Bone Microenvironment and Osteosarcoma Metastasis

 , , ,
, , ,
Abstract

1. Introduction
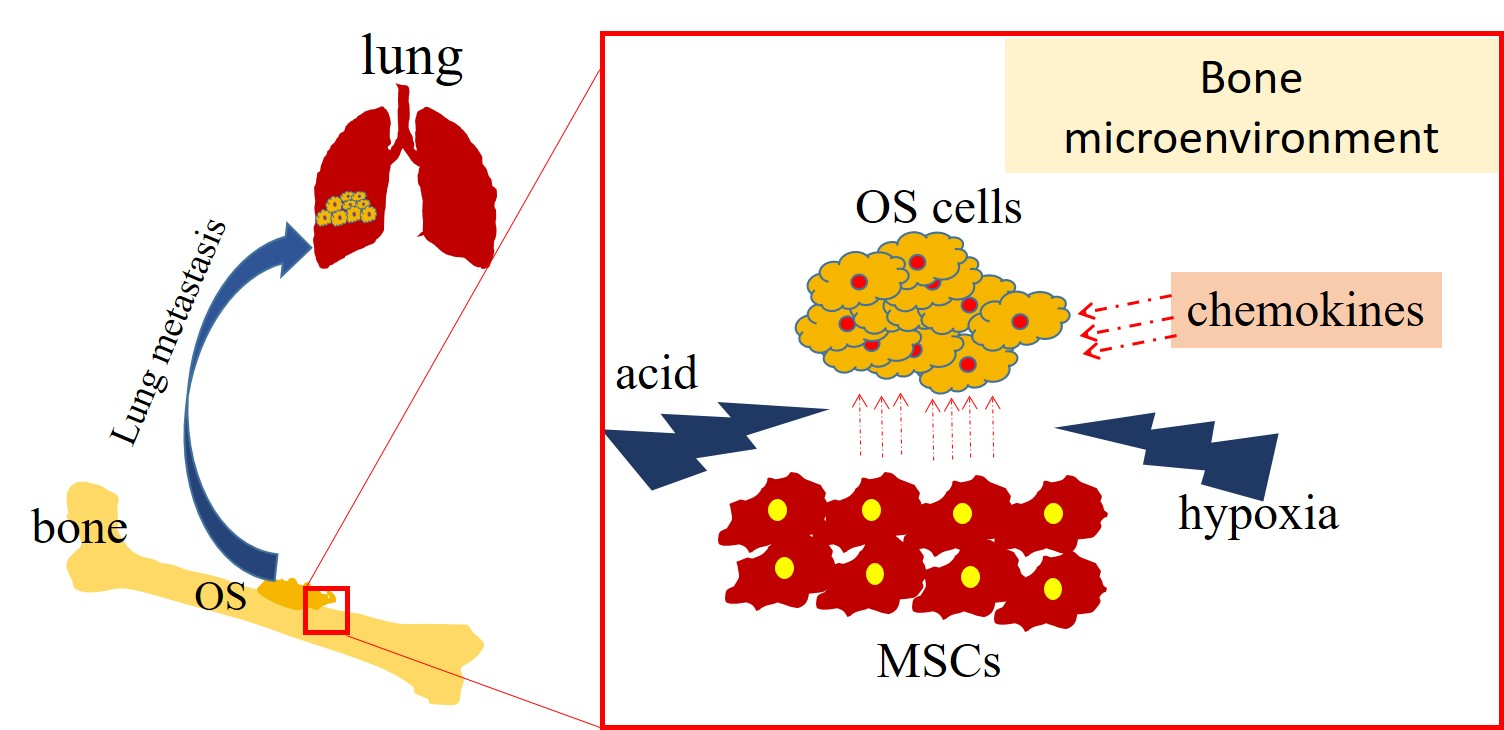
2. Bone Microenvironment and OS Metastasis
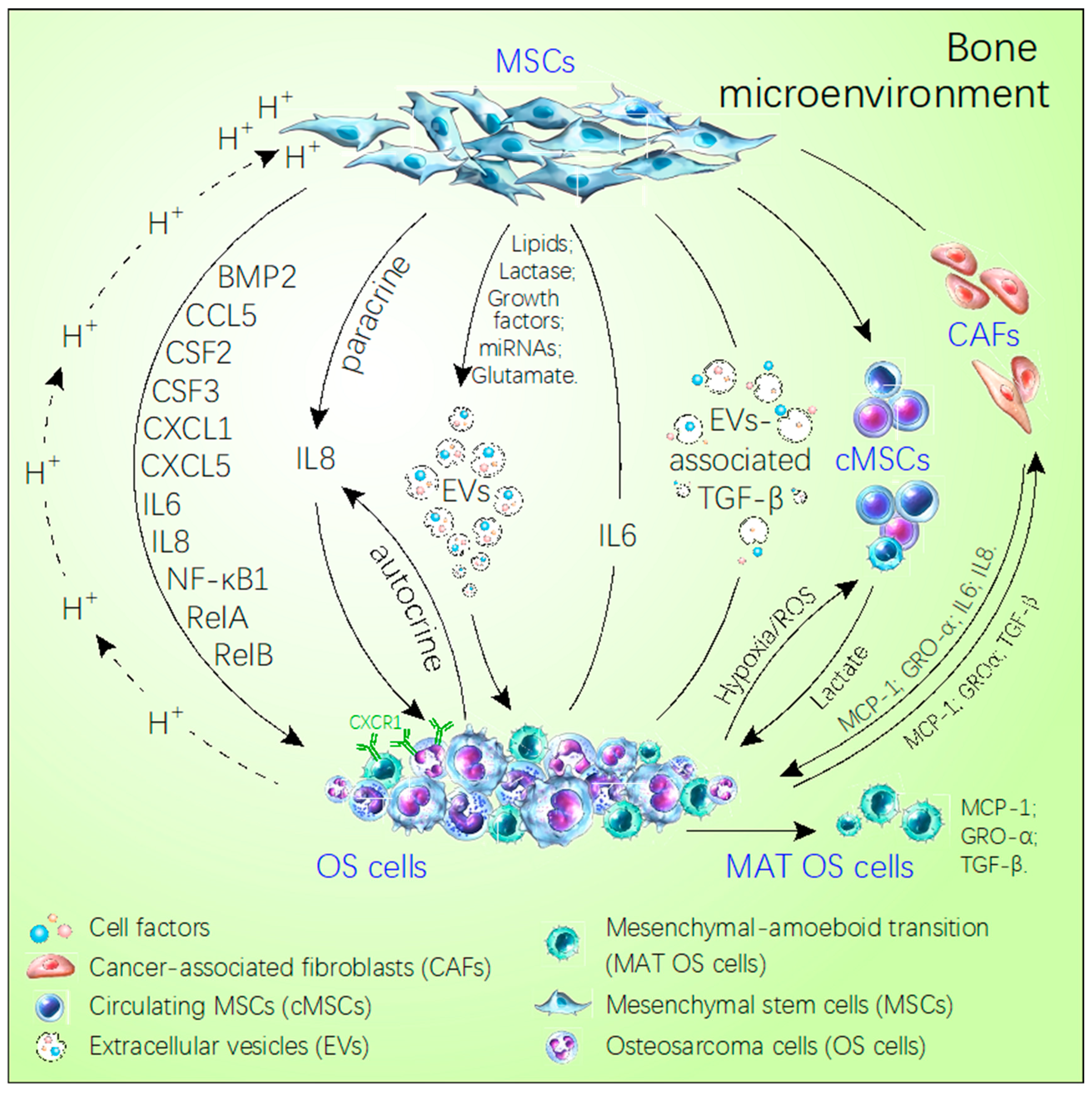
2.1. Mesenchymal Stem Cells and OS Metastasis
2.2. Effect of Hypoxia and Acidosis Environment on OS Metastasis
2.3. Chemokines and OS Metastasis
2.4. Functions of Extracellular Vesicles in the Tumor Microenvironment
3. Signal Pathways in OS Metastasis
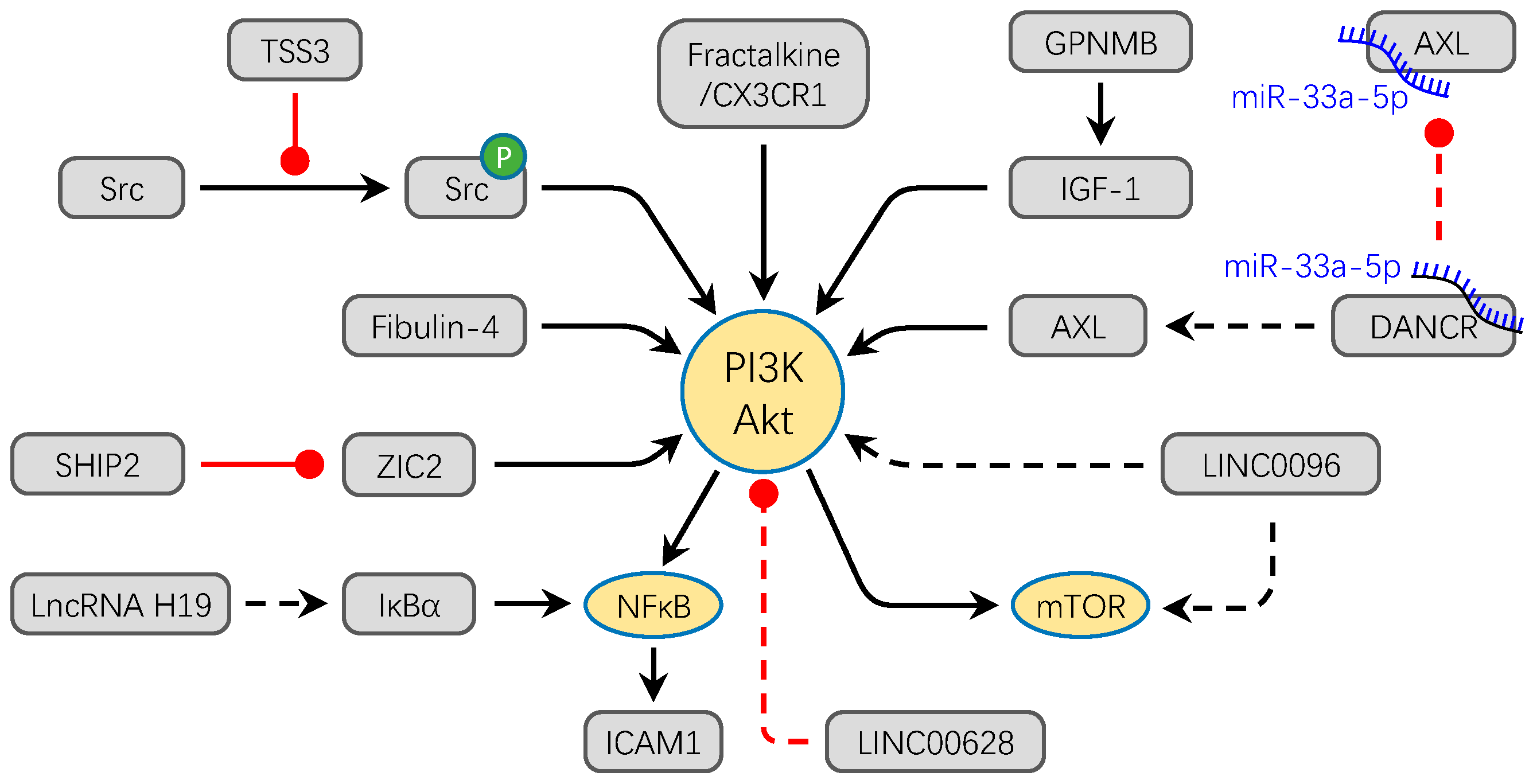
3.1. PI3K/Akt Signaling Pathway
3.2. Wnt/β-Catenin Signaling Pathway
3.3. MAPK/ERK Signaling Pathway
3.4. Hedgehog Signaling Pathway
3.5. Notch Signaling Pathway
4. The Bone Microenvironment as a Promising Treatment
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ANGPTL4 | Angiopoietin-like 4 |
| AXL | Tyrosine-protein kinase receptor UFO |
| CK1α | Casein kinase I α |
| CRNDE | Colorectal Neoplasia Differentially Expressed |
| DANCR | Differentiation Antagonizing Non-Protein Coding RNA |
| DTX1 | Protein deltex-1 |
| DUSP1 | Dual specificity protein phosphatase 1 |
| ECM | Extracellular matrices |
| GPNMB | Glycoprotein non-metastatic melanoma protein B |
| HIF2PUT | Hypoxia-inducible factor-2α promoter upstream transcript |
| MALAT1 | Metastasis associated lung adenocarcinoma transcript 1 |
| TWIST | twist family bHLH transcription factor 1 |
| MCP-1 | Monocyte chemoattractant protein 1 |
| ONZIN | placenta specific 8 |
| PTK2 | Protein tyrosine kinase 2 |
| SENP1 | Sentrin-specific protease 1 |
| SNHG12 | small nucleolar RNA host gene 12 |
| TET3 | Tet Methylcytosine Dioxygenase 3 |
| TSSC3 | tumor- suppressing STF cDNA 3 |
| ZIC2 | Zinc finger protein 2 |
| β-TrcP | β-transducin repeats-containing proteins |
References
- Zheng, Y.; Zhou, H.; Dunstan, C.R.; Sutherland, R.L.; Seibel, M.J. The role of the bone microenvironment in skeletal metastasis. J. Bone Oncol. 2013, 2, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Junttila, M.R.; de Sauvage, F.J. Influence of tumour micro-environment heterogeneity on therapeutic response. Nature 2013, 501, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Belli, C.; Trapani, D.; Viale, G.; D’Amico, P.; Duso, B.A.; Della Vigna, P.; Orsi, F.; Curigliano, G. Targeting the microenvironment in solid tumors. Cancer Treat. Rev. 2018, 65, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Buenrostro, D.; Mulcrone, P.L.; Owens, P.; Sterling, J.A. The Bone Microenvironment: A Fertile Soil for Tumor Growth. Curr. Osteoporos. Rep. 2016, 14, 151–158. [Google Scholar] [CrossRef]
- Hui, L.; Chen, Y. Tumor microenvironment: Sanctuary of the devil. Cancer Lett. 2015, 368, 7–13. [Google Scholar] [CrossRef]
- Seton-Rogers, S. Mapping dysfunction of the bone marrow stroma in leukaemia. Nat. Rev. Cancer 2019, 19, 368. [Google Scholar] [CrossRef]
- Yoneda, T.; Sasaki, A.; Mundy, G.R. Osteolytic bone metastasis in breast cancer. Breast Cancer Res. Treat. 1994, 32, 73–84. [Google Scholar] [CrossRef]
- Alfranca, A.; Martinez-Cruzado, L.; Tornin, J.; Abarrategi, A.; Amaral, T.; de Alava, E.; Menendez, P.; Garcia-Castro, J.; Rodriguez, R. Bone microenvironment signals in osteosarcoma development. Cell. Mol. Life Sci. 2015, 72, 3097–3113. [Google Scholar] [CrossRef]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun. Signal. 2020, 18, 59. [Google Scholar] [CrossRef]
- Le Nail, L.R.; Brennan, M.; Rosset, P.; Deschaseaux, F.; Piloquet, P.; Pichon, O.; Le Caignec, C.; Crenn, V.; Layrolle, P.; Herault, O.; et al. Comparison of Tumor- and Bone Marrow-Derived Mesenchymal Stromal/Stem Cells from Patients with High-Grade Osteosarcoma. Int. J. Mol. Sci. 2018, 19, 707. [Google Scholar] [CrossRef]
- Cortini, M.; Avnet, S.; Baldini, N. Mesenchymal stroma: Role in osteosarcoma progression. Cancer Lett. 2017, 405, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Botter, S.M.; Neri, D.; Fuchs, B. Recent advances in osteosarcoma. Curr. Opin. Pharmacol. 2014, 16, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Khoury, J.D.; Hoffer, F.A.; Wu, J.; Billups, C.A.; Heck, R.K.; Quintana, J.; Poe, D.; Rao, B.N.; Daw, N.C. Telangiectatic osteosarcoma: The St. Jude Children’s Research Hospital’s experience. Cancer 2007, 109, 1627–1637. [Google Scholar] [CrossRef] [PubMed]
