Alzheimer’s Disease Mouse as a Model of Testis Degeneration




 ,
,
Abstract
1. Introduction
2. Results
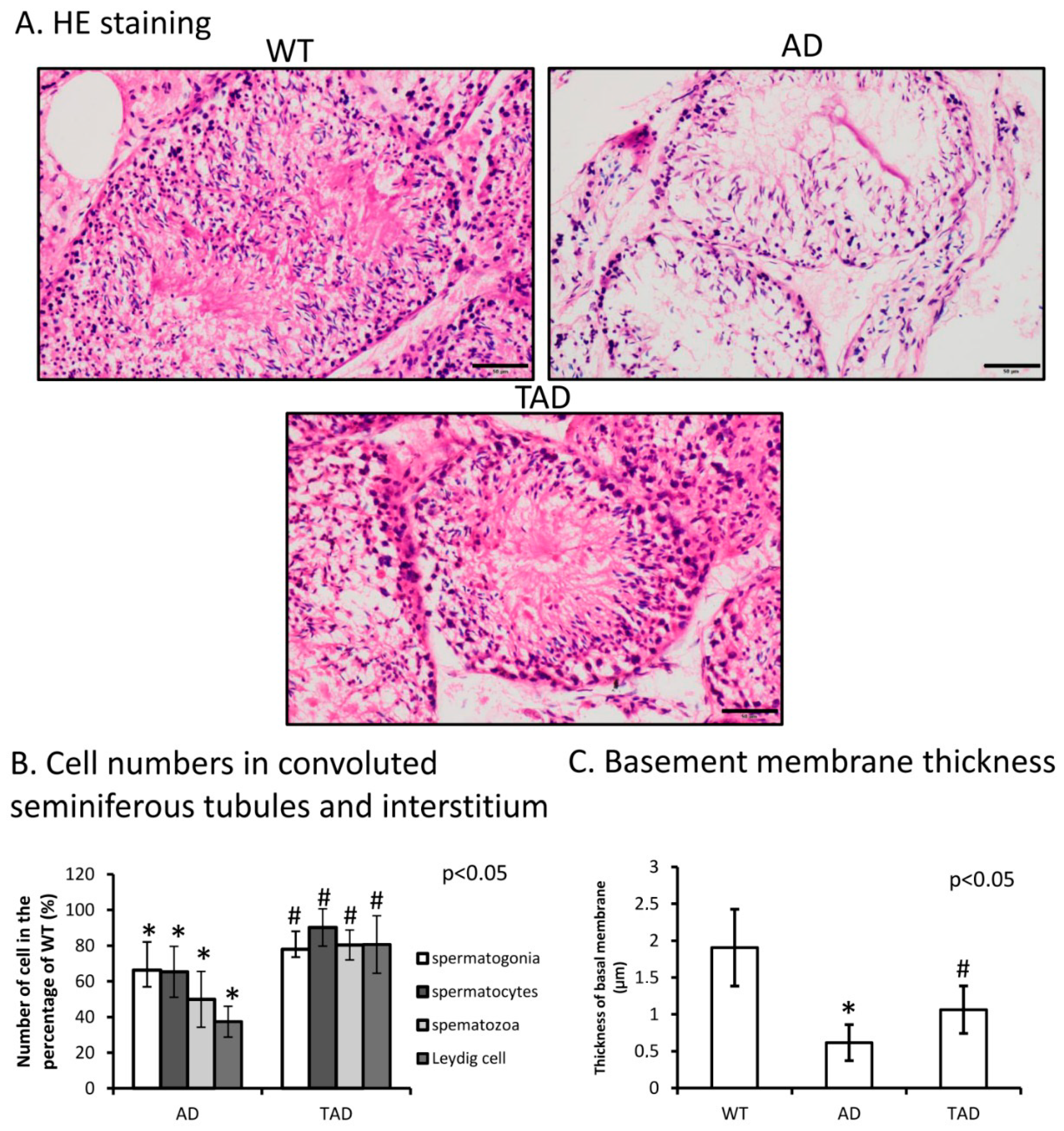
2.1. Decreased Cell Density and Thinned Basement Membrane of Convoluted Seminiferous Tubules in Alzheimer’s Disease
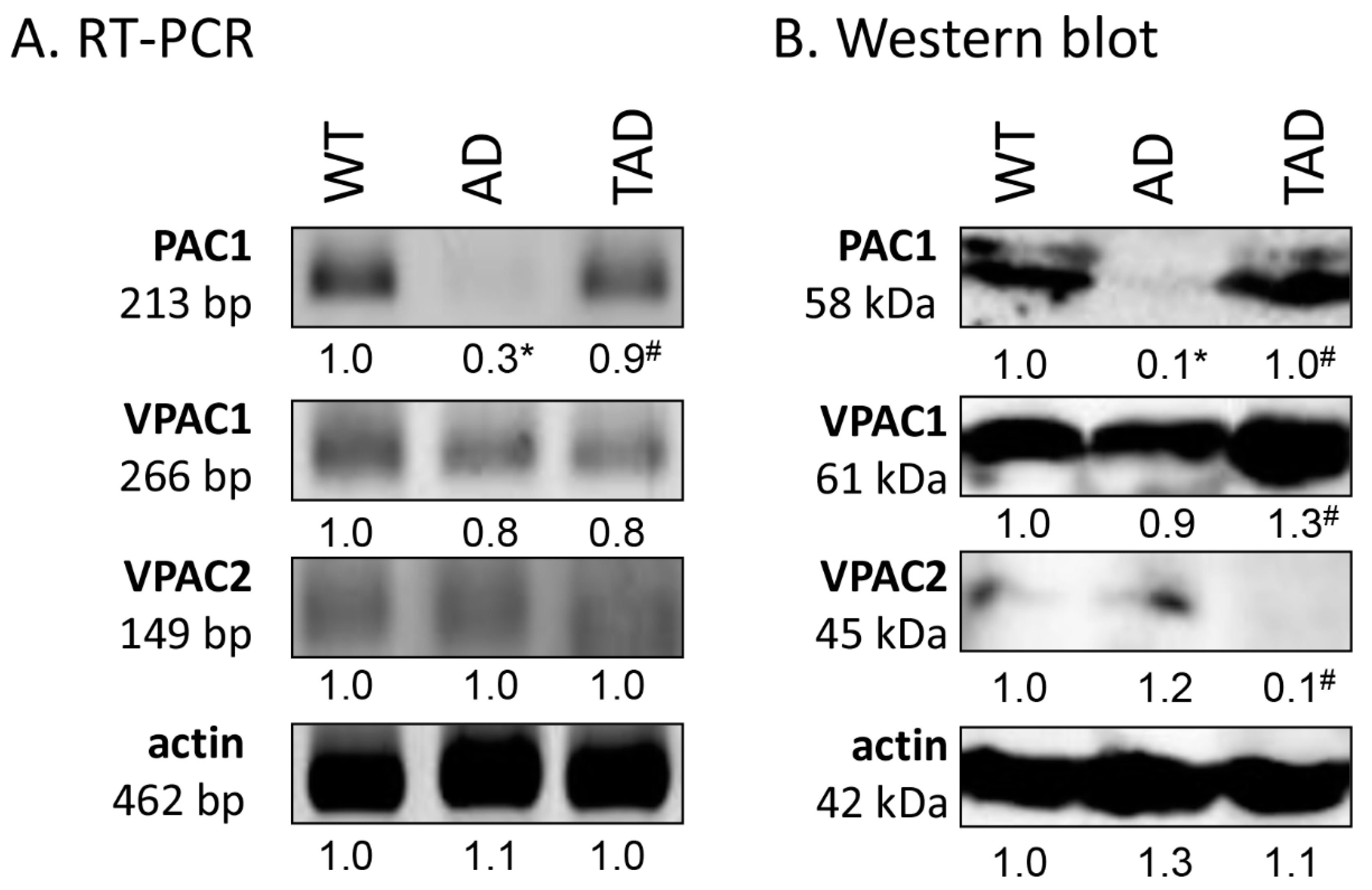
2.2. Decreased PAC1 Receptor Expression in Testis of AD Mice
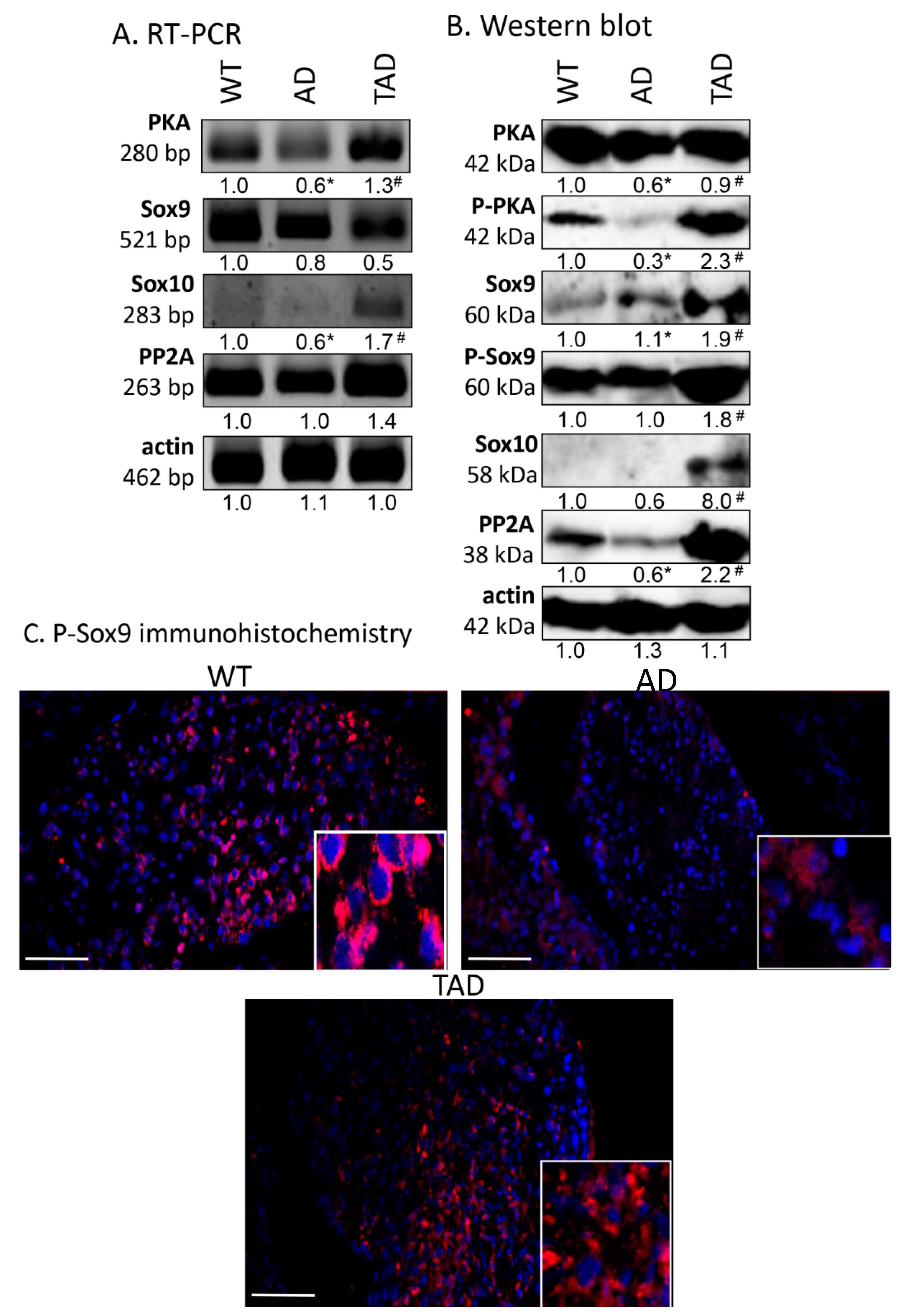
2.3. Reduction in PACAP Signaling Elements in AD Testis
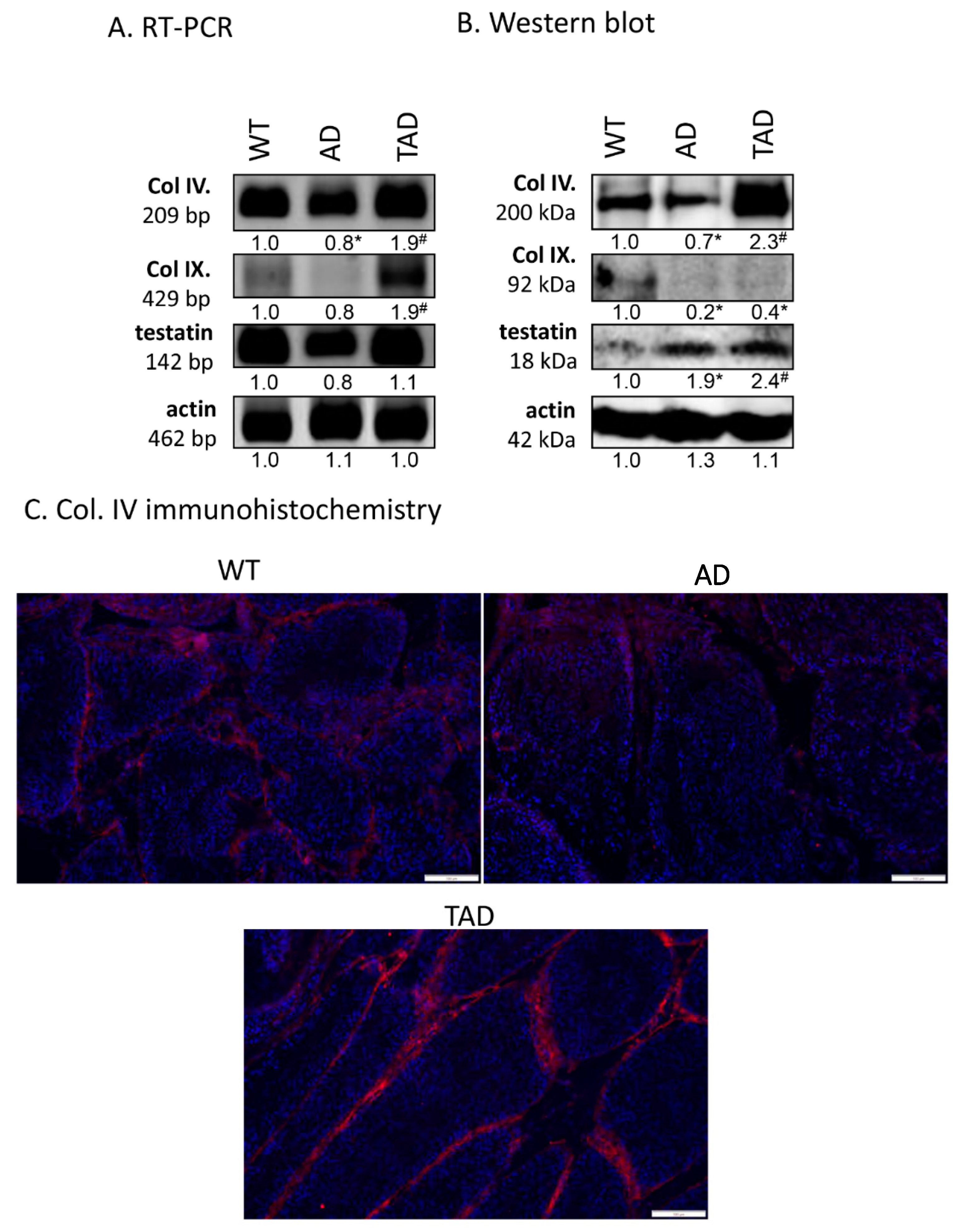
2.4. Expression of Basement Membrane Components Altered in Seminiferous Tubules in AD
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Light Microscopical Morphology
4.3. Immunohistochemistry
4.4. RT-PCR Analysis
4.5. Western Blot Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| PACAP | Pituitary adenylate cyclase activating polypeptide |
| cAMP | Cyclic adenosine monophosphate |
| PKA | Protein kinase A |
| P-PKA | Phosphorylated protein kinase A |
| CNS | Central nervous system |
| Aβ | Beta amyloid |
| APP | Amyloid precursor protein |
| KO mouse | Knock out mouse |
| PP2A | Protein phosphatase 2A |
| mRNA | Messenger ribonucleic acid |
| HE staining | Hematoxylin and eosin staining |
| Sox9 | SRY-related HMG-box 9 |
| WT mouse | Wild type mouse |
| AD mouse | Alzheimer’s disease mouse |
| TAD mouse | Trained Alzheimer’s disease mouse |
| VPAC | Vasoactive intestinal peptide receptor |
| PAC1 | Pituitary adenylate cyclase-activating polypeptide type I receptor |
References
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharm. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Banki, E.; Kovacs, K.; Nagy, D.; Juhasz, T.; Degrell, P.; Csanaky, K.; Kiss, P.; Jancso, G.; Toth, G.; Tamas, A.; et al. Molecular mechanisms underlying the nephroprotective effects of PACAP in diabetes. J. Mol. Neurosci. 2014, 54, 300–309. [Google Scholar] [CrossRef]
- Clason, T.A.; Girard, B.M.; May, V.; Parsons, R.L. Activation of MEK/ERK signaling by PACAP in guinea pig cardiac neurons. J. Mol. Neurosci. 2016, 59, 309–316. [Google Scholar] [CrossRef]
- Jozsa, G.; Szegeczki, V.; Palfi, A.; Kiss, T.; Helyes, Z.; Fulop, B.; Cserhati, C.; Daroczi, L.; Tamas, A.; Zakany, R.; et al. Signalling alterations in bones of pituitary adenylate cyclase activating polypeptide (PACAP) gene deficient mice. Int. J. Mol. Sci. 2018, 19, 2538. [Google Scholar] [CrossRef]
- Fulop, B.D.; Sandor, B.; Szentleleky, E.; Karanyicz, E.; Reglodi, D.; Gaszner, B.; Zakany, R.; Hashimoto, H.; Juhasz, T.; Tamas, A. Altered Notch signaling in developing molar teeth of pituitary adenylate cyclase-activating polypeptide (PACAP)-deficient mice. J. Mol. Neurosci. 2019, 68, 377–388. [Google Scholar] [CrossRef]
- Laszlo, E.; Juhasz, T.; Varga, A.; Czibere, B.; Kovacs, K.; Degrell, P.; Horvath, G.; Jancso, G.; Szakaly, P.; Tamas, A.; et al. Protective effect of PACAP on ischemia/reperfusion-induced kidney injury of male and female rats: Gender differences. J. Mol. Neurosci. 2019, 68, 408–419. [Google Scholar] [CrossRef]
- Reglodi, D.; Cseh, S.; Somoskoi, B.; Fulop, B.D.; Szentleleky, E.; Szegeczki, V.; Kovacs, A.; Varga, A.; Kiss, P.; Hashimoto, H.; et al. Disturbed spermatogenic signaling in pituitary adenylate cyclase activating polypeptide-deficient mice. Reproduction 2018, 155, 129–139. [Google Scholar] [CrossRef]
