Parainfluenza Virus 5 Infection in Neurological Disease and Encephalitis of Cattle


Abstract
:1. Introduction
2. Results
2.1. Molecular Confirmation of Viruses
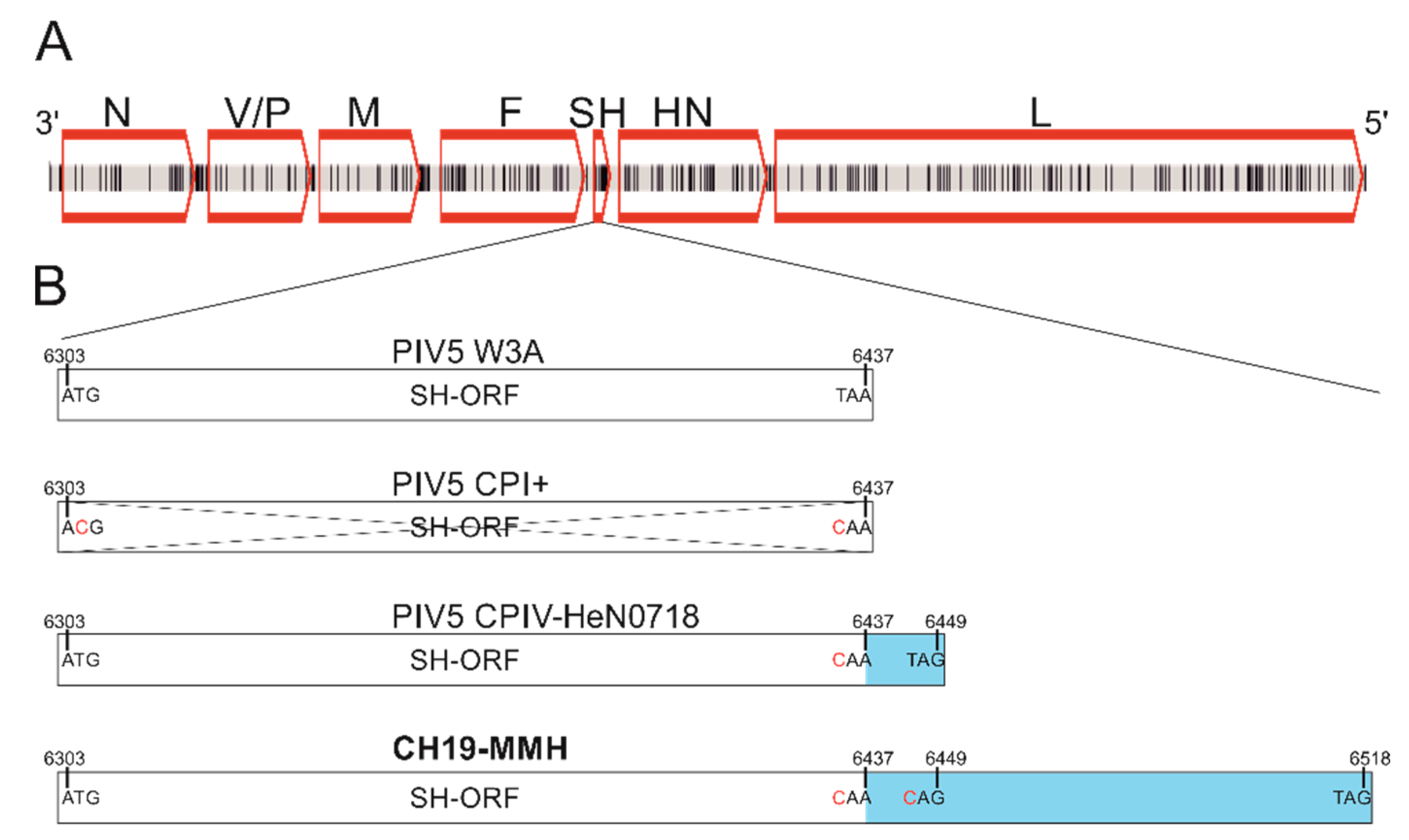
2.2. Genome Sequences
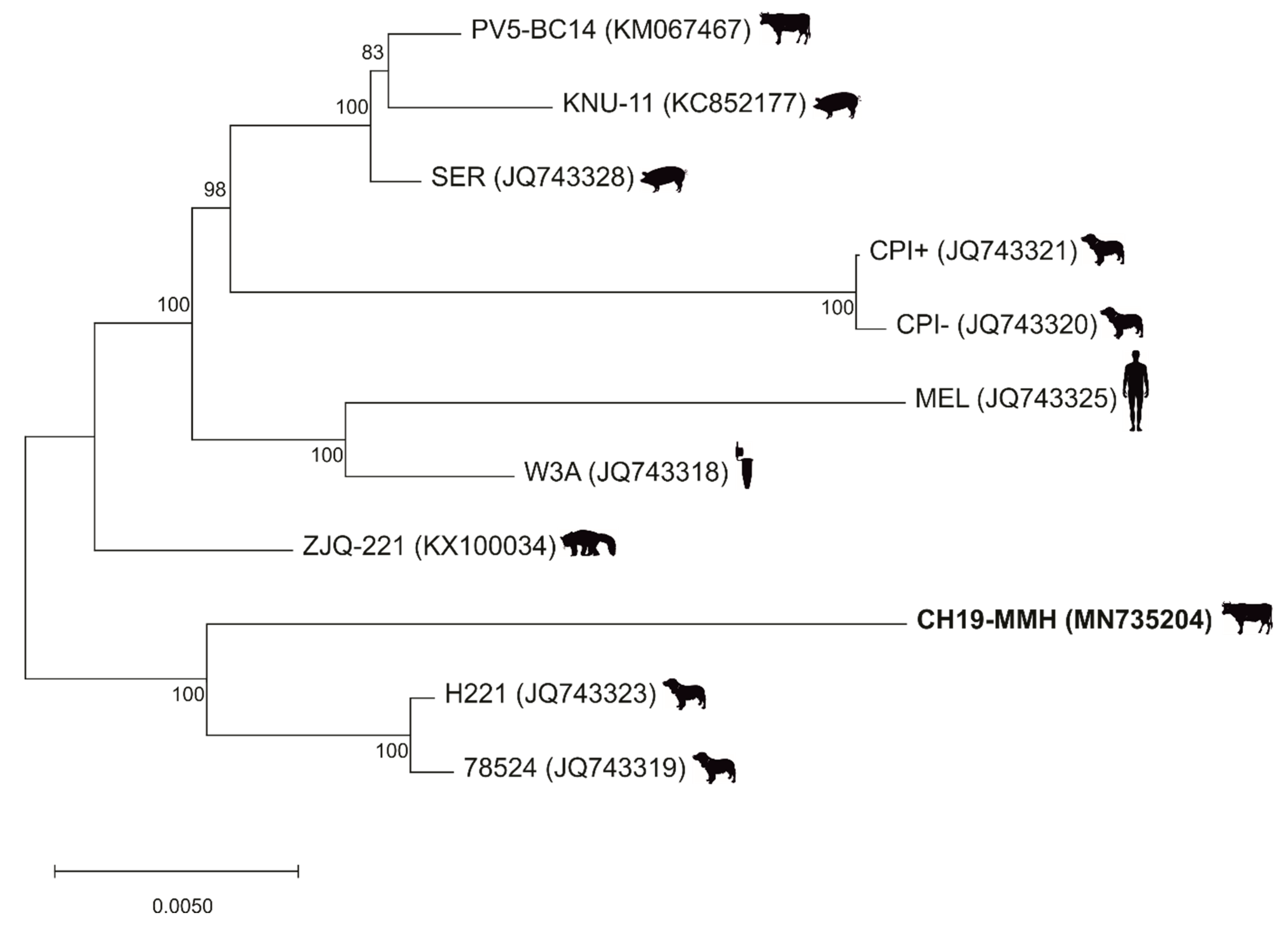
2.3. Phylogenetic Analysis
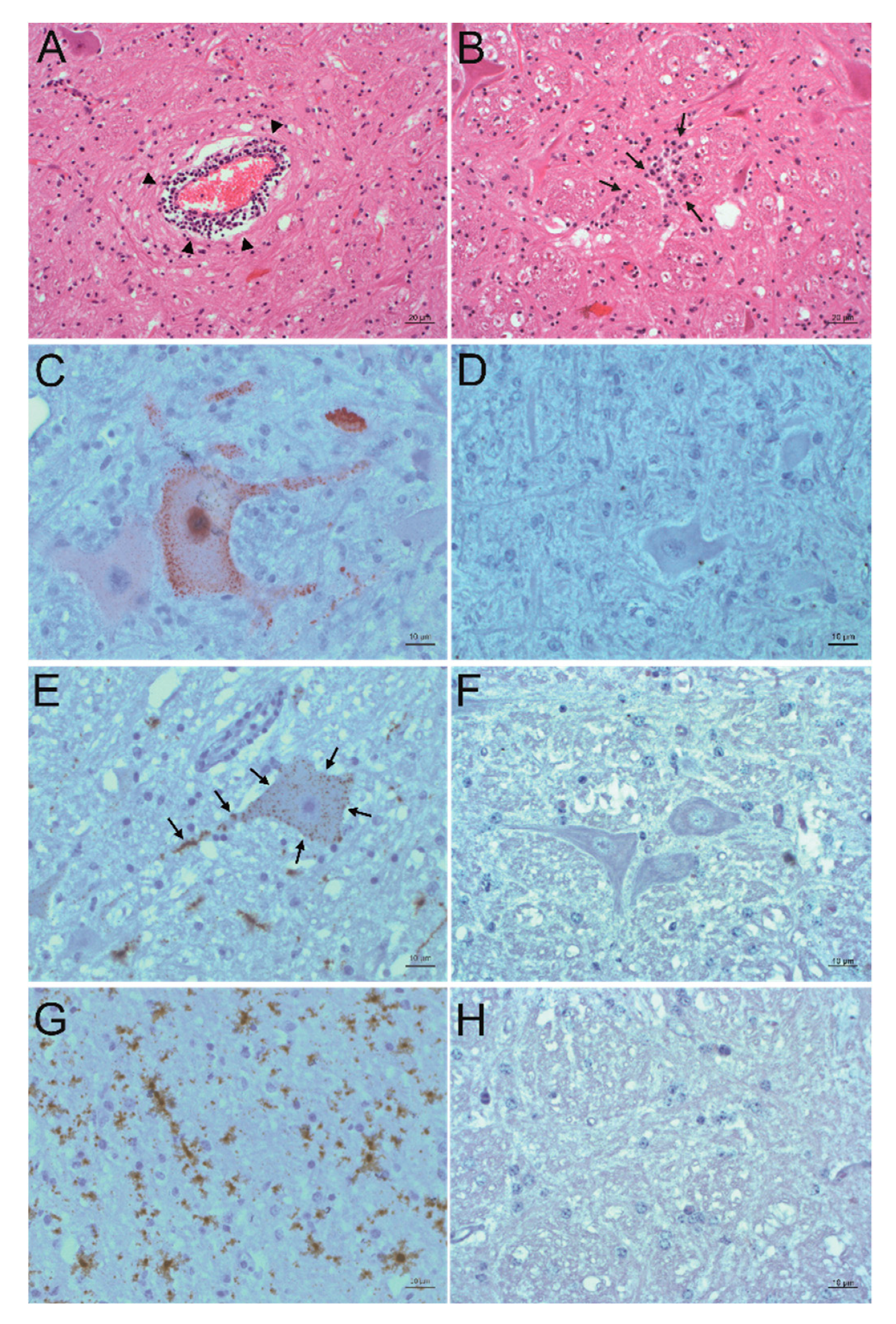
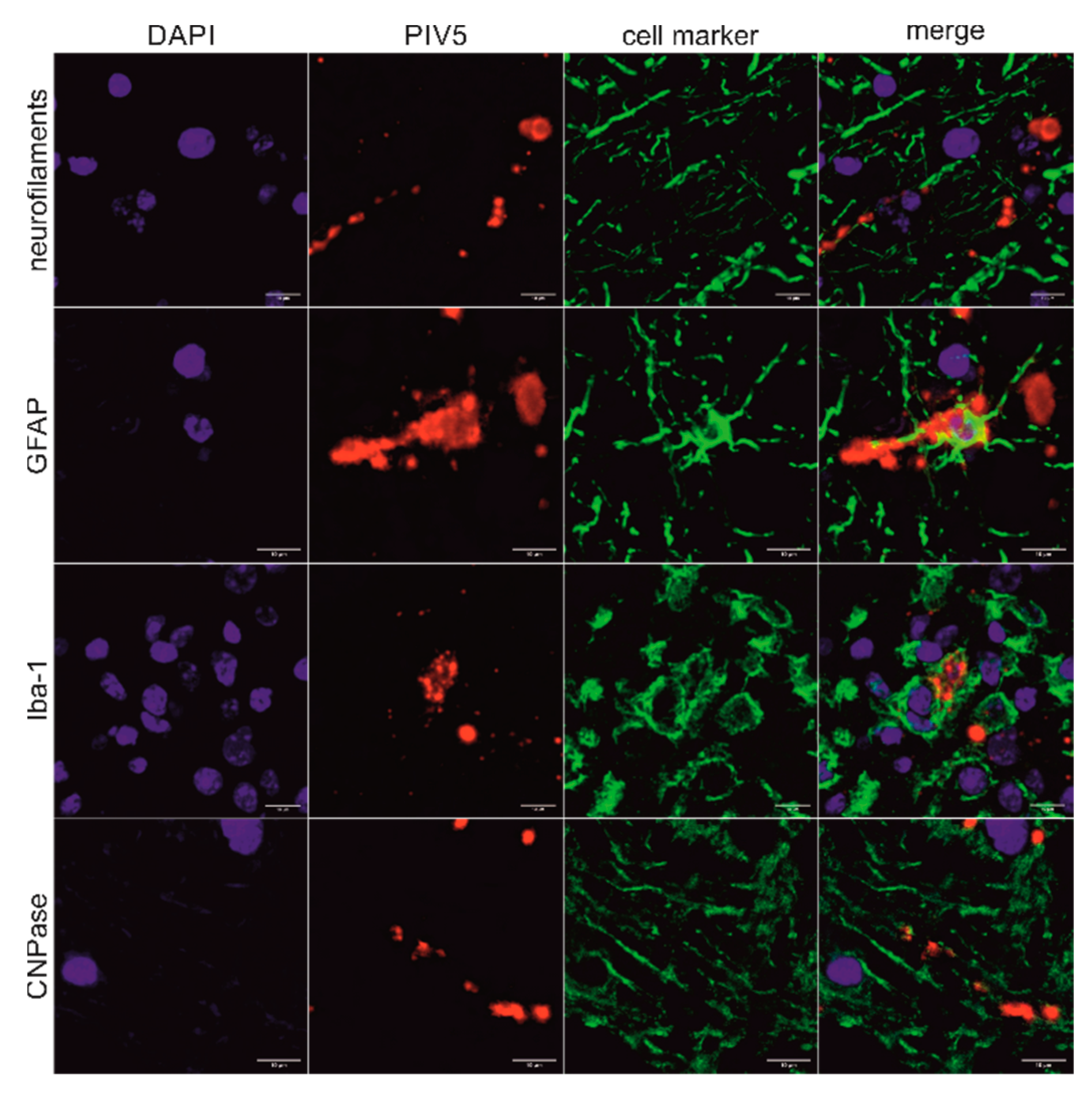
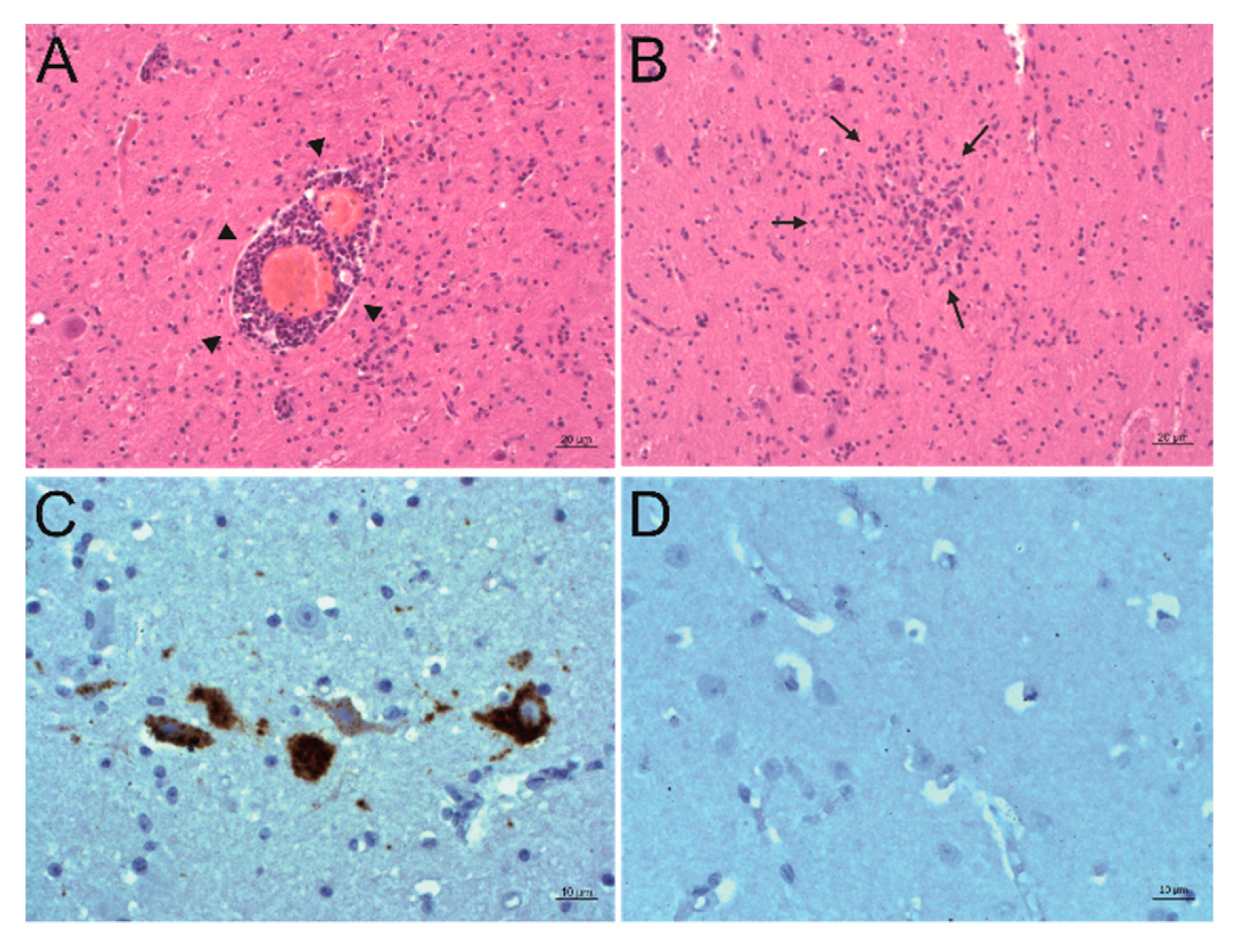
2.4. In Situ Detection of PIV5
2.5. In Situ Detection of Other Viruses
2.6. Comparison of Viral Copy Numbers
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. Nucleic Acid Extraction
4.3. PCR
4.4. Copy Number Determination
4.5. Sanger Sequencing
4.6. Rapid Amplification of cDNA Ends
4.7. Genome Comparison
4.8. Phylogenetic Tree
4.9. Immunohistochemistry
4.10. In Situ Hybridization
4.11. Combination of In Situ Hybridization and Immunofluorescence
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BoHV6 | bovine herpesvirus 6 |
| BoAstV CH13 | bovine astrovirus CH13 |
| BoPyV2 | bovine polyomavirus 2 |
| BSE | bovine spongiform encephalopathy |
| CNPase ESBE | 2′,3′-cyclic-nucleotide 3′-phosphodiesterase European Sporadic Bovine Encephalomyelitis |
| F | fusion protein |
| FFPE | formalin-fixed, paraffin-embedded |
| GFAP | glial fibrillary acidic protein |
| H&E | hematoxylin and eosin |
| HN | hemagglutinin-neuraminidase protein |
| Iba-1 | ionized calcium-binding adapter molecule 1 |
| IF | immunofluorescence |
| IHC | immunohistochemistry |
| ISH | in situ hybridization |
| L | large protein |
| M | matrix protein |
| N | nucleocapsid protein |
| NGS | Next Generation Sequencing |