- Ragland, B.D.; Bell, W.C.; Lopez, R.R.; Siegal, G.P. Cytogenetics and molecular biology of osteosarcoma. Lab. Invest. 2002, 82, 365–373. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Walkley, C.R.; Qudsi, R.; Sankaran, V.G.; Perry, J.A.; Gostissa, M.; Roth, S.I.; Rodda, S.J.; Snay, E.; Dunning, P.; Fahey, F.H.; et al. Conditional mouse osteosarcoma, dependent on p53 loss and potentiated by loss of Rb, mimics the human disease. Genes Dev. 2008, 22, 1662–1676. [Google Scholar] [CrossRef]
- Mutsaers, A.J.; Ng, A.J.; Baker, E.K.; Russell, M.R.; Chalk, A.M.; Wall, M.; Liddicoat, B.J.; Ho, P.W.; Slavin, J.L.; Goradia, A.; et al. Modeling distinct osteosarcoma subtypes in vivo using Cre:lox and lineage-restricted transgenic shRNA. Bone 2013, 55, 166–178. [Google Scholar] [CrossRef]
- Berman, S.D.; Calo, E.; Landman, A.S.; Danielian, P.S.; Miller, E.S.; West, J.C.; Fonhoue, B.D.; Caron, A.; Bronson, R.; Bouxsein, M.L.; et al. Metastatic osteosarcoma induced by inactivation of Rb and p53 in the osteoblast lineage. Proc. Natl. Acad. Sci. USA 2008, 105, 11851–11856. [Google Scholar] [CrossRef]
- Rubio, R.; Abarrategi, A.; Garcia-Castro, J.; Martinez-Cruzado, L.; Suarez, C.; Tornin, J.; Santos, L.; Astudillo, A.; Colmenero, I.; Mulero, F.; et al. Bone environment is essential for osteosarcoma development from transformed mesenchymal stem cells. Stem Cells 2014, 32, 1136–1148. [Google Scholar] [CrossRef]
- He, J.P.; Hao, Y.; Wang, X.L.; Yang, X.J.; Shao, J.F.; Guo, F.J.; Feng, J.X. Review of the molecular pathogenesis of osteosarcoma. Asian Pac. J. Cancer Prev. 2014, 15, 5967–5976. [Google Scholar] [CrossRef]
- Broadhead, M.L.; Clark, J.C.; Myers, D.E.; Dass, C.R.; Choong, P.F. The molecular pathogenesis of osteosarcoma: A review. Sarcoma 2011, 2011, 959248. [Google Scholar] [CrossRef]
- Denduluri, S.K.; Wang, Z.; Yan, Z.; Wang, J.; Wei, Q.; Mohammed, M.K.; Haydon, R.C.; Luu, H.H.; He, T.C. Molecular pathogenesis and therapeutic strategies of human osteosarcoma. J. Biomed. Res. 2015, 30. [Google Scholar] [CrossRef]
- Kansara, M.; Thomas, D.M. Molecular pathogenesis of osteosarcoma. DNA Cell Biol. 2007, 26, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bielack, S.S.; Kempf-Bielack, B.; Delling, G.; Exner, G.U.; Flege, S.; Helmke, K.; Kotz, R.; Salzer-Kuntschik, M.; Werner, M.; Winkelmann, W.; et al. Prognostic factors in high-grade osteosarcoma of the extremities or trunk: An analysis of 1,702 patients treated on neoadjuvant cooperative osteosarcoma study group protocols. J. Clin. Oncol. 2002, 20, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Mirabello, L.; Troisi, R.J.; Savage, S.A. Osteosarcoma incidence and survival rates from 1973 to 2004: Data from the Surveillance, Epidemiology, and End Results Program. Cancer 2009, 115, 1531–1543. [Google Scholar] [CrossRef] [PubMed]
- Ehnman, M.; Chaabane, W.; Haglund, F.; Tsagkozis, P. The Tumor Microenvironment of Pediatric Sarcoma: Mesenchymal Mechanisms Regulating Cell Migration and Metastasis. Curr. Oncol. Rep. 2019, 21, 90. [Google Scholar] [CrossRef]
- Mohseny, A.B.; Szuhai, K.; Romeo, S.; Buddingh, E.P.; Briaire-de Bruijn, I.; de Jong, D.; van Pel, M.; Cleton-Jansen, A.M.; Hogendoorn, P.C. Osteosarcoma originates from mesenchymal stem cells in consequence of aneuploidization and genomic loss of Cdkn2. J. Pathol. 2009, 219, 294–305. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Honoki, K.; Fujii, H.; Tohma, Y.; Kido, A.; Mori, T.; Tsujiuchi, T.; Tanaka, Y. Mesenchymal stem cells promote tumor engraftment and metastatic colonization in rat osteosarcoma model. Int. J. Oncol. 2012, 40, 163–169. [Google Scholar]
- Zhang, M.; Zhang, X. Association of MMP-2 expression and prognosis in osteosarcoma patients. Int. J. Clin. Exp. Pathol. 2015, 8, 14965–14970. [Google Scholar]
- Neklyudova, O.; Arlt, M.J.; Brennecke, P.; Thelen, M.; Gvozdenovic, A.; Kuzmanov, A.; Robl, B.; Botter, S.M.; Born, W.; Fuchs, B. Altered CXCL12 expression reveals a dual role of CXCR4 in osteosarcoma primary tumor growth and metastasis. J. Cancer Res. Clin. Oncol. 2016, 142, 1739–1750. [Google Scholar] [CrossRef]
- Bonuccelli, G.; Avnet, S.; Grisendi, G.; Salerno, M.; Granchi, D.; Dominici, M.; Kusuzaki, K.; Baldini, N. Role of mesenchymal stem cells in osteosarcoma and metabolic reprogramming of tumor cells. Oncotarget 2014, 5, 7575–7588. [Google Scholar] [CrossRef]
- Avnet, S.; Di Pompo, G.; Chano, T.; Errani, C.; Ibrahim-Hashim, A.; Gillies, R.J.; Donati, D.M.; Baldini, N. Cancer-associated mesenchymal stroma fosters the stemness of osteosarcoma cells in response to intratumoral acidosis via NF-κB activation. Int. J. Cancer 2017, 140, 1331–1345. [Google Scholar] [CrossRef] [PubMed]