- Juhasz, T.; Matta, C.; Katona, E.; Somogyi, C.; Takacs, R.; Gergely, P.; Csernoch, L.; Panyi, G.; Toth, G.; Reglodi, D.; et al. Pituitary adenylate cyclase activating polypeptide (PACAP) signalling exerts chondrogenesis promoting and protecting effects: Implication of calcineurin as a downstream target. PLoS ONE 2014, 9, e91541. [Google Scholar] [CrossRef]
- Nakamura, K.; Nakamachi, T.; Endo, K.; Ito, K.; Machida, T.; Oka, T.; Hori, M.; Ishizaka, K.; Shioda, S. Distribution of pituitary adenylate cyclase-activating polypeptide (PACAP) in the human testis and in testicular germ cell tumors. Andrologia 2014, 46, 465–471. [Google Scholar] [CrossRef]
- Brubel, R.; Kiss, P.; Vincze, A.; Varga, A.; Varnagy, A.; Bodis, J.; Mark, L.; Jambor, E.; Maasz, G.; Hashimoto, H.; et al. Effects of pituitary adenylate cyclase activating polypeptide on human sperm motility. J. Mol. Neurosci. 2012, 48, 623–630. [Google Scholar] [CrossRef]
- Prisco, M.; Rosati, L.; Morgillo, E.; Mollica, M.P.; Agnese, M.; Andreuccetti, P.; Valiante, S. Pituitary adenylate cyclase-activating peptide (PACAP) and its receptors in Mus musculus testis. Gen. Comp. Endocrinol. 2019, 286, 113297. [Google Scholar] [CrossRef]
- Meggyes, M.; Lajko, A.; Fulop, B.D.; Reglodi, D.; Szereday, L. Phenotypic characterization of testicular immune cells expressing immune checkpoint molecules in wild-type and PACAP-deficient mice. Am. J. Reprod. Immunol. 2019, e13212. [Google Scholar] [CrossRef]
- Ohtaki, H.; Satoh, A.; Nakamachi, T.; Yofu, S.; Dohi, K.; Mori, H.; Ohara, K.; Miyamoto, K.; Hashimoto, H.; Shintani, N.; et al. Regulation of oxidative stress by pituitary adenylate cyclase-activating polypeptide (PACAP) mediated by PACAP receptor. J. Mol. Neurosci. 2010, 42, 397–403. [Google Scholar] [CrossRef]
- de Boisvilliers, M.; Perrin, F.; Hebache, S.; Balandre, A.C.; Bensalma, S.; Garnier, A.; Vaudry, D.; Fournier, A.; Festy, F.; Muller, J.M.; et al. VIP and PACAP analogs regulate therapeutic targets in high-risk neuroblastoma cells. Peptides 2016, 78, 30–41. [Google Scholar] [CrossRef]
- Szegeczki, V.; Bauer, B.; Jungling, A.; Fulop, B.D.; Vago, J.; Perenyi, H.; Tarantini, S.; Tamas, A.; Zakany, R.; Reglodi, D.; et al. Age-related alterations of articular cartilage in pituitary adenylate cyclase-activating polypeptide (PACAP) gene-deficient mice. Geroscience 2019, 41, 775–793. [Google Scholar] [CrossRef]
- Reglodi, D.; Jungling, A.; Longuespee, R.; Kriegsmann, J.; Casadonte, R.; Kriegsmann, M.; Juhasz, T.; Bardosi, S.; Tamas, A.; Fulop, B.D.; et al. Accelerated pre-senile systemic amyloidosis in PACAP knockout mice-a protective role of PACAP in age-related degenerative processes. J. Pathol. 2018, 245, 478–490. [Google Scholar] [CrossRef]
- Lee, E.H.; Seo, S.R. Neuroprotective roles of pituitary adenylate cyclase-activating polypeptide in neurodegenerative diseases. BMB Rep. 2014, 47, 369–375. [Google Scholar] [CrossRef]
- Han, P.; Tang, Z.; Yin, J.; Maalouf, M.; Beach, T.G.; Reiman, E.M.; Shi, J. Pituitary adenylate cyclase-activating polypeptide protects against beta-amyloid toxicity. Neurobiol. Aging 2014, 35, 2064–2071. [Google Scholar] [CrossRef]
