LH-induced Transcriptional Regulation of Klf4 Expression in Granulosa Cells Occurs via the cAMP/PKA Pathway and Requires a Putative Sp1 Binding Site


Abstract

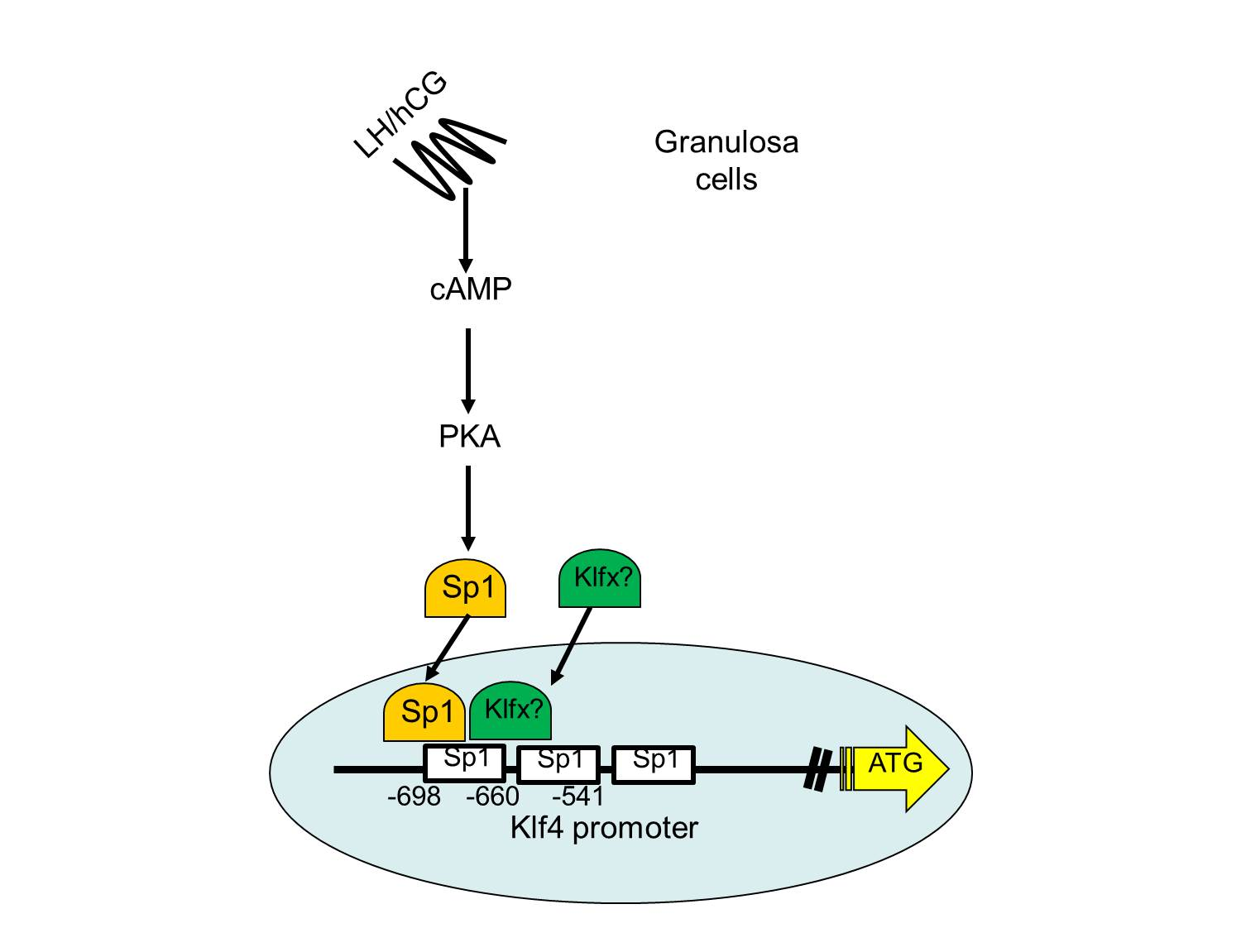{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. LH Induces Transient Klf4 Expression in Cultured Preovulatory GCs
2.2. LH Modulates Klf4 Transcription via A cAMP/PKA-Dependent Signaling Pathway
2.3. LH-Induced Increases in Klf4 Promoter Activity Require Regulatory Elements between −715 and −402 bp of The 5′ region of The Klf4 Promoter
2.4. LH-Induced Upregulation of Klf4 Is Mediated by The Sp1 Binding Motif in GCs
2.5. Sp1 Binds to The Klf4 Promoter Region in Vivo
2.6. Mutation of The Sp-1 Binding Site at −698/−688 Results in Loss of LH-Induced Klf4 Promoter Activity
3. Discussion
4. Materials and Methods
4.1. Animals and Reagents
4.2. Preparation and Culture of Preovulatory Granulosa Cells
4.3. Plasmids
4.4. Transient Transfection and Luciferase Reporter Assays
4.5. Real Time Quantitative PCR
4.6. Western Blot Analysis
4.7. Chromatin Immunoprecipitation (ChIP) Assays
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Richards, J.S. Ovulation: New factors that prepare the oocyte for fertilization. Mol. Cell. Endocrinol. 2005, 234, 75–79. [Google Scholar] [CrossRef]
- Richards, J.S.; Russell, D.L.; Ochsner, S.; Espey, L.L. Ovulation: New dimensions and new regulators of the inflammatory-like response. Annu. Rev. Physiol. 2002, 64, 69–92. [Google Scholar] [CrossRef]
- Choi, H.; Ryu, K.-Y.; Roh, J. Krüppel-like factor 4 plays a role in the luteal transition in steroidogenesis by downregulating Cyp19A1 expression. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E1071–E1080. [Google Scholar] [CrossRef]
- Dang, D.T.; Pevsner, J.; Yang, V.W. The biology of the mammalian Krüppel-like family of transcription factors. Int. J. Biochem. Cell Biol. 2000, 32, 1103–1121. [Google Scholar] [CrossRef]
- Garrett-Sinha, L.A.; Eberspaecher, H.; Seldin, M.F.; de Crombrugghe, B. A gene for a novel zinc-finger protein expressed in differentiated epithelial cells and transiently in certain mesenchymal cells. J. Biol. Chem. 1996, 271, 31384–31390. [Google Scholar] [CrossRef]
- Katz, J.P.; Perreault, N.; Goldstein, B.G.; Lee, C.S.; Labosky, P.A.; Yang, V.W.; Kaestner, K.H. The zinc-finger transcription factor Klf4 is required for terminal differentiation of goblet cells in the colon. Development 2002, 129, 2619–2628. [Google Scholar] [PubMed]
- Shields, J.M.; Christy, R.J.; Yang, V.W. Identification and characterization of a gene encoding a gut-enriched Krüppel-like factor expressed during growth arrest. J. Biol. Chem. 1996, 271, 20009–20017. [Google Scholar] [CrossRef]
