.; Zusso, M.; Costa, B.; Giusti, P. Palmitoylethanolamide, a naturally occurring disease-modifying agent in neuropathic pain. Inflammopharmacology 2013, 22, 79–94. [Google Scholar] [CrossRef]
- Gatti, A.; Lazzari, M.; Gianfelice, V.; Di Paolo, A.; Sabato, E.; Sabato, A.F. Palmitoylethanolamide in the Treatment of Chronic Pain Caused by Different Etiopathogenesis. Pain Med. 2012, 13, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Bacci, C.; Cassetta, G.; Emanuele, B.; Berengo, M. Randomized Split-Mouth Study on Postoperative Effects of Palmitoylethanolamide for Impacted Lower Third Molar Surgery. ISRN Surg. 2011, 2011, 1–6. [Google Scholar] [CrossRef]
- Truini, A.; Biasiotta, A.; Di Stefano, G.; La Cesa, S.; Leone, C.; Cartoni, C.; Federico, V.; Petrucci, M.T.; Cruccu, G. Palmitoylethanolamide restores myelinated-fibre function in patients with chemotherapy-induced painful neuropathy. CNS Neurol. Disord. Drug Targets 2011, 10, 916–920. [Google Scholar] [CrossRef] [PubMed]
- Schifilliti, C.; Cucinotta, L.; Fedele, V.; Ingegnosi, C.; Luca, S.; Leotta, C. Micronized Palmitoylethanolamide Reduces the Symptoms of Neuropathic Pain in Diabetic Patients. Pain Res. Treat. 2014, 2014, 1–5. [Google Scholar] [CrossRef]
- Cocito, D.; Peci, E.; Ciaramitaro, P.; Merola, A.; Lopiano, L. Short-Term Efficacy of Ultramicronized Palmitoylethanolamide in Peripheral Neuropathic Pain. Pain Res. Treat. 2014, 2014, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Passavanti, M.B.; Fiore, M.; Sansone, P.; Aurilio, C.; Pota, V.; Barbarisi, M.; Fierro, D.; Pace, M.C. The beneficial use of ultramicronized palmitoylethanolamide as add-on therapy to Tapentadol in the treatment of low back pain: A pilot study comparing prospective and retrospective observational arms. BMC Anesthesiol. 2017, 17, 171. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, V.; Martini, A.; Bellamoli, P.; Donadello, K.; Schievano, C.; Balzo, G.D.; Sarzi-Puttini, P.; Parolini, M.; Polati, E. Ultramicronized Palmitoylethanolamide (um-PEA) as Add-on Treatment in Fibromyalgia Syndrome (FMS): Retrospective Observational Study on 407 Patients. CNS Neurol. Disord. Drug Targets 2019, 18, 326–333. [Google Scholar] [CrossRef]
- Jahan, F.; Nanji, K.; Qidwai, W.; Qasim, R. Fibromyalgia Syndrome: An Overview of Pathophysiology, Diagnosis and Management. Oman Med. J. 2012, 27, 192–195. [Google Scholar] [CrossRef]
- Paladini, A.; Fusco, M.; Cenacchi, T.; Schievano, C.; Piroli, A.; Varrassi, G. Palmitoylethanolamide, a Special Food for Medical Purposes, in the Treatment of Chronic Pain: A Pooled Data Meta-analysis. Pain Physician 2016, 19, 11–24. [Google Scholar]



{kind=link}
{kind=link}
| Central Neuropathic Pain | Peripheral Neuropathic Pain |
|---|---|
| Spinal cord injury | Diabetic painful neuropathy (DPN) |
| Central post-stroke pain | Alcoholic neuropathy |
| Parkinson disease pain | Cancer pain |
| Multiple sclerosis-related pain | Chemotherapy-induced peripheral neuropathy (CIPN) |
| Transverse myelitis | Charcot-Marie-Tooth disease |
| Neuromyelitis optica | Trigeminal neuralgia |
| Syringomyelia | Acute e chronic inflammatory demyelinating polyneuropathy |
| Human immunodeficiency virus-associated neuropathy | |
| Post-traumatic neuropathy | |
| Phantom limb pain Compressive mononeuropathies and many more. |
| Ion Channels | Na, Ca, K |
|---|---|
| Ionotropic and metabotropic receptors | Glutamatergic, GABA (γ-aminobutyric acid) ergic, serotoninergic, adrenergic |
| Inflammatory cytokines | IL-1β, IL-6, TNF-α |
| Nerve growth factors | NGF |
| Intracellular regulators | protein kinase C |
| Transcriptional factors | nuclear factor-κB |
| Allodynia | Hyperalgesia |
|---|---|
| refers to pain produced by a normally non-painful stimulus, and it may result from decreased stimulation thresholds | refers to exaggerated pain perception as a result of damaged peripheral pain fibers |
| Classified: -mechanical (pain in response to light touch)—thermal (hot or cold: pain from normally mild skin temperatures in the affected area) -movement pain triggered by normal movement of joints or muscles | Classified: -primary hyperalgesia: occurs directly in injured tissue as a result of sensitization of peripheral nociceptors (for example, tenderness after a cut), -secondary hyperalgesia: occurs in adjacent undamaged tissue owing to sensitization within the CNS |
| An example is a patient with diabetic neuropathy whose feet are sensitive to putting on socks. | A clinical example of hyperalgesia might be an amputee who is unable to use a prosthesis because of tenderness overlying the stump. |