- Griffith, C.M.; Eid, T.; Rose, G.M.; Patrylo, P.R. Evidence for altered insulin receptor signaling in Alzheimer’s disease. Neuropharmacology 2018, 136, 202–215. [Google Scholar] [CrossRef]
- Csiszar, A.; Tarantini, S.; Fulop, G.A.; Kiss, T.; Valcarcel-Ares, M.N.; Galvan, V.; Ungvari, Z.; Yabluchanskiy, A. Hypertension impairs neurovascular coupling and promotes microvascular injury: Role in exacerbation of Alzheimer’s disease. Geroscience 2017, 39, 359–372. [Google Scholar] [CrossRef]
- Morris, J.K.; Honea, R.A.; Vidoni, E.D.; Swerdlow, R.H.; Burns, J.M. Is Alzheimer’s disease a systemic disease? Biochim. Biophys. Acta 2014, 1842, 1340–1349. [Google Scholar] [CrossRef]
- Seyedreza, P.; Alireza, M.N.; Seyedebrahim, H. Role of testosterone in memory impairment of Alzheimer disease induced by Streptozotocin in male rats. Daru 2012, 20, 98. [Google Scholar] [CrossRef]
- Silva, J.V.; Yoon, S.; Domingues, S.; Guimaraes, S.; Goltsev, A.V.; da Cruz, E.S.E.F.; Mendes, J.F.; da Cruz, E.S.O.A.; Fardilha, M. Amyloid precursor protein interaction network in human testis: Sentinel proteins for male reproduction. BMC Bioinform. 2015, 16, 12. [Google Scholar] [CrossRef]
- Shoji, M.; Kawarabayashi, T.; Harigaya, Y.; Yamaguchi, H.; Hirai, S.; Kamimura, T.; Sugiyama, T. Alzheimer amyloid beta-protein precursor in sperm development. Am. J. Pathol. 1990, 137, 1027–1032. [Google Scholar]
- Whelly, S.; Johnson, S.; Powell, J.; Borchardt, C.; Hastert, M.C.; Cornwall, G.A. Nonpathological extracellular amyloid is present during normal epididymal sperm maturation. PLoS ONE 2012, 7, e36394. [Google Scholar] [CrossRef]
- Micera, A.; Bruno, L.; Cacciamani, A.; Rongioletti, M.; Squitti, R. Alzheimer’s disease and retinal degeneration: A glimpse at essential trace metals in ocular fluids and tissues. Curr. Alzheimer Res. 2019, 16, 1073–1083. [Google Scholar] [CrossRef]
- Bhattarai, P.; Thomas, A.K.; Cosacak, M.I.; Papadimitriou, C.; Mashkaryan, V.; Zhang, Y.; Kizil, C. Modeling amyloid-beta42 toxicity and neurodegeneration in adult zebrafish brain. J. Vis. Exp. 2017, 128, 56014. [Google Scholar] [CrossRef]
- Weissmann, R.; Huttenrauch, M.; Kacprowski, T.; Bouter, Y.; Pradier, L.; Bayer, T.A.; Kuss, A.W.; Wirths, O. Gene expression profiling in the APP/PS1KI mouse model of familial Alzheimer’s disease. J. Alzheimers Dis. 2016, 50, 397–409. [Google Scholar] [CrossRef]
- Collins, J.M.; King, A.E.; Woodhouse, A.; Kirkcaldie, M.T.; Vickers, J.C. The effect of focal brain injury on beta-amyloid plaque deposition, inflammation and synapses in the APP/PS1 mouse model of Alzheimer’s disease. Exp. Neurol. 2015, 267, 219–229. [Google Scholar] [CrossRef]
- Reiserer, R.S.; Harrison, F.E.; Syverud, D.C.; McDonald, M.P. Impaired spatial learning in the APPSwe + PSEN1DeltaE9 bigenic mouse model of Alzheimer’s disease. Genes Brain Behav. 2007, 6, 54–65. [Google Scholar] [CrossRef]