- Mahatan, C.S.; Kaestner, K.H.; Geiman, D.E.; Vincent, W.Y. Characterization of the structure and regulation of the murine gene encoding gut-enriched Krüppel-like factor (Krüppel-like factor 4). Nucleic Acids Res. 1999, 27, 4562–4569. [Google Scholar] [CrossRef]
- Godmann, M.; Katz, J.P.; Guillou, F.; Simoni, M.; Kaestner, K.H.; Behr, R. Krüppel-like factor 4 is involved in functional differentiation of testicular Sertoli cells. Dev. Biol. 2008, 315, 552–566. [Google Scholar] [CrossRef]
- Liu, H.C.; He, Z.; Rosenwaks, Z. Application of complementary DNA microarray (DNA chip) technology in the study of gene expression profiles during folliculogenesis. Fertil. Steril. 2001, 75, 947–955. [Google Scholar] [CrossRef]
- Choi, H.; Roh, J. Role of Klf4 in the regulation of apoptosis and cell cycle in rat granulosa cells during the periovulatory period. Int. J. Mol. Sci. 2019, 20, 87. [Google Scholar] [CrossRef]
- Jansen, E.; Laven, J.S.; Dommerholt, H.B.; Polman, J.; van Rijt, C.; van den Hurk, C.; Westland, J.; Mosselman, S.; Fauser, B.C. Abnormal gene expression profiles in human ovaries from polycystic ovary syndrome patients. Mol. Endocrinol. 2004, 18, 3050–3063. [Google Scholar] [CrossRef]
- Sivritas, D.; Becher, M.U.; Ebrahimian, T.; Arfa, O.; Rapp, S.; Bohner, A.; Mueller, C.F.; Umemura, T.; Wassmann, S.; Nickenig, G. Antiproliferative effect of estrogen in vascular smooth muscle cells is mediated by Kruppel-like factor-4 and manganese superoxide dismutase. Basic Res. Cardiol. 2011, 106, 563–575. [Google Scholar] [CrossRef]
- Birsoy, K.; Chen, Z.; Friedman, J. Transcriptional regulation of adipogenesis by KLF4. Cell Metab. 2008, 7, 339–347. [Google Scholar] [CrossRef]
- Cullingford, T.E.; Butler, M.J.; Marshall, A.K.; Sugden, P.H.; Clerk, A. Differential regulation of Krüppel-like factor family transcription factor expression in neonatal rat cardiac myocytes: Effects of endothelin-1, oxidative stress and cytokines. Biochim. Et Biophys. Acta BBA Mol. Cell Res. 2008, 1783, 1229–1236. [Google Scholar] [CrossRef]
- McLean, D.J.; Friel, P.J.; Pouchnik, D.; Griswold, M.D. Oligonucleotide microarray analysis of gene expression in follicle-stimulating hormone-treated rat Sertoli cells. Mol. Endocrinol. 2002, 16, 2780–2792. [Google Scholar] [CrossRef]
- Natesampillai, S.; Kerkvliet, J.; Leung, P.C.; Veldhuis, J.D. Regulation of Kruppel-like factor 4, 9, and 13 genes and the steroidogenic genes LDLR, StAR, and CYP11A in ovarian granulosa cells. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E385–E391. [Google Scholar] [CrossRef]