| ORF | open reading frame |
| P | phosphoprotein |
| PBMCs | peripheral blood mononuclear cells |
| PIV5 | parainfluenza virus 5 |
| RACE | rapid amplification of cDNA ends |
| SH | small hydrophobic protein |
| SV5 | simian virus 5 |
| V | V-protein |
References
- Olival, K.J.; Daszak, P. The ecology of emerging neurotropic viruses. J. Neurovirol. 2005, 11, 441–446. [Google Scholar] [CrossRef]
- Taylor, L.H.; Latham, S.M.; Woolhouse, M.E. Risk factors for human disease emergence. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 983–989. [Google Scholar] [CrossRef]
- Heim, D.; Fatzer, R.; Hornlimann, B.; Vandevelde, M. Frequency of neurologic diseases in cattle. Schweiz. Arch. Tierheilkd. 1997, 139, 354–362. [Google Scholar]
- Fatzer, R.; Steck, F. Histological differential diagnosis in cattle suspected of rabies. Schweiz. Schweiz. Arch. Tierheilkd. 1974, 116, 347–356. [Google Scholar] [PubMed]
- Vandevelde, M.; Higgins, R.J.; Oevermann, A. Veterinary Neuropathology: Essentials of Theory and Practice, 1st ed.; Wiley-Blackwell: Oxford, UK, 2012. [Google Scholar]
- Fooks, A.R.; Cliquet, F.; Finke, S.; Freuling, C.; Hemachudha, T.; Mani, R.S.; Muller, T.; Nadin-Davis, S.; Picard-Meyer, E.; Wilde, H.; et al. Rabies. Nat. Rev. Dis. Primers 2017, 3, 17091. [Google Scholar] [CrossRef] [PubMed]
- Wilde, H.; Hemachudha, T.; Wacharapluesadee, S.; Lumlertdacha, B.; Tepsumethanon, V. Rabies in Asia: The classical zoonosis. Curr. Top. Microbiol. Immunol. 2013, 365, 185–203. [Google Scholar] [CrossRef] [PubMed]