- Pietrovito, L.; Leo, A.; Gori, V.; Lulli, M.; Parri, M.; Becherucci, V.; Piccini, L.; Bambi, F.; Taddei, M.L.; Chiarugi, P. Bone marrow-derived mesenchymal stem cells promote invasiveness and transendothelial migration of osteosarcoma cells via a mesenchymal to amoeboid transition. Mol. Oncol. 2018, 12, 659–676. [Google Scholar] [CrossRef] [PubMed]
- Baglio, S.R.; Lagerweij, T.; Perez-Lanzon, M.; Ho, X.D.; Leveille, N.; Melo, S.A.; Cleton-Jansen, A.M.; Jordanova, E.S.; Roncuzzi, L.; Greco, M.; et al. Blocking Tumor-Educated MSC Paracrine Activity Halts Osteosarcoma Progression. Clin. Cancer Res. 2017, 23, 3721–3733. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, X.; Miao, W.; Wang, B.; Qiu, Y. CXCL8 promotes the invasion of human osteosarcoma cells by regulation of PI3K/Akt signaling pathway. APMIS 2017, 125, 773–780. [Google Scholar] [CrossRef]
- Liu, M.; Wang, D.; Li, N. MicroRNA-20b Downregulates HIF-1α and Inhibits the Proliferation and Invasion of Osteosarcoma Cells. Oncol. Res. 2016, 23, 257–266. [Google Scholar] [CrossRef]
- Vallabhaneni, K.C.; Hassler, M.Y.; Abraham, A.; Whitt, J.; Mo, Y.Y.; Atfi, A.; Pochampally, R. Mesenchymal Stem/Stromal Cells under Stress Increase Osteosarcoma Migration and Apoptosis Resistance via Extracellular Vesicle Mediated Communication. PLoS ONE 2016, 11, e0166027. [Google Scholar] [CrossRef]
- Geng, S.; Zhang, X.; Chen, J.; Liu, X.; Zhang, H.; Xu, X.; Ma, Y.; Li, B.; Zhang, Y.; Bi, Z.; et al. The tumor suppressor role of miR-124 in osteosarcoma. PLoS ONE 2014, 9, e91566. [Google Scholar] [CrossRef]
- Lagerweij, T.; Perez-Lanzon, M.; Baglio, S.R. A Preclinical Mouse Model of Osteosarcoma to Define the Extracellular Vesicle-mediated Communication Between Tumor and Mesenchymal Stem Cells. J. Vis. Exp. 2018, e56932. [Google Scholar] [CrossRef]
- Du, L.; Han, X.G.; Tu, B.; Wang, M.Q.; Qiao, H.; Zhang, S.H.; Fan, Q.M.; Tang, T.T. CXCR1/Akt signaling activation induced by mesenchymal stem cell-derived IL-8 promotes osteosarcoma cell anoikis resistance and pulmonary metastasis. Cell Death Dis. 2018, 9, 714. [Google Scholar] [CrossRef]
- McMahon, S.; Charbonneau, M.; Grandmont, S.; Richard, D.E.; Dubois, C.M. Transforming growth factor beta1 induces hypoxia-inducible factor-1 stabilization through selective inhibition of PHD2 expression. J. Biol. Chem. 2006, 281, 24171–24181. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, C.; Wang, K.; Liu, X.; Liu, Q. MicroRNA-33b Inhibits the Proliferation and Migration of Osteosarcoma Cells via Targeting Hypoxia-Inducible Factor-1α. Oncol. Res. 2017, 25, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ren, T.; Huang, Y.; Bao, X.; Sun, K.; Shen, D.; Guo, W. BMPR2 and HIF1-α overexpression in resected osteosarcoma correlates with distant metastasis and patient survival. Chin. J. Cancer Res. 2017, 29, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.C.; Tang, C.; Dong, Y.; Zhang, J.; Yuan, T.; Tao, S.C.; Li, X.L. Targeting the long noncoding RNA MALAT1 blocks the pro-angiogenic effects of osteosarcoma and suppresses tumour growth. Int. J. Biol. Sci. 2017, 13, 1398–1408. [Google Scholar] [CrossRef]
- Zhang, T.; Kastrenopoulou, A.; Larrouture, Q.; Athanasou, N.A.; Knowles, H.J. Angiopoietin-like 4 promotes osteosarcoma cell proliferation and migration and stimulates osteoclastogenesis. BMC Cancer 2018, 18, 536. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; He, X.; Xue, R.; Zhang, Y.; Zhang, X.; Lu, J.; Zhang, Z.; Xue, L. Combined over-expression of the hypoxia-inducible factor 2α gene and its long non-coding RNA predicts unfavorable prognosis of patients with osteosarcoma. Pathol. Res. Pract. 2016, 212, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Odagiri, H.; Kadomatsu, T.; Endo, M.; Masuda, T.; Morioka, M.S.; Fukuhara, S.; Miyamoto, T.; Kobayashi, E.; Miyata, K.; Aoi, J.; et al. The secreted protein ANGPTL2 promotes metastasis of osteosarcoma cells through integrin α5β1, p38 MAPK, and matrix metalloproteinases. Sci. Signal. 2014, 7, ra7. [Google Scholar] [CrossRef]
- Wang, X.; Liang, X.; Liang, H.; Wang, B. SENP1/HIF-1α feedback loop modulates hypoxia-induced cell proliferation, invasion, and EMT in human osteosarcoma cells. J. Cell. Biochem. 2018, 119, 1819–1826. [Google Scholar] [CrossRef]
