Deimination and Peptidylarginine Deiminases in Skin Physiology and Diseases


Abstract
:1. Introduction
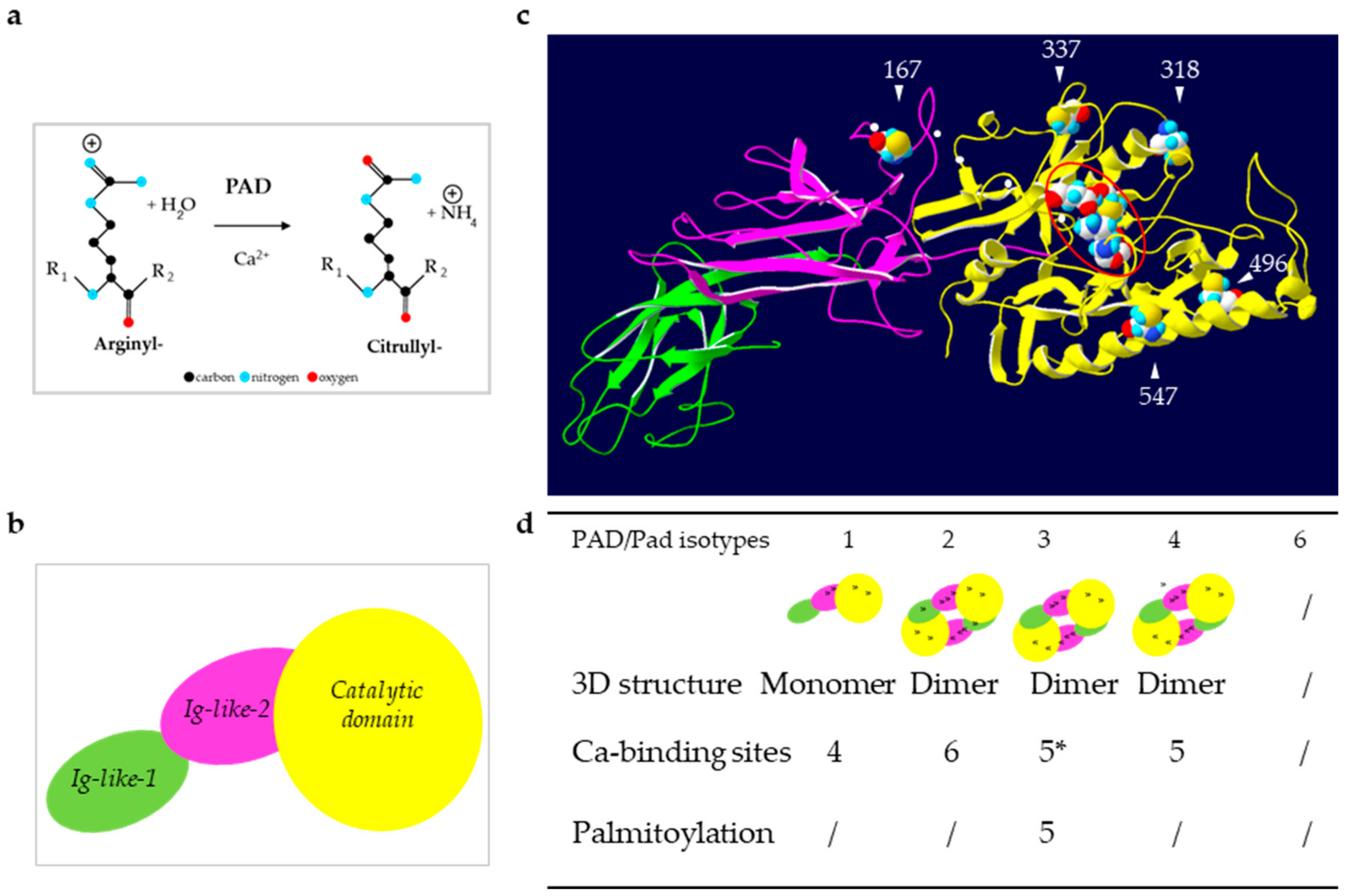
2. Expression and Role of PADs in Normal Skin
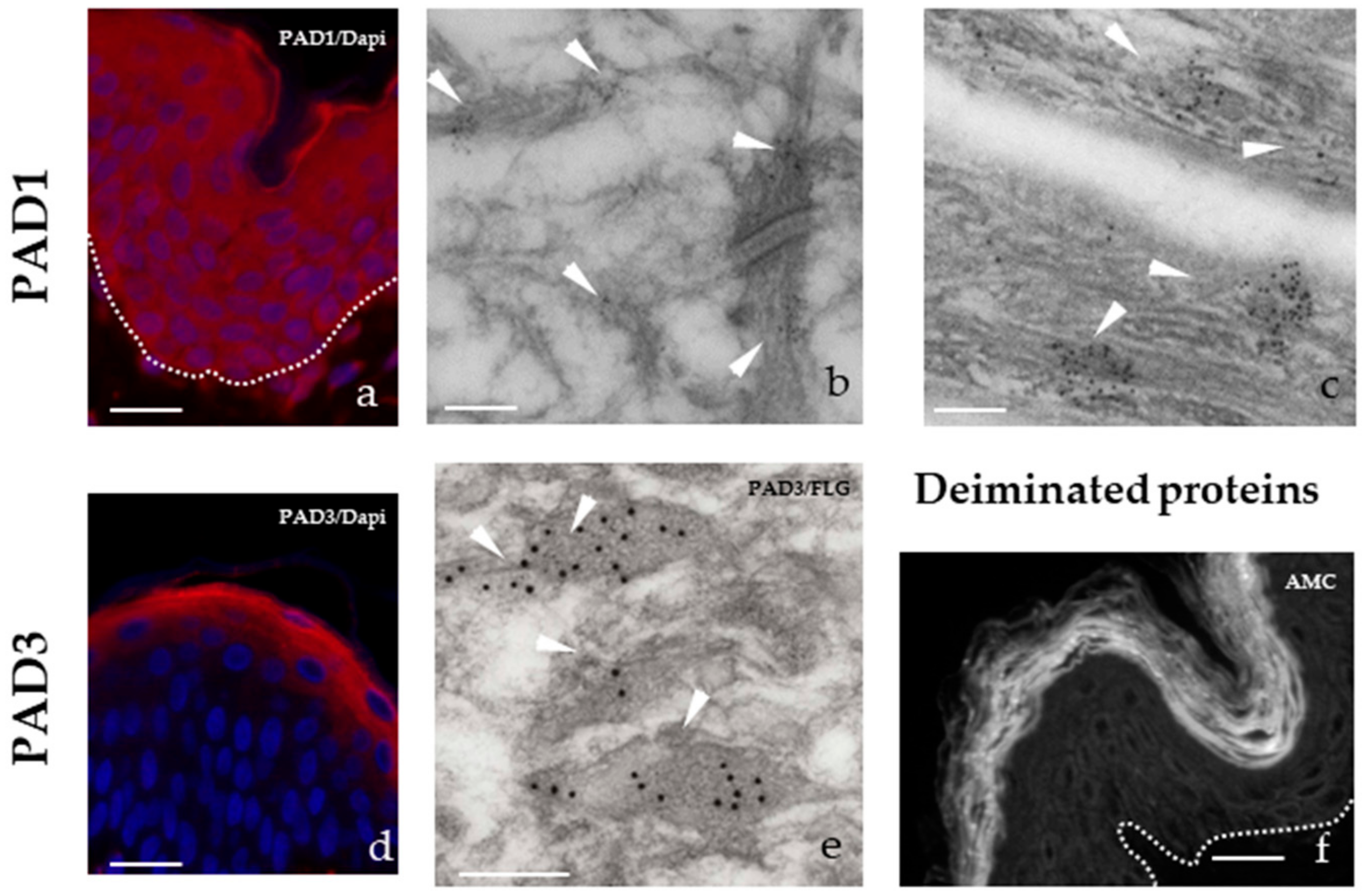
2.1. PADs and Deiminated Proteins in the Epidermis
2.2. PADs and Deiminated Proteins in Hair Follicles
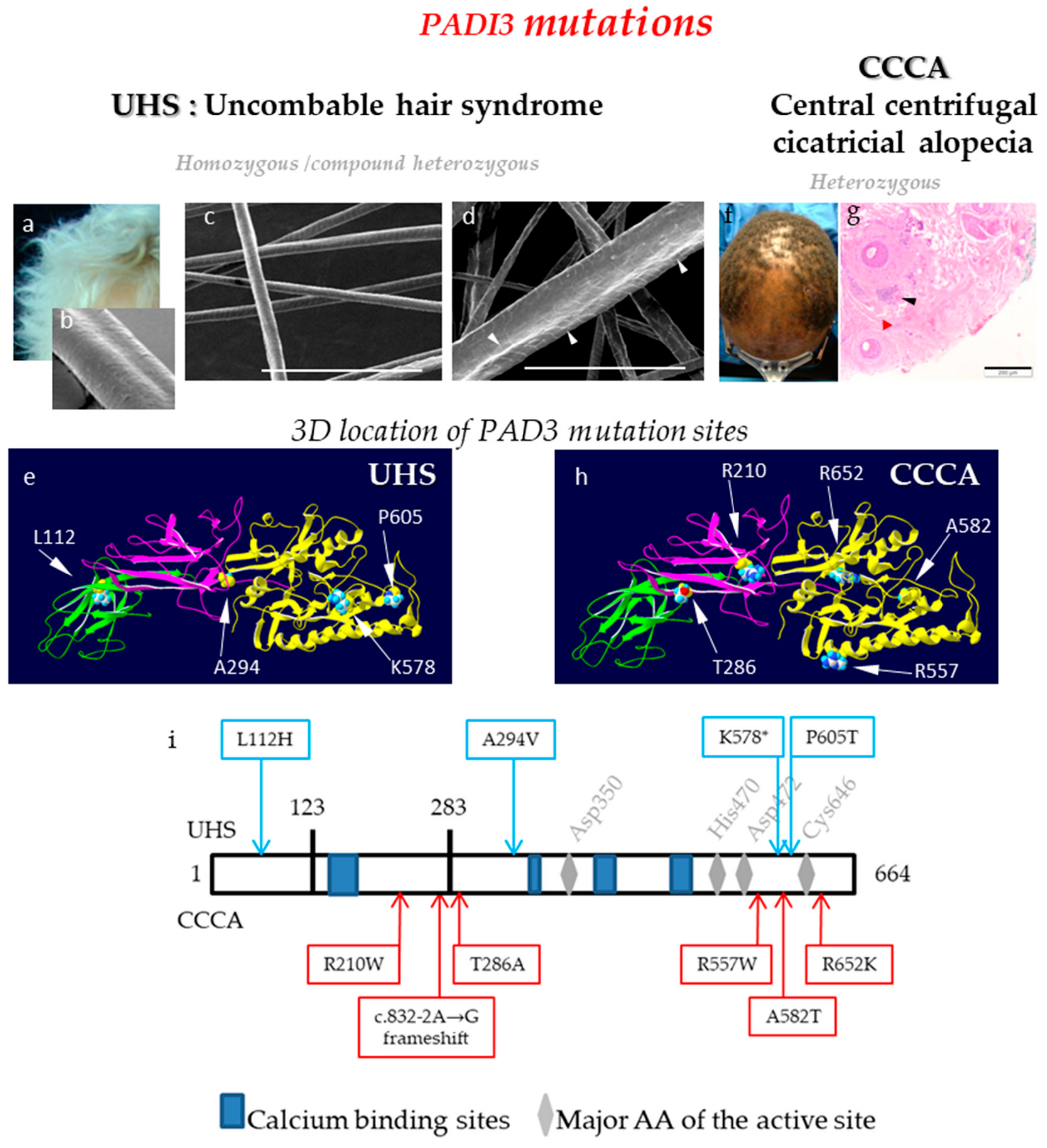
3. PAD Defects and Skin Diseases
4. Conclusions
- In skin, PADs are regulated at multiple levels, including gene expression, mRNA translation, and post-translational modifications, such as autodeimination and S-palmitoylation;
- Cornified envelopes contain deiminated proteins, including filaggrin, hornerin, and the newly identified keratinocyte proline rich protein, deimination improving the efficacy of cross-linking by transglutaminases;