- Callan, R.J.; Van Metre, D.C. Viral diseases of the ruminant nervous system. Vet. Clin. N. Am. Food Anim. Pract. 2004, 20, 327–362. [Google Scholar] [CrossRef]
- Dauphin, G.; Legay, V.; Pitel, P.H.; Zientara, S. Borna disease: Current knowledge and virus detection in France. Vet. Res. 2002, 33, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Burrows, R. Herpes virus infections of animals—A brief review. J. Antimicrobial Chemother. 1977, 3, 9–14. [Google Scholar] [CrossRef]
- Herder, V.; Wohlsein, P.; Peters, M.; Hansmann, F.; Baumgartner, W. Salient lesions in domestic ruminants infected with the emerging so-called Schmallenberg virus in Germany. Vet. Pathol. 2012, 49, 588–591. [Google Scholar] [CrossRef]
- Gibbens, N. Schmallenberg virus: A novel viral disease in northern Europe. Vet. Rec. 2012, 170, 58. [Google Scholar] [CrossRef] [PubMed]
- Theil, D.; Fatzer, R.; Schiller, I.; Caplazi, P.; Zurbriggen, A.; Vandevelde, M. Neuropathological and aetiological studies of sporadic non-suppurative meningoencephalomyelitis of cattle. Vet. Rec. 1998, 143, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Clark, E.G.; Wobeser, G.A.; Janzen, E.D.; Philibert, H. A retrospective study of non-suppurative encephalitis in beef cattle from western Canada. Can. Vet. J. 2013, 54, 1127–1132. [Google Scholar]
- Fankhauser, R. Sporadic meningo-encephalomyelitis in cattle. Schweiz. Arch. Tierheilkd. 1961, 103, 225–235. [Google Scholar]