- Charo, I.F.; Ransohoff, R.M. The many roles of chemokines and chemokine receptors in inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef]
- Zlotnik, A.; Burkhardt, A.M.; Homey, B. Homeostatic chemokine receptors and organ-specific metastasis. Nat. Rev. Immunol. 2011, 11, 597–606. [Google Scholar] [CrossRef]
- Li, Y.; Flores, R.; Yu, A.; Okcu, M.F.; Murray, J.; Chintagumpala, M.; Hicks, J.; Lau, C.C.; Man, T.K. Elevated expression of CXC chemokines in pediatric osteosarcoma patients. Cancer 2011, 117, 207–217. [Google Scholar] [CrossRef]
- Pradelli, E.; Karimdjee-Soilihi, B.; Michiels, J.F.; Ricci, J.E.; Millet, M.A.; Vandenbos, F.; Sullivan, T.J.; Collins, T.L.; Johnson, M.G.; Medina, J.C.; et al. Antagonism of chemokine receptor CXCR3 inhibits osteosarcoma metastasis to lungs. Int. J. Cancer 2009, 125, 2586–2594. [Google Scholar] [CrossRef]
- Ren, Z.; Liang, S.; Yang, J.; Han, X.; Shan, L.; Wang, B.; Mu, T.; Zhang, Y.; Yang, X.; Xiong, S.; et al. Coexpression of CXCR4 and MMP9 predicts lung metastasis and poor prognosis in resected osteosarcoma. Tumour Biol. 2016, 37, 5089–5096. [Google Scholar] [CrossRef]
- Han, Y.; Wu, C.; Wang, J.; Liu, N. CXCR7 maintains osteosarcoma invasion after CXCR4 suppression in bone marrow microenvironment. Tumour Biol. 2017, 39, 1010428317701631. [Google Scholar] [CrossRef]
- Wang, S.W.; Liu, S.C.; Sun, H.L.; Huang, T.Y.; Chan, C.H.; Yang, C.Y.; Yeh, H.I.; Huang, Y.L.; Chou, W.Y.; Lin, Y.M.; et al. CCL5/CCR5 axis induces vascular endothelial growth factor-mediated tumor angiogenesis in human osteosarcoma microenvironment. Carcinogenesis 2015, 36, 104–114. [Google Scholar] [CrossRef]
- Kushlinskii, N.E.; Timofeev, Y.S.; Solov’ev, Y.N.; Gerstein, E.S.; Lyubimova, N.V.; Bulycheva, I.V. Components of the RANK/RANKL/OPG system, IL-6, IL-8, IL-16, MMP-2, and calcitonin in the sera of patients with bone tumors. Bull. Exp. Biol. Med. 2014, 157, 520–523. [Google Scholar] [CrossRef]
- Jerez, S.; Araya, H.; Hevia, D.; Irarrazaval, C.E.; Thaler, R.; van Wijnen, A.J.; Galindo, M. Extracellular vesicles from osteosarcoma cell lines contain miRNAs associated with cell adhesion and apoptosis. Gene 2019, 710, 246–257. [Google Scholar] [CrossRef]
- Lan, M.; Zhu, X.P.; Cao, Z.Y.; Liu, J.M.; Lin, Q.; Liu, Z.L. Extracellular vesicles-mediated signaling in the osteosarcoma microenvironment: Roles and potential therapeutic targets. J. Bone Oncol. 2018, 12, 101–104. [Google Scholar] [CrossRef]
- Qi, J.; Zhou, Y.; Jiao, Z.; Wang, X.; Zhao, Y.; Li, Y.; Chen, H.; Yang, L.; Zhu, H.; Li, Y. Exosomes Derived from Human Bone Marrow Mesenchymal Stem Cells Promote Tumor Growth Through Hedgehog Signaling Pathway. Cell Physiol. Biochem. 2017, 42, 2242–2254. [Google Scholar] [CrossRef]
- Lin, S.; Zhu, B.; Huang, G.; Zeng, Q.; Wang, C. Microvesicles derived from human bone marrow mesenchymal stem cells promote U2OS cell growth under hypoxia: The role of PI3K/AKT and HIF-1α. Hum. Cell 2019, 32, 64–74. [Google Scholar] [CrossRef]
- Mannerstrom, B.; Kornilov, R.; Abu-Shahba, A.G.; Chowdhury, I.M.; Sinha, S.; Seppanen-Kaijansinkko, R.; Kaur, S. Epigenetic alterations in mesenchymal stem cells by osteosarcoma-derived extracellular vesicles. Epigenetics 2019, 14, 352–364. [Google Scholar] [CrossRef]
- Garimella, R.; Washington, L.; Isaacson, J.; Vallejo, J.; Spence, M.; Tawfik, O.; Rowe, P.; Brotto, M.; Perez, R. Extracellular Membrane Vesicles Derived from 143B Osteosarcoma Cells Contain Pro-Osteoclastogenic Cargo: A Novel Communication Mechanism in Osteosarcoma Bone Microenvironment. Transl. Oncol. 2014, 7, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Macklin, R.; Wang, H.; Loo, D.; Martin, S.; Cumming, A.; Cai, N.; Lane, R.; Ponce, N.S.; Topkas, E.; Inder, K.; et al. Extracellular vesicles secreted by highly metastatic clonal variants of osteosarcoma preferentially localize to the lungs and induce metastatic behaviour in poorly metastatic clones. Oncotarget 2016, 7, 43570–43587. [Google Scholar] [CrossRef] [PubMed]
- Perut, F.; Roncuzzi, L.; Baldini, N. The Emerging Roles of Extracellular Vesicles in Osteosarcoma. Front. Oncol. 2019, 9, 1342. [Google Scholar] [CrossRef] [PubMed]
- Engelman, J.A. Targeting PI3K signalling in cancer: Opportunities, challenges and limitations. Nat. Rev. Cancer 2009, 9, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, X.H.; Yan, Y.G.; Wang, C.; Wang, W.J. PI3K/Akt signaling in osteosarcoma. Clin. Chim. Acta 2015, 444, 182–192. [Google Scholar] [CrossRef]