- Ghate, P.S.; Sidhar, H.; Carlson, G.A.; Giri, R.K. Development of a novel cellular model of Alzheimer’s disease utilizing neurosphere cultures derived from B6C3-Tg(APPswe,PSEN1dE9)85Dbo/J embryonic mouse brain. Springerplus 2014, 3, 161. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.M.; Sichler, M.E.; Schleicher, E.M.; Franke, T.N.; Irwin, C.; Low, M.J.; Beindorff, N.; Bouter, C.; Bayer, T.A.; Bouter, Y. Analysis of motor function in the Tg4-42 mouse model of Alzheimer’s disease. Front. Behav. Neurosci. 2019, 13, 107. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, P.L.; Castillo-Garcia, A.; Morales, J.S.; de la Villa, P.; Hampel, H.; Emanuele, E.; Lista, S.; Lucia, A. Exercise benefits on Alzheimer’s disease: State-of-the-science. Ageing Res. Rev. 2020, 62, 101108. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Lin, M.S.; Tzeng, I.S. Relationship between exercise and Alzheimer’s disease: A narrative literature review. Front. Neurosci. 2020, 14, 131. [Google Scholar] [CrossRef] [PubMed]
- Radak, Z.; Hart, N.; Sarga, L.; Koltai, E.; Atalay, M.; Ohno, H.; Boldogh, I. Exercise plays a preventive role against Alzheimer’s disease. J. Alzheimers Dis. 2010, 20, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Abraham, D.; Feher, J.; Scuderi, G.L.; Szabo, D.; Dobolyi, A.; Cservenak, M.; Juhasz, J.; Ligeti, B.; Pongor, S.; Gomez-Cabrera, M.C.; et al. Exercise and probiotics attenuate the development of Alzheimer’s disease in transgenic mice: Role of microbiome. Exp. Gerontol. 2019, 115, 122–131. [Google Scholar] [CrossRef]
- Torma, F.; Koltai, E.; Nagy, E.; Ziaaldini, M.M.; Posa, A.; Koch, L.G.; Britton, S.L.; Boldogh, I.; Radak, Z. Exercise increases markers of spermatogenesis in rats selectively bred for low running capacity. PLoS ONE 2014, 9, e114075. [Google Scholar] [CrossRef]
- Yang, S.H. Cellular and Molecular Mediators of Neuroinflammation in Alzheimer Disease. Int. Neurourol. J. 2019, 23, S54–62. [Google Scholar] [CrossRef]
- Jensen, C.S.; Bahl, J.M.; Ostergaard, L.B.; Hogh, P.; Wermuth, L.; Heslegrave, A.; Zetterberg, H.; Heegaard, N.H.H.; Hasselbalch, S.G.; Simonsen, A.H. Exercise as a potential modulator of inflammation in patients with Alzheimer’s disease measured in cerebrospinal fluid and plasma. Exp. Gerontol. 2019, 121, 91–98. [Google Scholar] [CrossRef]
- Torvell, M.; Hampton, D.W.; Connick, P.; MacLullich, A.M.J.; Cunningham, C.; Chandran, S. A single systemic inflammatory insult causes acute motor deficits and accelerates disease progression in a mouse model of human tauopathy. Alzheimers Dement. (N. Y.) 2019, 5, 579–591. [Google Scholar] [CrossRef]
- Vinothkumar, G.; Krishnakumar, S.; Sureshkumar; Shivashekar, G.; Sreedhar, S.; Preethikrishnan; Dinesh, S.; Sundaram, A.; Balakrishnan, D.; Riya; et al. Therapeutic impact of rHuEPO on abnormal platelet APP, BACE 1, presenilin 1, ADAM 10 and Abeta expressions in chronic kidney disease patients with cognitive dysfunction like Alzheimer’s disease: A pilot study. Biomed. Pharm. 2018, 104, 211–222. [Google Scholar]
- Press, M.; Jung, T.; Konig, J.; Grune, T.; Hohn, A. Protein aggregates and proteostasis in aging: Amylin and beta-cell function. Mech. Ageing Dev. 2019, 177, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Vaczy, A.; Rubio-Beltran, E.; MaassenVanDenBrink, A. Protective effects of PACAP in ischemia. J. Headache Pain 2018, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, M.; Nakamachi, T.; Sugiyama, K.; Tsuchikawa, D.; Watanabe, J.; Hori, M.; Yoshikawa, A.; Imai, N.; Kagami, N.; Matkovits, A.; et al. PACAP stimulates functional recovery after spinal cord injury through axonal regeneration. J. Mol. Neurosci. 