- Bachmann, P.A.; ter Meulen, V.; Jentsch, G.; Appel, M.; Iwasaki, Y.; Meyermann, R.; Koprowski, H.; Mayr, A. Sporadic bovine meningo-encephalitis-isolation of a paramyxovirus. Arch. Virol. 1975, 48, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Bestetti, G.; Fatzer, R.; Fankhauser, R. Ultrastructural investigations concerning a sporadically occurring meningo-encephalomyelitis of cattle in Switzerland. Schweiz. Arch. Tierheilkd. 1976, 118, 351–357. [Google Scholar]
- Bozzetta, E.; Caramelli, M.; Casalone, C.; Acutis, P.L.; Ru, G. BSE surveillance in Italy: Neuropathological findings in cattle in the frame of the passive surveillance programme. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2003, 50, 48–49. [Google Scholar] [CrossRef]
- Jeffrey, M.; Wilesmith, J.W. Idiopathic brainstem neuronal chromatolysis and hippocampal sclerosis: A novel encephalopathy in clinically suspect cases of bovine spongiform encephalopathy. Vet. Rec. 1992, 131, 359–362. [Google Scholar] [CrossRef]
- Jeffrey, M. A neuropathological survey of brains submitted under the Bovine Spongiform Encephalopathy Orders in Scotland. Vet. Rec. 1992, 131, 332–337. [Google Scholar] [CrossRef]
- Miyashita, M.; Stierstorfer, B.; Schmahl, W. Neuropathological findings in brains of Bavarian cattle clinically suspected of bovine spongiform encephalopathy. J. Vet. Med. B Infect. Dis. Vet. Public Health 2004, 51, 209–215. [Google Scholar] [CrossRef]
- Munday, B.L.; Mason, R.W.; Cumming, R. Observations on diseases of the central nervous system of cattle in Tasmania. Aust. Vet. J. 1973, 49, 451–455. [Google Scholar] [CrossRef] [PubMed]
- McGill, I.S.; Wells, G.A. Neuropathological findings in cattle with clinically suspect but histologically unconfirmed bovine spongiform encephalopathy (BSE). J. Comp. Pathol. 1993, 108, 241–260. [Google Scholar] [CrossRef]