- Zhao, G.; Cai, C.; Yang, T.; Qiu, X.; Liao, B.; Li, W.; Ji, Z.; Zhao, J.; Zhao, H.; Guo, M.; et al. MicroRNA-221 induces cell survival and cisplatin resistance through PI3K/Akt pathway in human osteosarcoma. PLoS ONE 2013, 8, e53906. [Google Scholar] [CrossRef]
- Liu, J.F.; Tsao, Y.T.; Hou, C.H. Fractalkine/CX3CL1 induced intercellular adhesion molecule-1-dependent tumor metastasis through the CX3CR1/PI3K/Akt/NF-κB pathway in human osteosarcoma. Oncotarget 2017, 8, 54136–54148. [Google Scholar] [CrossRef]
- Zhao, G.S.; Gao, Z.R.; Zhang, Q.; Tang, X.F.; Lv, Y.F.; Zhang, Z.S.; Zhang, Y.; Tan, Q.L.; Peng, D.B.; Jiang, D.M.; et al. TSSC3 promotes autophagy via inactivating the Src-mediated PI3K/Akt/mTOR pathway to suppress tumorigenesis and metastasis in osteosarcoma, and predicts a favorable prognosis. J. Exp. Clin. Cancer Res. 2018, 37, 188. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, S.; Chen, J.; Liu, H.; Lu, J.; Jiang, H.; Huang, A.; Chen, Y. Fibulin-4 promotes osteosarcoma invasion and metastasis by inducing epithelial to mesenchymal transition via the PI3K/Akt/mTOR pathway. Int. J. Oncol. 2017, 50, 1513–1530. [Google Scholar] [CrossRef]
- Jin, R.; Jin, Y.Y.; Tang, Y.L.; Yang, H.J.; Zhou, X.Q.; Lei, Z. GPNMB silencing suppresses the proliferation and metastasis of osteosarcoma cells by blocking the PI3K/Akt/mTOR signaling pathway. Oncol. Rep. 2018, 39, 3034–3040. [Google Scholar] [CrossRef]
- Huang, S.; Jin, A. ZIC2 promotes viability and invasion of human osteosarcoma cells by suppressing SHIP2 expression and activating PI3K/AKT pathways. J. Cell. Biochem. 2018, 119, 2248–2257. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yuan, D.; Sun, P.; Liu, W.; Wu, P.F.; Liu, H.; Yu, G.Y. LINC00968 functions as an oncogene in osteosarcoma by activating the PI3K/AKT/mTOR signaling. J. Cell. Physiol. 2018, 233, 8639–8647. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Wu, J.X.; Zhang, Y.; Che, H.; Yang, L. LncRNA LINC00628 overexpression inhibits the growth and invasion through regulating PI3K/Akt signaling pathway in osteosarcoma. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5857–5866. [Google Scholar] [PubMed]
- Jiang, N.; Wang, X.; Xie, X.; Liao, Y.; Liu, N.; Liu, J.; Miao, N.; Shen, J.; Peng, T. lncRNA DANCR promotes tumor progression and cancer stemness features in osteosarcoma by upregulating AXL via miR-33a-5p inhibition. Cancer Lett. 2017, 405, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wang, Q.; Zhou, X.; Fu, C.; Cheng, M.; Guo, R.; Liu, H.; Zhang, B.; Dai, M. Celastrol negatively regulates cell invasion and migration ability of human osteosarcoma via downregulation of the PI3K/Akt/NF-κB signaling pathway in vitro. Oncol. Lett. 2016, 12, 3423–3428. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Liu, G.; Rus, I.A.; Yao, S.; Chen, Y.; Akiri, G.; Grumolato, L.; Aaronson, S.A. High-frequency canonical Wnt activation in multiple sarcoma subtypes drives proliferation through a TCF/β-catenin target gene, CDC25A. Cancer Cell 2011, 19, 601–612. [Google Scholar] [CrossRef]
- Jin, H.; Luo, S.; Wang, Y.; Liu, C.; Piao, Z.; Xu, M.; Guan, W.; Li, Q.; Zou, H.; Tan, Q.Y.; et al. miR-135b Stimulates Osteosarcoma Recurrence and Lung Metastasis via Notch and Wnt/beta-Catenin Signaling. Mol. Ther. Nucleic Acids 2017, 8, 111–122. [Google Scholar] [CrossRef]
- Yang, X.; Wang, L.; Wang, Q.; Li, L.; Fu, Y.; Sun, J. MiR-183 inhibits osteosarcoma cell growth and invasion by regulating LRP6-Wnt/beta-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2018, 496, 1197–1203. [Google Scholar] [CrossRef]
- Xu, E.; Zhao, J.; Ma, J.; Wang, C.; Zhang, C.; Jiang, H.; Cheng, J.; Gao, R.; Zhou, X. miR-146b-5p promotes invasion and metastasis contributing to chemoresistance in osteosarcoma by targeting zinc and ring finger 3. Oncol. Rep. 2016, 35, 275–283. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, L.; Lv, Z.; Li, Q.; Gong, W.; Wu, H. MicroRNA-342-3p Inhibits the Proliferation, Migration, and Invasion of Osteosarcoma Cells by Targeting Astrocyte-Elevated Gene-1 (AEG-1). Oncol. Res. 2017, 25, 1505–1515. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Li, C.B.; Yuan, B.T.; Qi, W.; Li, H.L.; Shen, X.Z.; Zhao, G.; Wang, J.T.; Liu, Y.J. MicroRNA-26a induces osteosarcoma cell growth and metastasis via the Wnt/beta-catenin pathway. Oncol. Lett. 2016, 11, 1592–1596. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.B.; Zhang, Z.C.; Han, G.S.; Han, J.Z.; Qiu, D.P. Overexpression of miR-214 promotes the progression of human osteosarcoma by regulating the Wnt/β-catenin signaling pathway. Mol. Med. Rep. 2017, 15, 1884–1892. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Yang, G.; Jiang, P.; Zhang, L.; Wang, J.; Sun, S. Long non-coding RNA Sox4 promotes proliferation and migration by activating Wnt/β-catenin signaling pathway in osteosarcoma. Pharmazie 2017, 72, 537–542. [Google Scholar]