2014, 54, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Waschek, J.A. VIP and PACAP: Neuropeptide modulators of CNS inflammation, injury, and repair. Br. J. Pharm. 2013, 169, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Tamas, A.; Reglodi, D.; Farkas, O.; Kovesdi, E.; Pal, J.; Povlishock, J.T.; Schwarcz, A.; Czeiter, E.; Szanto, Z.; Doczi, T.; et al. Effect of PACAP in central and peripheral nerve injuries. Int. J. Mol. Sci. 2012, 13, 8430–8448. [Google Scholar] [CrossRef]
- Werling, D.; Reglodi, D.; Banks, W.A.; Salameh, T.S.; Kovacs, K.; Kvarik, T.; Vaczy, A.; Kovacs, L.; Mayer, F.; Danyadi, B.; et al. Ocular Delivery of PACAP1-27 Protects the Retina From Ischemic Damage in Rodents. Invest. Ophthalmol Vis. Sci. 2016, 57, 6683–6691. [Google Scholar] [CrossRef]
- Reglodi, D.; Tamas, A.; Lengvari, I.; Toth, G.; Szalontay, L.; Lubics, A. Comparative study of the effects of PACAP in young, aging, and castrated males in a rat model of Parkinson’s disease. Ann. N. Y. Acad. Sci. 2006, 1070, 518–524. [Google Scholar] [CrossRef]
- Chen, X.Y.; Du, Y.F.; Chen, L. Neuropeptides Exert Neuroprotective Effects in Alzheimer’s Disease. Front. Mol. Neurosci. 2018, 11, 493. [Google Scholar] [CrossRef]
- Reglodi, D.; Atlasz, T.; Szabo, E.; Jungling, A.; Tamas, A.; Juhasz, T.; Fulop, B.D.; Bardosi, A. PACAP deficiency as a model of aging. Geroscience 2018, 40, 437–452. [Google Scholar] [CrossRef]
- Lacombe, A.; Lelievre, V.; Roselli, C.E.; Salameh, W.; Lue, Y.H.; Lawson, G.; Muller, J.M.; Waschek, J.A.; Vilain, E. Delayed testicular aging in pituitary adenylate cyclase-activating peptide (PACAP) null mice. Proc. Natl. Acad. Sci. USA 2006, 103, 3793–3798. [Google Scholar] [CrossRef] [PubMed]
- Jozsa, G.; Fulop, B.D.; Kovacs, L.; Czibere, B.; Szegeczki, V.; Kiss, T.; Hajdu, T.; Tamas, A.; Helyes, Z.; Zakany, R.; et al. Lack of pituitary adenylate cyclase-activating polypeptide (PACAP) disturbs callus formation. J. Mol. Neurosci. 2019, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Doan, N.D.; Chatenet, D.; Letourneau, M.; Vaudry, H.; Vaudry, D.; Fournier, A. Receptor-independent cellular uptake of pituitary adenylate cyclase-activating polypeptide. Biochim. Biophys. Acta 2012, 1823, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Dubal, D.B.; Broestl, L.; Worden, K. Sex and gonadal hormones in mouse models of Alzheimer’s disease: What is relevant to the human condition? Biol. Sex. Differ. 2012, 3, 24. [Google Scholar] [CrossRef]
- Szentleleky, E.; Szegeczki, V.; Karanyicz, E.; Hajdu, T.; Tamas, A.; Toth, G.; Zakany, R.; Reglodi, D.; Juhasz, T. Pituitary adenylate cyclase activating polypeptide (PACAP) reduces oxidative and mechanical stress-evoked matrix degradation in chondrifying cell cultures. Int. J. Mol. Sci. 2019, 20, 168. [Google Scholar] [CrossRef]
- Li, M.; Funahashi, H.; Mbikay, M.; Shioda, S.; Arimura, A. Pituitary adenylate cyclase activating polypeptide-mediated intracrine signaling in the testicular germ cells. Endocrine 2004, 23, 59–75. [Google Scholar] [CrossRef]