- Bouzalas, I.G.; Wuthrich, D.; Walland, J.; Drogemuller, C.; Zurbriggen, A.; Vandevelde, M.; Oevermann, A.; Bruggmann, R.; Seuberlich, T. Neurotropic astrovirus in cattle with nonsuppurative encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Seuberlich, T. Frequency and Pathological Phenotype of Bovine Astrovirus CH13/NeuroS1 Infection in Neurologically-Diseased Cattle: Towards Assessment of Causality. Viruses 2017, 9, 12. [Google Scholar] [CrossRef]
- Wuthrich, D.; Boujon, C.L.; Truchet, L.; Selimovic-Hamza, S.; Oevermann, A.; Bouzalas, I.G.; Bruggmann, R.; Seuberlich, T. Exploring the virome of cattle with non-suppurative encephalitis of unknown etiology by metagenomics. Virology 2016, 493, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Bouzalas, I.G.; Wuthrich, D.; Selimovic-Hamza, S.; Drogemuller, C.; Bruggmann, R.; Seuberlich, T. Full-genome based molecular characterization of encephalitis-associated bovine astroviruses. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 44, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Hull, R.N.; Minner, J.R.; Smith, J.W. New viral agents recovered from tissue cultures of monkey kidney cells. I. Origin and properties of cytopathogenic agents S.V.1, S.V.2, S.V.4, S.V.5, S.V.6, S.V.11, S.V.12 and S.V.15. Am. J. Hyg. 1956, 63, 204–215. [Google Scholar]
- Tribe, G.W. An investigation of the incidence, epidemiology and control of Simian virus 5. Br. J. Exp. Pathol. 1966, 47, 472–479. [Google Scholar]