- Jiang, Z.; Jiang, C.; Fang, J. Upregulated lnc-SNHG1 contributes to osteosarcoma progression through sequestration of miR-577 and activation of WNT2B/Wnt/β-catenin pathway. Biochem. Biophys. Res. Commun. 2018, 495, 238–245. [Google Scholar] [CrossRef]
- Zhao, H.; Hou, W.; Tao, J.; Zhao, Y.; Wan, G.; Ma, C.; Xu, H. Upregulation of lncRNA HNF1A-AS1 promotes cell proliferation and metastasis in osteosarcoma through activation of the Wnt/β-catenin signaling pathway. Am. J. Transl. Res. 2016, 8, 3503–3512. [Google Scholar]
- Vega, O.A.; Lucero, C.M.J.; Araya, H.F.; Jerez, S.; Tapia, J.C.; Antonelli, M.; Salazar-Onfray, F.; Las Heras, F.; Thaler, R.; Riester, S.M.; et al. Wnt/beta-Catenin Signaling Activates Expression of the Bone-Related Transcription Factor RUNX2 in Select Human Osteosarcoma Cell Types. J. Cell. Biochem. 2017, 118, 3662–3674. [Google Scholar] [CrossRef]
- Lv, Y.F.; Dai, H.; Yan, G.N.; Meng, G.; Zhang, X.; Guo, Q.N. Downregulation of tumor suppressing STF cDNA 3 promotes epithelial-mesenchymal transition and tumor metastasis of osteosarcoma by the Wnt/GSK-3β/β-catenin/Snail signaling pathway. Cancer Lett. 2016, 373, 164–173. [Google Scholar] [CrossRef]
- Han, W.; Liu, J. Epigenetic silencing of the Wnt antagonist APCDD1 by promoter DNA hyper-methylation contributes to osteosarcoma cell invasion and metastasis. Biochem. Biophys. Res. Commun. 2017, 491, 91–97. [Google Scholar] [CrossRef]
- McCain, J. The MAPK (ERK) Pathway: Investigational Combinations for the Treatment of BRAF-Mutated Metastatic Melanoma. P&T A Peer-Rev. J. Formul. Manag. 2013, 38, 96–108. [Google Scholar]
- Cheng, G.; Gao, F.; Sun, X.; Bi, H.; Zhu, Y. Paris saponin VII suppresses osteosarcoma cell migration and invasion by inhibiting MMP-2/9 production via the p38 MAPK signaling pathway. Mol. Med. Rep. 2016, 14, 3199–3205. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.M.; Park, B.S.; Kang, H.K.; Park, H.R.; Yu, S.B.; Kim, I.R. Delphinidin induces apoptosis and inhibits epithelial-to-mesenchymal transition via the ERK/p38 MAPK-signaling pathway in human osteosarcoma cell lines. Environ. Toxicol. 2018, 33, 640–649. [Google Scholar] [CrossRef] [PubMed]
- Lopes, L.J.S.; Tesser-Gamba, F.; Petrilli, A.S.; de Seixas Alves, M.T.; Garcia-Filho, R.J.; Toledo, S.R.C. MAPK pathways regulation by DUSP1 in the development of osteosarcoma: Potential markers and therapeutic targets. Mol. Carcinog. 2017, 56, 1630–1641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, Q.; Li, G.; Li, L.; Liang, S.; Zhang, Y.; Liu, J.; Fan, Z.; Li, L.; Zhou, B.; et al. ONZIN Upregulation by Mutant p53 Contributes to Osteosarcoma Metastasis Through the CXCL5-MAPK Signaling Pathway. Cell. Physiol. Biochem. 2018, 48, 1099–1111. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, X.; Li, W.; Li, M.; Tu, T.; Ba, X.; Wu, Y.; Huang, Z.; Fan, G.; Zhou, G.; et al. Macrophage migration inhibitory factor promotes osteosarcoma growth and lung metastasis through activating the RAS/MAPK pathway. Cancer Lett. 2017, 403, 271–279. [Google Scholar] [CrossRef]
- Salas, S.; Jiguet-Jiglaire, C.; Campion, L.; Bartoli, C.; Frassineti, F.; Deville, J.L.; Maues De Paula, A.; Forest, F.; Jézéquel, P.; Gentet, J.C.; et al. Correlation between ERK1 and STAT3 expression and chemoresistance in patients with conventional osteosarcoma. BMC Cancer 2014, 14, 606. [Google Scholar] [CrossRef]
- Pencik, J.; Pham, H.T.; Schmoellerl, J.; Javaheri, T.; Schlederer, M.; Culig, Z.; Merkel, O.; Moriggl, R.; Grebien, F.; Kenner, L. JAK-STAT signaling in cancer: From cytokines to non-coding genome. Cytokine 2016, 87, 26–36. [Google Scholar] [CrossRef]
- Jiang, R.; Zhang, C.; Liu, G.; Gu, R.; Wu, H. MicroRNA-126 Inhibits Proliferation, Migration, Invasion, and EMT in Osteosarcoma by Targeting ZEB1. J. Cell. Biochem. 2017, 118, 3765–3774. [Google Scholar] [CrossRef]