- Channing, C.P.; Tsafriri, A. Mechanism of action of luteinizing hormone and follicle-stimulating hormone on the ovary in vitro. Metabolism 1977, 26, 413–468. [Google Scholar] [CrossRef]
- El-Fouly, M.A.; Cook, B.; Nekola, M.; Nalbandov, A.V. Role of the Ovum in Follicular Luteinization. Endocrinology 1970, 87, 288–293. [Google Scholar] [CrossRef]
- Morris, J.; Richards, J. Luteinizing hormone induces prostaglandin endoperoxide synthase-2 and luteinization in vitro by A-kinase and C-kinase pathways. Endocrinology 1995, 136, 1549–1558. [Google Scholar] [CrossRef]
- Dijkmans, T.; van Hooijdonk, L.; Schouten, T.; Kamphorst, J.; Fitzsimons, C.; Vreugdenhil, E. Identification of new Nerve Growth Factor-responsive immediate-early genes. Brain Res. 2009, 1249, 19–33. [Google Scholar] [CrossRef]
- Godmann, M.; Kosan, C.; Behr, R. Krüppel-like factor 4 is widely expressed in the mouse male and female reproductive tract and responds as an immediate early gene to activation of the protein kinase A in TM4 Sertoli cells. Reproduction 2010, 139, 771–782. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Wang, X.; Zhou, Y.; Offner, G.; Tseng, C.-C. Destabilization of Krüppel-like factor 4 protein in response to serum stimulation involves the ubiquitin-proteasome pathway. Cancer Res. 2005, 65, 10394–10400. [Google Scholar] [CrossRef][Green Version]
- Rogers, S.; Wells, R.; Rechsteiner, M. Amino acid sequences common to rapidly degraded proteins: The PEST hypothesis. Science 1986, 234, 364–368. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Rex, S.; Tseng, C.C. Kruppel-like factor 4 is transactivated by butyrate in colon cancer cells. J. Nutr. 2004, 134, 792–798. [Google Scholar] [CrossRef]
- Horton, C.D.; Halvorson, L.M. The cAMP signaling system regulates LHbeta gene expression: Roles of early growth response protein-1, SP1 and steroidogenic factor-1. J. Mol. Endocrinol. 2004, 32, 291–306. [Google Scholar] [CrossRef]
- Alliston, T.N.; Maiyar, A.C.; Buse, P.; Firestone, G.L.; Richards, J.S. Follicle stimulating hormone-regulated expression of serum/glucocorticoid-inducible kinase in rat ovarian granulosa cells: A functional role for the Sp1 family in promoter activity. Mol. Endocrinol. (Baltim. Md.) 1997, 11, 1934–1949. [Google Scholar] [CrossRef][Green Version]
- Singh, D.; Hui, Y.Y.; LaVoie, H.A. Expression, localization, and hormonal regulation of LDL receptor, Sp1, and SREBP-1 in the porcine ovary. Biol. Reprod. 2003, 68, 239–240. [Google Scholar]