- Atoynatan, T.; Hsiung, G.D. Epidemiologic studies of latent virus infections in captive monkeys and baboons. II. Serologic evidence of myxovirus infections with special reference to SV5. Am. J. Epidemiol. 1969, 89, 472–479. [Google Scholar] [CrossRef]
- Binn, L.N.; Eddy, G.A.; Lazar, E.C.; Helms, J.; Murnane, T. Viruses recovered from laboratory dogs with respiratory disease. Proc. Soc. Exp. Biol. Med. 1967, 126, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Appel, M.J.; Percy, D.H. SV-5-like parainfluenza virus in dogs. J. Am. Vet. Med. Assoc. 1970, 156, 1778–1781. [Google Scholar] [PubMed]
- McCandlish, I.A.; Thompson, H.; Cornwell, H.J.; Wright, N.G. A study of dogs with kennel cough. Vet. Rec. 1978, 102, 293–301. [Google Scholar] [CrossRef]
- Chatziandreou, N.; Stock, N.; Young, D.; Andrejeva, J.; Hagmaier, K.; McGeoch, D.J.; Randall, R.E. Relationships and host range of human, canine, simian and porcine isolates of simian virus 5 (parainfluenza virus 5). J. Gen. Virol. 2004, 85, 3007–3016. [Google Scholar] [CrossRef] [PubMed]
- Rima, B.K.; Gatherer, D.; Young, D.F.; Norsted, H.; Randall, R.E.; Davison, A.J. Stability of the parainfluenza virus 5 genome revealed by deep sequencing of strains isolated from different hosts and following passage in cell culture. J. Virol. 2014, 88, 3826–3836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erles, K.; Dubovi, E.J.; Brooks, H.W.; Brownlie, J. Longitudinal study of viruses associated with canine infectious respiratory disease. J. Clin. Microbiol. 2004, 42, 4524–4529. [Google Scholar] [CrossRef] [Green Version]
- ICTVonline. Available online: https://talk.ictvonline.org//taxonomy/p/taxonomy-history?taxnode_id=201851632 (accessed on 13 November 2019).