- Paladini, F.; Porciello, N.; Camilli, G.; Tuncer, S.; Cocco, E.; Fiorillo, M.T.; Sorrentino, R. Single nucleotide polymorphisms in the 3’UTR of VPAC-1 cooperate in modulating gene expression and impact differently on the interaction with miR525-5p. PLoS ONE 2014, 9, e112646. [Google Scholar] [CrossRef]
- Juhasz, T.; Helgadottir, S.L.; Tamas, A.; Reglodi, D.; Zakany, R. PACAP and VIP signaling in chondrogenesis and osteogenesis. Peptides 2015, 66, 51–57. [Google Scholar] [CrossRef]
- Sanchez-Mut, J.V.; Aso, E.; Heyn, H.; Matsuda, T.; Bock, C.; Ferrer, I.; Esteller, M. Promoter hypermethylation of the phosphatase DUSP22 mediates PKA-dependent TAU phosphorylation and CREB activation in Alzheimer’s disease. Hippocampus 2014, 24, 363–368. [Google Scholar] [CrossRef]
- Melling, C.W.; Krause, M.P.; Noble, E.G. PKA-mediated ERK1/2 inactivation and hsp70 gene expression following exercise. J. Mol. Cell Cardiol. 2006, 41, 816–822. [Google Scholar] [CrossRef]
- Jo, A.; Denduluri, S.; Zhang, B.; Wang, Z.; Yin, L.; Yan, Z.; Kang, R.; Shi, L.L.; Mok, J.; Lee, M.J.; et al. The versatile functions of Sox9 in development, stem cells, and human diseases. Genes Dis. 2014, 1, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhu, X.; Xie, J.; Zhao, Y.; Liu, X. beta-amyloid increases neurocan expression through regulating Sox9 in astrocytes: A potential relationship between Sox9 and chondroitin sulfate proteoglycans in Alzheimer’s disease. Brain Res. 2016, 1646, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Gonen, N.; Lovell-Badge, R. The regulation of Sox9 expression in the gonad. Curr. Top. Dev. Biol. 2019, 134, 223–252. [Google Scholar] [PubMed]
- Nicholls, R.E.; Sontag, J.M.; Zhang, H.; Staniszewski, A.; Yan, S.; Kim, C.Y.; Yim, M.; Woodruff, C.M.; Arning, E.; Wasek, B.; et al. PP2A methylation controls sensitivity and resistance to beta-amyloid-induced cognitive and electrophysiological impairments. Proc. Natl. Acad. Sci. USA 2016, 113, 3347–3352. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, A.; Wani, W.Y.; Bal, A.; Gill, K.D.; Kaur, J. Okadaic Acid and Hypoxia Induced Dementia Model of Alzheimer’s Type in Rats. Neurotox. Res. 2019, 35, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, T.; Matta, C.; Somogyi, C.; Katona, E.; Takacs, R.; Soha, R.F.; Szabo, I.A.; Cserhati, C.; Szody, R.; Karacsonyi, Z.; et al. Mechanical loading stimulates chondrogenesis via the PKA/CREB-Sox9 and PP2A pathways in chicken micromass cultures. Cell Signal. 2014, 26, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Zakany, R.; Szijgyarto, Z.; Matta, C.; Juhasz, T.; Csortos, C.; Szucs, K.; Czifra, G.; Biro, T.; Modis, L.; Gergely, P. Hydrogen peroxide inhibits formation of cartilage in chicken micromass cultures and decreases the activity of calcineurin: Implication of ERK1/2 and Sox9 pathways. Exp. Cell Res. 2005, 305, 190–199. [Google Scholar] [CrossRef]
- McKenzie-Nickson, S.; Chan, J.; Perez, K.; Hung, L.W.; Cheng, L.; Sedjahtera, A.; Gunawan, L.; Adlard, P.A.; Hayne, D.J.; McInnes, L.E.; et al. Modulating protein phosphatase 2A rescues disease phenotype in neurodegenerative tauopathies. ACS Chem. Neurosci. 2018, 9, 2731–2740. [Google Scholar] [CrossRef]