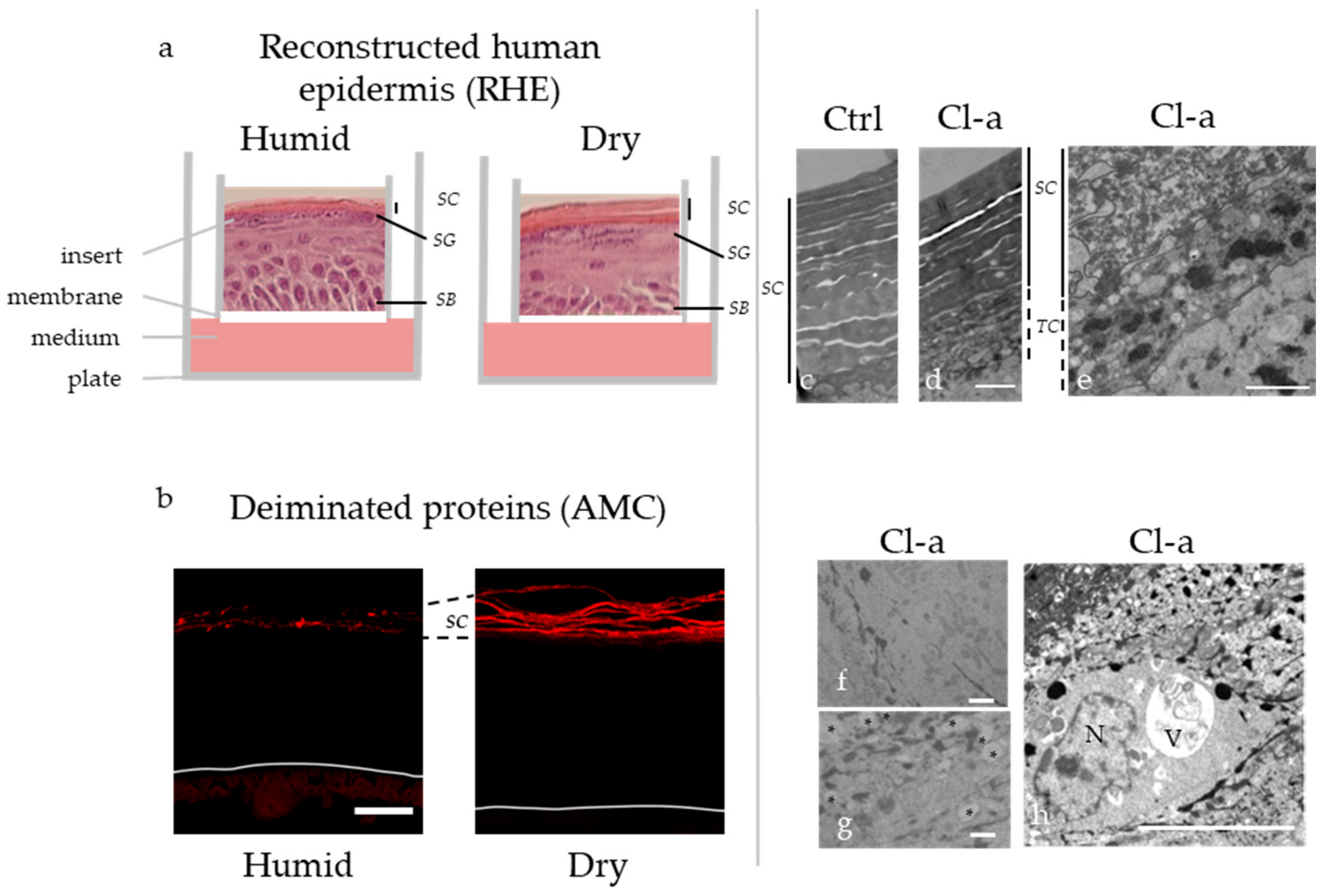
- PAD inhibition using chloro-amidine slows down cornification and alters the associated autophagy process in the granular keratinocytes, showing the importance of these enzymes in the last steps of keratinocyte differentiation;
- As demonstrated by the comparison of reconstructed human epidermis produced in dry versus humid atmospheric conditions, keratinocyte external environment modifies PAD1 expression and deimination, and the subsequent filaggrin proteolysis;
- The direct implication of PAD3 gene mutations has been demonstrated in two human hair disorders, a rare hair shaft dysplasia and a common scarring inflammatory alopecia.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CCCA | Central Centrifugal Cicatricial Alopecia |