- Evermann, J.F.; Lincoln, J.D.; McKiernan, A.J. Isolation of a paramyxovirus from the cerebrospinal fluid of a dog with posterior paresis. J. Am. Vet. Med. Assoc. 1980, 177, 1132–1134. [Google Scholar]
- Baumgartner, W.K.; Metzler, A.E.; Krakowka, S.; Koestner, A. In vitro identification and characterization of a virus isolated from a dog with neurological dysfunction. Infect. Immun. 1981, 31, 1177–1183. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, W.K.; Krakowka, S.; Koestner, A.; Evermann, J. Acute encephalitis and hydrocephalus in dogs caused by canine parainfluenza virus. Vet. Pathol. 1982, 19, 79–92. [Google Scholar] [CrossRef]
- Liu, Y.; Li, N.; Zhang, S.; Zhang, F.; Lian, H.; Hu, R. Parainfluenza Virus 5 as Possible Cause of Severe Respiratory Disease in Calves, China. Emerg. Infect. Dis. 2015, 21, 2242–2244. [Google Scholar] [CrossRef]
- Yang, D.K.; Nah, J.J.; Kim, H.H.; Choi, S.S.; Bae, Y.C.; Park, J.W.; Song, J.Y. Isolation of novel bovine parainfluenza virus type 5 (bPIV5) and its incidence in Korean cattle. Korean J. Vet. Res. 2014, 54, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Luthi, R.; Boujon, C.L.; Kauer, R.; Koch, M.C.; Bouzalas, I.G.; Seuberlich, T. Accurate and precise real-time RT-PCR assays for the identification of astrovirus associated encephalitis in cattle. Sci. Rep. 2018, 8, 9215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boujon, C.L.; Selimovic-Hamza, S.; Bouzalas, I.; Seuberlich, T. Development and validation of an immunohistochemistry procedure for the detection of a neurotropic bovine astrovirus. J. Virol. Methods 2017, 239, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolakofsky, D.; Pelet, T.; Garcin, D.; Hausmann, S.; Curran, J.; Roux, L. Paramyxovirus RNA synthesis and the requirement for hexamer genome length: The rule of six revisited. J. Virol. 1998, 72, 891–899. [Google Scholar] [CrossRef] [Green Version]
- He, B.; Lin, G.Y.; Durbin, J.E.; Durbin, R.K.; Lamb, R.A. The SH integral membrane protein of the paramyxovirus simian virus 5 is required to block apoptosis in MDBK cells. J. Virol. 2001, 75, 4068–4079. [Google Scholar] [CrossRef] [Green Version]
- Yuan, P.; Thompson, T.B.; Wurzburg, B.A.; Paterson, R.G.; Lamb, R.A.; Jardetzky, T.S. Structural studies of the parainfluenza virus 5 hemagglutinin-neuraminidase tetramer in complex with its receptor, sialyllactose. Structure 2005, 13, 803–815. [Google Scholar] [CrossRef] [Green Version]
- Kubota, M.; Takeuchi, K.; Watanabe, S.; Ohno, S.; Matsuoka, R.; Kohda, D.; Nakakita, S.I.; Hiramatsu, H.; Suzuki, Y.; Nakayama, T.; et al. Trisaccharide containing alpha2,3-linked sialic acid is a receptor for mumps virus. Proc. Natl. Acad. Sci. USA 2016, 113, 11579–11584. [Google Scholar] [CrossRef] [Green Version]
- Chatziandreou, N.; Young, D.; Andrejeva, J.; Goodbourn, S.; Randall, R.E. Differences in interferon sensitivity and biological properties of two related isolates of simian virus 5: A model for virus persistence. Virology 2002, 293, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, W.; Krakowka, S.; Durchfeld, B. In vitro cytopathogenicity and in vivo virulence of two strains of canine parainfluenza virus. Vet. Pathol. 1991, 28, 324–331. [Google Scholar] [CrossRef]
- Young, D.F.; Carlos, T.S.; Hagmaier, K.; Fan, L.; Randall, R.E. AGS and other tissue culture cells can unknowingly be persistently infected with PIV5; a virus that blocks interferon signalling by degrading STAT1. Virology 2007, 365, 238–240. [Google Scholar] [CrossRef] [Green Version]
- Zakstelskaya, L.Y.; Zhdanov, V.M.; Yakhno, M.A.; Gushchin, B.V.; Klimenko, S.M.; Demidova, S.A.; Konovalova, N.G.; Gushchina, E.A. Persistent SV5 virus infection in continuous cell cultures. Acta Virol. 1976, 20, 506–511. [Google Scholar] [PubMed]
- Goswami, K.K.; Cameron, K.R.; Russell, W.C.; Lange, L.S.; Mitchell, D.N. Evidence for the persistence of paramyxoviruses in human bone marrows. J. Gen. Virol. 1984, 65, 1881–1888. [Google Scholar] [CrossRef] [PubMed]
- Rovnak, J.; Quackenbush, S.L.; Reyes, R.A.; Baines, J.D.; Parrish, C.R.; Casey, J.W. Detection of a novel bovine lymphotropic herpesvirus. J. Virol. 1998, 72, 4237–4242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubis, P.; Materniak, M.; Kuzmak, J. Comparison of nested PCR and qPCR for the detection and quantitation of BoHV6 DNA. J. Virol. Methods 2013, 194, 94–101. [Google Scholar] [CrossRef]
- Rivers, T.M. Viruses and Koch’s Postulates. J. Bacteriol. 1937, 33, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lipkin, W.I. The changing face of pathogen discovery and surveillance. Nat. Rev. Microbiol. 2013, 11, 133–141. [Google Scholar] [CrossRef]
- Schulz, B.S.; Kurz, S.; Weber, K.; Balzer, H.J.; Hartmann, K. Detection of respiratory viruses and Bordetella bronchiseptica in dogs with acute respiratory tract infections. Vet. J. 2014, 201, 365–369. [Google Scholar] [CrossRef]
- Mitchell, J.A.; Cardwell, J.M.; Leach, H.; Walker, C.A.; Le Poder, S.; Decaro, N.; Rusvai, M.; Egberink, H.; Rottier, P.; Fernandez, M.; et al. European surveillance of emerging pathogens associated with canine infectious respiratory disease. Vet. Microbiol. 2017, 212, 31–38. [Google Scholar] [CrossRef]
- Zhang, W.; Li, L.; Deng, X.; Kapusinszky, B.; Delwart, E. What is for dinner? Viral metagenomics of US store bought beef, pork, and chicken. Virology 2014, 468–470, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Endemo. Available online: http://endmemo.com/bio/dnacopynum.php (accessed on 9 March 2019).