- Hopp, S.C.; Bihlmeyer, N.A.; Corradi, J.P.; Vanderburg, C.; Cacace, A.M.; Das, S.; Clark, T.W.; Betensky, R.A.; Hyman, B.T.; Hudry, E. Neuronal calcineurin transcriptional targets parallel changes observed in Alzheimer disease brain. J. Neurochem. 2018, 147, 24–39. [Google Scholar] [CrossRef]
- Georg, I.; Barrionuevo, F.; Wiech, T.; Scherer, G. Sox9 and Sox8 are required for basal lamina integrity of testis cords and for suppression of FOXL2 during embryonic testis development in mice. Biol. Reprod. 2012, 87, 99. [Google Scholar] [CrossRef]
- Chen, H.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Regulation of spermatogenesis by a local functional axis in the testis: Role of the basement membrane-derived noncollagenous 1 domain peptide. FASEB J. 2017, 31, 3587–3607. [Google Scholar] [CrossRef] [PubMed]
- Christov, A.; Ottman, J.; Hamdheydari, L.; Grammas, P. Structural changes in Alzheimer’s disease brain microvessels. Curr. Alzheimer Res. 2008, 5, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, Y.; Isobe, Y.; Fukushima, K. Type IV collagen prevents amyloid beta-protein fibril formation. Life Sci. 2002, 70, 1555–1564. [Google Scholar] [CrossRef]
- Levit, A.; Hachinski, V.; Whitehead, S.N. Neurovascular unit dysregulation, white matter disease, and executive dysfunction: The shared triad of vascular cognitive impairment and Alzheimer disease. Geroscience 2020, 42, 445–465. [Google Scholar] [CrossRef] [PubMed]
- Schmid, N.; Missel, A.; Petkov, S.; Stockl, J.B.; Flenkenthaler, F.; Arnold, G.J.; Frohlich, T.; Behr, R.; Mayerhofer, A. A translational cellular model for the study of peritubular cells of the testis. Reproduction 2020, 160, 259–268. [Google Scholar] [CrossRef]
- Samy, A.; El-Adl, M.; Rezk, S.; Marghani, B.; Eldomany, W.; Eldesoky, A.; Elmetwally, M.A. The potential protective and therapeutic effects of platelet-rich plasma on ischemia/reperfusion injury following experimental torsion/detorsion of testis in the Albino rat model. Life Sci. 2020, 256, 117982. [Google Scholar] [CrossRef]
- Amann, R.P. The cycle of the seminiferous epithelium in humans: A need to revisit? J. Androl. 2008, 29, 469–487. [Google Scholar] [CrossRef]
- Li, M.; Arimura, A. Neuropeptides of the pituitary adenylate cyclase-activating polypeptide/vasoactive intestinal polypeptide/growth hormone-releasing hormone/secretin family in testis. Endocrine 2003, 20, 201–214. [Google Scholar] [CrossRef]
- Ivic, I.; Fulop, B.D.; Juhasz, T.; Reglodi, D.; Toth, G.; Hashimoto, H.; Tamas, A.; Koller, A. Backup mechanisms maintain PACAP/VIP-induced arterial relaxations in pituitary adenylate cyclase-activating polypeptide-deficient mice. J. Vasc. Res. 2017, 54, 180–192. [Google Scholar] [CrossRef]
- Salmina, A.B.; Komleva, Y.K.; Lopatina, O.L.; Birbrair, A. Pericytes in Alzheimer’s Disease: Novel clues to cerebral amyloid angiopathy pathogenesis. Adv. Exp. Med. Biol. 2019, 1147, 147–166. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Nucleotide Sequence (5′→3′) | GenBank ID | Annealing Temperature | Amplimer Size (bp) |
|---|---|---|---|---|---|
| PAC1 | sense | TATTACTACCTGTCGGTGAAG (912–932) | NM_007407.4 | 52 °C | 213 |
| antisense | ATGACTGCTGTCCTGCTC (1107–1124) | ||||
| VPAC1 | sense | TTT GAG GAT TTC GGG TGC (974–991) | NM_011703.4 | 53 °C | 266 |
| antisense | TGG GCC TTA AAG TTG TCG (1222–1239) | ||||