| Protagonists | Function | References |
|---|---|---|
| Astrocytes | Involved in brain homeostasis, provide metabolites and growth factors to neurons, support synapse formation and plasticity, participate in BBB maintenance and permeability. | [61,62,63] |
| Oligondencytes | Involved in the formation of myelin, propagation of action potentials along axons, production of neurotrophic factors that support to neurons | [55,64] |
| Inflammasomes | Involved in induction of the pyroptosis process, responsible for the secretion of the inflammatory cytokines | [65,66] |
| Cytokines and chemokines | Involved in tissue repair and homeostasis restoration, responsible to cell migration | [67,68] |
| Drug Class | Kind of Neuropathic Pain | Effects | Side Effects | References | |
|---|---|---|---|---|---|
| First-Line Drugs | |||||
| Antidepressants | -Tricyclic antidepressants (TCAs): amitriptyline, nortriptyline, desipramine, imipramine -Serotonin-norepinephrine reuptake inhibitors (SNRIs): Duloxetina Venlafaxina | Painful diabetic neuropathy Post-herpetic neuralgia Post-stroke pain Painful polyneuropathy Low back pain | Inhibition the reuptake of serotonin and noradrenaline into the spinal synapses between nociceptors (or first-order neurons) and the spinothalamic neurons (or second-order neurons) | Sedation Heart problems Constipation Drowsiness Light-headedness Weight gain Dry mouth Nausea (SNRIs less side effects than TCAs) | [76,77,78] |
| Anticonvulsants | -Phenytoin -Gabapentin -Carbamazepine -Oxcarbazepine -Valproic acid | Lancinating pain and allodynia Painful diabetic neuropathy Trigeminal neuralgia Post-herpetic neuralgia Painful polyneuropathy Low back pain | Reduction of neuronal excitability and local neuronal discharges, acting through sodium channel blockade or modulation of calcium channels | Dizziness Somnolence Skin reactions such as Stevens–Johnson syndrome Leukopenia Hyponatremia | [79,80,81,82] |
| Second- and Third-Line Drugs | |||||
| Topical agents | -Lidocaine -Capsaicin -Clonidine -EMLA (eutectic mixture of local anesthetics) | Allodynia Post-herpetic neuralgia Chemotherapy-induced peripheral neuropathy Post-surgical and post-traumatic neuropathic pain | Block of voltage-gated sodium channels expressed by nerve fibers, responsible for the propagation of action potentials. | Local irritation Possible hypersensitivity | [75,83,84,85,86] |
| Opioids | -Morphine -Oxycodone, -Hydromorphone -Tramadol | Diabetic peripheral neuropathy Post-herpetic neuropathy Polyneuropathy Phantom limb pain | Opioid receptors are coupled to calcium and potassium channels, block synaptic transmission, restricting the number of nociceptive stimuli | Drowsiness Nausea Dependence Overdoses | [76,87,88,89] |
| Corticosteroids | -Prednisone -Desametasone | Allodynia Spinal cord compression Post-herpetic neuralgia | Inhibition of prostaglandin synthesis, reduction inflammation, vascular permeability and tissue edema | Gastrointestinal disease Psychiatric disorders Electrolyte imbalances Bone demineralization | [90,91] |
| Alternative non-pharmacologic therapies | -Acupuncture -Magnetic insoles -Repetitive transcranial magnetic stimulation (rTMS) | Chemotherapy-induced peripheral neuropathy Trigeminal neuralgia Post-stroke pain Post-herpetic pain | Local inhibition of nociceptive fibres; stimulates blood flow to restore nerve damage. | Bruising Infection | [92,93] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amico, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. ALIAmides Update: Palmitoylethanolamide and Its Formulations on Management of Peripheral Neuropathic Pain. Int. J. Mol. Sci. 2020, 21, 5330. https://doi.org/10.3390/ijms21155330
D’Amico R, Impellizzeri D, Cuzzocrea S, Di Paola R. ALIAmides Update: Palmitoylethanolamide and Its Formulations on Management of Peripheral Neuropathic Pain. International Journal of Molecular Sciences. 2020; 21(15):5330. https://doi.org/10.3390/ijms21155330
Chicago/Turabian StyleD’Amico, Ramona, Daniela Impellizzeri, Salvatore Cuzzocrea, and Rosanna Di Paola. 2020. "ALIAmides Update: Palmitoylethanolamide and Its Formulations on Management of Peripheral Neuropathic Pain" International Journal of Molecular Sciences 21, no. 15: 5330. https://doi.org/10.3390/ijms21155330
APA StyleD’Amico, R., Impellizzeri, D., Cuzzocrea, S., & Di Paola, R. (2020). ALIAmides Update: Palmitoylethanolamide and Its Formulations on Management of Peripheral Neuropathic Pain. International Journal of Molecular Sciences, 21(15), 5330. https://doi.org/10.3390/ijms21155330