- Toussaint, J.F.; Sailleau, C.; Breard, E.; Zientara, S.; De Clercq, K. Bluetongue virus detection by two real-time RT-qPCRs targeting two different genomic segments. J. Virol. Methods 2007, 140, 115–123. [Google Scholar] [CrossRef]
- Bottero, M.T.; Civera, T.; Nucera, D.; Rosati, S.; Sacchi, P.; Turi, R.M. A multiplex polymerase chain reaction for the identification of cows’, goats’ and sheep’s milk in dairy products. Int. Dairy J. 2003, 13, 277–282. [Google Scholar] [CrossRef]
- Li, Z.; Yu, M.; Zhang, H.; Wang, H.Y.; Wang, L.F. Improved rapid amplification of cDNA ends (RACE) for mapping both the 5′ and 3′ terminal sequences of paramyxovirus genomes. J. Virol. Methods 2005, 130, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| History | 26731 | Animal 26875 | 27020 |
|---|---|---|---|
| Date of receipt | 27 February 1998 | 28 April 1998 | 10 July 1998 |
| Age (years) at time of death | 5 | 2 | 3 |
| Reported neuro-logicalsymptoms | stall walking | peracute: tremor of head and neck, reduced vision, hypersensitivity in hind limbs | neurological symptoms for 3 weeks |
| Histological examination at time of admission | - meningo-encephalitis - mononuclear, perivascular infiltrations - gliosis - neuronal necrosis - location: basal ganglia, diencephalon and mesencephalon | - meningo-encephalitis - mononuclear infiltrations (and mononuclear meningitis) - gliosis - neuronal necrosis - location: cerebellum and brainstem | - meningo-encephalitis - mononuclear infiltrations (and mononuclear meningitis) - gliosis - location: cortex, brainstem (especially mesencephalon) and cerebellum |
| Studies PIV5 was detected | Wüthrich et al., Virology (2016) 1 | Bouzalas et al., Infect Genet Evol (2016) 2 | Wüthrich et al., Virology (2016) 1 |
| Next Generation Sequencing results for PIV5 | + 1 low read depth and low coverage | + 2 high read depth and nearly complete coverage | - 1 |
| RT-PCR results for PIV5 | + 1 | not performed | + 1 |
| Other viruses found in the brain (method) | BoHV6 (qPCR 1), BoPyV2 (PCR 1) | BoAstV-CH13 (RT-PCR 3, NGS2, ISH 3, IHC 4, RT-qPCR 5) | BoHV6 (qPCR 1), BoPyV2 (PCR1), BoAstV CH13 (ISH 6, RT-qPCR 5) |
| Virus | 26731 | Animal 26875 | 27020 |
|---|---|---|---|
| PIV5 A | 4.09 × 104 | 1.87 × 102 | 1.11 × 101 |
| BoAstV CH13 A | - | 5.74 × 105 | 5.14 × 102 |
| BoHV6 B | 2.79 × 102 | - | 1.58 × 102 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hierweger, M.M.; Werder, S.; Seuberlich, T. Parainfluenza Virus 5 Infection in Neurological Disease and Encephalitis of Cattle. Int. J. Mol. Sci. 2020, 21, 498. https://doi.org/10.3390/ijms21020498
Hierweger MM, Werder S, Seuberlich T. Parainfluenza Virus 5 Infection in Neurological Disease and Encephalitis of Cattle. International Journal of Molecular Sciences. 2020; 21(2):498. https://doi.org/10.3390/ijms21020498
Chicago/Turabian StyleHierweger, Melanie M., Simea Werder, and Torsten Seuberlich. 2020. "Parainfluenza Virus 5 Infection in Neurological Disease and Encephalitis of Cattle" International Journal of Molecular Sciences 21, no. 2: 498. https://doi.org/10.3390/ijms21020498
APA StyleHierweger, M. M., Werder, S., & Seuberlich, T. (2020). Parainfluenza Virus 5 Infection in Neurological Disease and Encephalitis of Cattle. International Journal of Molecular Sciences, 21(2), 498. https://doi.org/10.3390/ijms21020498