- Momoi, K.; Waterman, M.R.; Simpson, E.R.; Zanger, U.M. 3′,5′-cyclic adenosine monophosphate-dependent transcription of the CYP11A (cholesterol side chain cleavage cytochrome P450) gene involves a DNA response element containing a putative binding site for transcription factor Sp1. Mol. Endocrinol. (Baltim. Md.) 1992, 6, 1682–1690. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rohlff, C.; Ahmad, S.; Borellini, F.; Lei, J.; Glazer, R.I. Modulation of transcription factor Sp1 by cAMP-dependent protein kinase. J. Biol. Chem. 1997, 272, 21137–21141. [Google Scholar] [CrossRef]
- Lu, S.; Jenster, G.; Epner, D.E. Androgen induction of cyclin-dependent kinase inhibitor p21 gene: Role of androgen receptor and transcription factor Sp1 complex. Mol. Endocrinol. (Baltim. Md.) 2000, 14, 753–760. [Google Scholar] [CrossRef]
- Deaton, R.A.; Gan, Q.; Owens, G.K. Sp1-dependent activation of KLF4 is required for PDGF-BB-induced phenotypic modulation of smooth muscle. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1027–H1037. [Google Scholar] [CrossRef]
- Sun, Z.Y.; Yu, S.T.; Chen, S.; Liu, H.; Chen, Z. SP1 regulates KLF4 via SP1 binding motif governed by DNA methylation during odontoblastic differentiation of human dental pulp cells. J. Cell. Biochem. 2019, 120, 14688–14699. [Google Scholar] [CrossRef]
- Suske, G.; Bruford, E.; Philipsen, S. Mammalian SP/KLF transcription factors: Bring in the family. Genomics 2005, 85, 551–556. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, H.; Roh, J. LH-induced Transcriptional Regulation of Klf4 Expression in Granulosa Cells Occurs via the cAMP/PKA Pathway and Requires a Putative Sp1 Binding Site. Int. J. Mol. Sci. 2020, 21, 7385. https://doi.org/10.3390/ijms21197385
Choi H, Roh J. LH-induced Transcriptional Regulation of Klf4 Expression in Granulosa Cells Occurs via the cAMP/PKA Pathway and Requires a Putative Sp1 Binding Site. International Journal of Molecular Sciences. 2020; 21(19):7385. https://doi.org/10.3390/ijms21197385
Chicago/Turabian StyleChoi, Hyeonhae, and Jaesook Roh. 2020. "LH-induced Transcriptional Regulation of Klf4 Expression in Granulosa Cells Occurs via the cAMP/PKA Pathway and Requires a Putative Sp1 Binding Site" International Journal of Molecular Sciences 21, no. 19: 7385. https://doi.org/10.3390/ijms21197385
APA StyleChoi, H., & Roh, J. (2020). LH-induced Transcriptional Regulation of Klf4 Expression in Granulosa Cells Occurs via the cAMP/PKA Pathway and Requires a Putative Sp1 Binding Site. International Journal of Molecular Sciences, 21(19), 7385. https://doi.org/10.3390/ijms21197385