| VPAC2 | sense | CTC CTG GTA GCC ATC CTT (805–822) | NM_009511.2 | 53 °C | 149 |
| antisense | ATG CTG TGG TCG TTT GTG (936–953) | ||||
| PKA (Prkaca) | sense | GCAAAGGCTACAACAAGGC (847–865) | NM_008854 | 53 °C | 280 |
| antisense | ATGGCAATCCAGTCAATCG (1109–1126) | ||||
| Sox9 | sense | GTA CCC GCA TCT GCA CAA CG (378–397) | NM_011448 | 62 °C | 521 |
| antisense | GTG GCA AGT ATT GGT CAA ACT CAT T (874–898) | ||||
| Sox10 | sense | ACG ACT GGA CGC TGG TGC (535–552) | NM_011437.1 | 58 °C | 283 |
| antisense | CGC CGA GGT TGG TAC TTG TAG (797–817) | ||||
| PP2A (Ppp2ca) | sense | CTC TGC GAG AAG GCT AAA (288–305) | NM_017039.2 | 54 °C | 436 |
| antisense | TGA TTC CCT CGG AGT ATG (706–723) | ||||
| collagen IV (Col4a1) | sense | TCG GCT ATT CCT TCG TGA TG (4963–4982) | NM_009931.2 | 52 °C | 209 |
| antisense | GGATGGCGTGGGCTTCTT (5154–5171) | ||||
| collagen IX (Col9a3) | sense | CAG GTT CCG ATG GTC TTC C (1357–1375) | NM_009936.2 | 55 °C | 492 |
| antisense | CTG TTG CTC CCT TGT CCC (1831–1848) | ||||
| testatin (Cys9) | sense | CTG GAG GGA GAA GGT AAA (274–291) | NM_009979.1 | 51 °C | 142 |
| antisense | CAG GCA GGT GAA GGT ATT (398–415) | ||||
| actin (Actb) | sense | GCCAACCGTGAAAAGATGA (419–437) | NM_007393.5 | 54 °C | 462 |
| antisense | CAAGAAGGAAGGCTGGAAAA (861–880) |
| Antibody | Host Animal | Dilution | Distributor |
|---|---|---|---|
| Anti-Col. IX. | rabbit, polyclonal, | 1:500 | Abcam, Cambridge, UK |
| Anti-Col. IV. | rabbit, polyclonal, | 1:800 | Abcam, Cambridge, UK |
| Anti-Testatin | rabbit, polyclonal, | 1:300 | Santa Cruz Biotechnology, Dallas, TX, USA |
| Anti-PKA C | rabbit, polyclonal, | 1:600 | Cell Signaling, Danvers, MA, USA |
| Anti-Sox9 | rabbit, polyclonal, | 1:600 | Abcam, Cambridge, UK |
| Anti-P-Sox9 | rabbit, polyclonal, | 1:800 | Sigma-Aldrich, St. Louis, MO, USA |
| Anti-Sox10 | rabbit, polyclonal, | 1:500 | Abcam, Cambridge, UK |
| Anti-PP2A C | rabbit, polyclonal, | 1:600 | Cell Signaling, Danvers, MA, USA |
| Anti-Actin | mouse, monoclonal, | 1:10,000 | Sigma-Aldrich, St. Louis, MO, USA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szegeczki, V.; Horváth, G.; Perényi, H.; Tamás, A.; Radák, Z.; Ábrahám, D.; Zákány, R.; Reglodi, D.; Juhász, T. Alzheimer’s Disease Mouse as a Model of Testis Degeneration. Int. J. Mol. Sci. 2020, 21, 5726. https://doi.org/10.3390/ijms21165726
Szegeczki V, Horváth G, Perényi H, Tamás A, Radák Z, Ábrahám D, Zákány R, Reglodi D, Juhász T. Alzheimer’s Disease Mouse as a Model of Testis Degeneration. International Journal of Molecular Sciences. 2020; 21(16):5726. https://doi.org/10.3390/ijms21165726
Chicago/Turabian StyleSzegeczki, Vince, Gabriella Horváth, Helga Perényi, Andrea Tamás, Zsolt Radák, Dóra Ábrahám, Róza Zákány, Dora Reglodi, and Tamás Juhász. 2020. "Alzheimer’s Disease Mouse as a Model of Testis Degeneration" International Journal of Molecular Sciences 21, no. 16: 5726. https://doi.org/10.3390/ijms21165726
APA StyleSzegeczki, V., Horváth, G., Perényi, H., Tamás, A., Radák, Z., Ábrahám, D., Zákány, R., Reglodi, D., & Juhász, T. (2020). Alzheimer’s Disease Mouse as a Model of Testis Degeneration. International Journal of Molecular Sciences, 21(16), 5726. https://doi.org/10.3390/ijms21165726