- Liu, L.H.; Li, H.; Li, J.P.; Zhong, H.; Zhang, H.C.; Chen, J.; Xiao, T. miR-125b suppresses the proliferation and migration of osteosarcoma cells through down-regulation of STAT3. Biochem. Biophys. Res. Commun. 2011, 416, 31–38. [Google Scholar] [CrossRef]
- Lum, L.; Beachy, P.A. The Hedgehog response network: Sensors, switches, and routers. Science 2004, 304, 1755–1759. [Google Scholar] [CrossRef]
- Yao, Z.; Han, L.; Chen, Y.; He, F.; Sun, B.; Kamar, S.; Zhang, Y.; Yang, Y.; Wang, C.; Yang, Z. Hedgehog signalling in the tumourigenesis and metastasis of osteosarcoma, and its potential value in the clinical therapy of osteosarcoma. Cell Death Dis. 2018, 9, 701. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Takebe, N.; Lorusso, P. Targeting the Hedgehog pathway in cancer. Ther. Adv. Med. Oncol. 2010, 2, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.H.; Wang, W.; Yeung, W.; Deng, Y.; Yuan, P.; Mak, K.K. Hedgehog signaling induces osteosarcoma development through Yap1 and H19 overexpression. Oncogene 2014, 33, 4857–4866. [Google Scholar] [CrossRef]
- Mohseny, A.B.; Cai, Y.; Kuijjer, M.; Xiao, W.; van den Akker, B.; de Andrea, C.E.; Jacobs, R.; ten Dijke, P.; Hogendoorn, P.C.; Cleton-Jansen, A.M. The activities of Smad and Gli mediated signalling pathways in high-grade conventional osteosarcoma. Eur. J. Cancer 2012, 48, 3429–3438. [Google Scholar] [CrossRef] [PubMed]
- Nagao-Kitamoto, H.; Setoguchi, T.; Kitamoto, S.; Nakamura, S.; Tsuru, A.; Nagata, M.; Nagano, S.; Ishidou, Y.; Yokouchi, M.; Kitajima, S.; et al. Ribosomal protein S3 regulates GLI2-mediated osteosarcoma invasion. Cancer Lett. 2015, 356 Pt B, 855–861. [Google Scholar] [CrossRef]
- Lo, W.W.; Wunder, J.S.; Dickson, B.C.; Campbell, V.; McGovern, K.; Alman, B.A.; Andrulis, I.L. Involvement and targeted intervention of dysregulated Hedgehog signaling in osteosarcoma. Cancer 2014, 120, 537–547. [Google Scholar] [CrossRef]
- Zhao, Z.; Jia, Q.; Wu, M.S.; Xie, X.; Wang, Y.; Song, G.; Zou, C.Y.; Tang, Q.; Lu, J.; Huang, G.; et al. Degalactotigonin, a Natural Compound from Solanum nigrum L., Inhibits Growth and Metastasis of Osteosarcoma through GSK3β Inactivation-Mediated Repression of the Hedgehog/Gli1 Pathway. Clin. Cancer Res. 2018, 24, 130–144. [Google Scholar] [CrossRef]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch signaling: Cell fate control and signal integration in development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, Y.; Nolo, R.; Zweidler-McKay, P.A.; Hughes, D.P. Regulation of NOTCH signaling by reciprocal inhibition of HES1 and Deltex 1 and its role in osteosarcoma invasiveness. Oncogene 2010, 29, 2916–2926. [Google Scholar] [CrossRef]
- Zheng, L.; Conner, S.D. PI5P4Kγ functions in DTX1-mediated Notch signaling. Proc. Natl. Acad. Sci. USA 2018, 115, E1983–E1990. [Google Scholar] [CrossRef]
- Zhou, S.; Yu, L.; Xiong, M.; Dai, G. LncRNA SNHG12 promotes tumorigenesis and metastasis in osteosarcoma by upregulating Notch2 by sponging miR-195-5p. Biochem. Biophys. Res. Commun. 2018, 495, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tang, Y.; Xing, W.; Dong, W.; Wang, Z. LncRNA, CRNDE promotes osteosarcoma cell proliferation, invasion and migration by regulating Notch1 signaling and epithelial-mesenchymal transition. Exp. Mol. Pathol. 2018, 104, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Angulo, P.; Kaushik, G.; Subramaniam, D.; Dandawate, P.; Neville, K.; Chastain, K.; Anant, S. Natural compounds targeting major cell signaling pathways: A novel paradigm for osteosarcoma therapy. J. Hematol. Oncol. 2017, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Liapis, V.; Labrinidis, A.; Zinonos, I.; Hay, S.; Ponomarev, V.; Panagopoulos, V.; DeNichilo, M.; Ingman, W.; Atkins, G.J.; Findlay, D.M.; et al. Hypoxia-activated pro-drug TH-302 exhibits potent tumor suppressive activity and cooperates with chemotherapy against osteosarcoma. Cancer Lett. 2015, 357, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Avnet, S.; Lemma, S.; Cortini, M.; Pellegrini, P.; Perut, F.; Zini, N.; Kusuzaki, K.; Chano, T.; Grisendi, G.; Dominici, M.; et al. Altered pH gradient at the plasma membrane of osteosarcoma cells is a key mechanism of drug resistance. Oncotarget 2016, 7, 63408–63423. [Google Scholar] [CrossRef]
- Wiklander, O.P.B.; Brennan, M.; Lötvall, J.; Breakefield, X.O.; El Andaloussi, S. Advances in therapeutic applications of extracellular vesicles. Sci. Transl. Med. 2019, 11, eaav8521. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Factor | Source | Pathway | Target | In Vitro or In Vivo | References |
|---|---|---|---|---|---|
| CXCL1 (GRO-α) | MSCs | NF-κB pathway in MSCs | OS cells | In vitro | [31] |
| OS cells | Trans-differentiate into cancer-associated fibroblasts in MSCs | MSCs | In vitro | [32] | |
| cancer-associated fibroblasts | MAT in OS cells | OS cells | In vitro | [32] | |
| CXCL5 | MSCs | NF-κB pathway in MSCs | OS cells | In vitro | [31] |
| IL-6 | MSCs | IL-6/STAT3 signaling pathway in OS cells | OS cells | In vitro and in vivo | [33] |
| MSCs | NF-κB pathway in MSCs | OS cells | In vitro | [31] | |
| cancer-associated fibroblasts | MAT in OS cells | OS cells | In vitro | [32] | |
| IL-8 | MSCs | NF-κB pathway in MSCs | OS cells | In vitro | [31] |
| MSCs and OS cells | IL-8/CXCR1/Akt signaling pathway, MAT | OS cells | In vitro and in vivo | [34] | |
| MSCs, cancer-associated fibroblasts | MAT in OS cells | OS cells | In vitro | [32] | |
| CXCL12 | MSCs | CXCL12/CXCR4/CXCR7 | OS cells | In vitro | [27,35] |
| CCL2 (MCP-1) | OS cells, cancer-associated fibroblasts | MAT in OS cells | OS cells | In vitro | [32] |
| CCL5 | MSCs | NF-κB pathway in MSCs | OS cells | In vitro | [31] |
| Lactate | MSCs | metabolic reprogramming OS cells | OS cells | In vitro | [30] |
| BMP-2 | MSCs | NF-κB pathway in MSCs | OS cells | In vitro | [31] |
| NF-κB1, RelA, RelB | MSCs | NF-κB pathway in MSCs | OS cells | In vitro | [31] |
| CSF2/GM-CSF | MSCs | NF-κB pathway in MSCs | OS cells | In vitro | [31] |
| CSF3/G-CSF | MSCs | NF-κB pathway in MSCs | OS cells | In vitro | [31] |
| TGF-β | OS extracellular vesicle | IL-6/STAT3 signaling pathway in OS | MSCs | In vitro and in vivo | [32,33] |
| has-mir-195 | MSCs extracellular vesicle | FAK/PTK2 in OS | OS cells | In vitro | [36] |
| has-mir-124 | MSCs extracellular vesicle | Has-mir-124/Rac1 | OS cells | In vitro | [36,37] |
| Physical Element | Factors | Interacting Molecule | Efficacy | Reference |
|---|---|---|---|---|
| Hypoxia | HIF-1α | CXCR4 | migration ↑ | [37] |
| TGF-β | osteolytic bone metastases ↑ | [40] | ||
| miR-20b | invasion and proliferation ↑ | [35] | ||
| miR-33b | [41] | |||
| BMPR2 | distant metastasis and poor survival rate ↑ | [42] | ||
| LncRNA MALAT1 | pro-angiogenic ↑ | [43] | ||
| ANGPTL4 | migration, proliferation ↑ | [44] | ||
| HIF2PUT | HIF2α | distant metastasis ↑ | [45] | |
| Acidic condition | CXCL1 | growth, metastasis ↑ | [31] | |
| CXCL2 | ||||
| CXCL5 | ||||
| CXCR4 | ||||
| BMP2 | colony formation ↑ | [31] | ||
| CSF2/GM-CSF | ||||
| CSF3/G-CSF | ||||
| IL1A | ||||
| IL1RN | ||||
| IL23A | ||||
| IL-6 | ||||
| IL-8 | ||||
| MMP2 | ||||
| NFκB1 | ||||
| RelA | ||||
| RelB |
| Non-coding RNA | Target Molecule | Effect | In Vitro or In Vivo | Reference |
|---|---|---|---|---|
| miR-135b, | GSK3β, CK1a, TET3 | lung metastasis, tumor recurrence ↑ | In vitro or in vivo | [78] |
| miR-183 | LRP6 | migration, invasion ↑ | In vitro | [79] |
| miR-146b-5p | ZNRF3 | invasion, metastasis, chemoresistance ↑ | In vitro | [80] |
| miR-26a | GSK-3β | proliferation, migration, invasion ↑ | In vitro | [82] |
| miR-214 | β-catenin | proliferation ↑ | In vitro | [83] |
| miR-342-3p | AEG-1 | proliferation, migration, invasion ↑ | In vitro | [81] |
| LncSox4 | β-catenin | cell viability ↑ | In vitro | [84] |
| Lnc-SNHG1 | miR-557/WNT2B | migration, EMT process, tumor growth ↑ | In vitro or in vivo | [85] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Tian, Y.; Zhao, F.; Chen, Z.; Su, P.; Li, Y.; Qian, A. Bone Microenvironment and Osteosarcoma Metastasis. Int. J. Mol. Sci. 2020, 21, 6985. https://doi.org/10.3390/ijms21196985
Yang C, Tian Y, Zhao F, Chen Z, Su P, Li Y, Qian A. Bone Microenvironment and Osteosarcoma Metastasis. International Journal of Molecular Sciences. 2020; 21(19):6985. https://doi.org/10.3390/ijms21196985
Chicago/Turabian StyleYang, Chaofei, Ye Tian, Fan Zhao, Zhihao Chen, Peihong Su, Yu Li, and Airong Qian. 2020. "Bone Microenvironment and Osteosarcoma Metastasis" International Journal of Molecular Sciences 21, no. 19: 6985. https://doi.org/10.3390/ijms21196985
APA StyleYang, C., Tian, Y., Zhao, F., Chen, Z., Su, P., Li, Y., & Qian, A. (2020). Bone Microenvironment and Osteosarcoma Metastasis. International Journal of Molecular Sciences, 21(19), 6985. https://doi.org/10.3390/ijms21196985