| NMF | Natural Moisturizing Factor |
| PAD | peptidylarginine deiminase |
| TGase | transglutaminase |
| UHS | Uncombable Hair Syndrome |
References
- Chavanas, S.; Méchin, M.C.; Takahara, H.; Kawada, A.; Nachat, R.; Serre, G.; Simon, M. Comparative analysis of the mouse and human peptidylarginine deiminase gene clusters reveals highly conserved non-coding segments and a new human gene, PADI6. Gene 2004, 330, 19–27. [Google Scholar] [CrossRef]
- Shimizu, A.; Handa, K.; Honda, T.; Abe, N.; Kojima, T.; Takahara, H. Three isozymes of peptidylarginine deiminase in the chicken: Molecular cloning, characterization, and tissue distribution. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2014, 167, 65–73. [Google Scholar] [CrossRef]
- Minato, T.; Unno, M.; Kitano, T. Evolution of S100A3 and PAD3, two important genes for mammalian hair. Gene 2019, 713, 143975. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Y. Peptidylarginine deiminases in citrullination, gene regulation, health and pathogenesis. Biochim. Biophys. Acta 2013, 1829, 1126–1135. [Google Scholar] [CrossRef] [Green Version]
- Christophorou, M.A.; Castelo-Branco, G.; Halley-Stott, R.P.; Oliveira, C.S.; Loos, R.; Radzisheuskaya, A.; Mowen, K.A.; Bertone, P.; Silva, J.C.; Zernicka-Goetz, M.; et al. Citrullination regulates pluripotency and histone H1 binding to chromatin. Nature 2014, 507, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Witalison, E.E.; Thompson, P.R.; Hofseth, L.J. Protein Arginine Deiminases and Associated Citrullination: Physiological Functions and Diseases Associated with Dysregulation. Curr. Drug Targets 2015, 16, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, O.E.; Borregaard, N. Neutrophil extracellular traps—The dark side of neutrophils. J. Clin. Invest. 2016, 126, 1612–1620. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Tan, D.; Piao, H. Myelin Basic Protein Citrullination in Multiple Sclerosis: A Potential Therapeutic Target for the Pathology. Neurochem. Res. 2016, 41, 1845–1856. [Google Scholar] [CrossRef] [PubMed]
- Arita, K.; Hashimoto, H.; Shimizu, T.; Nakashima, K.; Yamada, M.; Sato, M. Structural basis for Ca2+-induced activation of human PAD4. Nat. Struct. Mol. Biol. 2004, 11, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Slade, D.J.; Fang, P.; Dreyton, C.J.; Zhang, Y.; Fuhrmann, J.; Rempel, D.; Bax, B.D.; Coonrod, S.A.; Lewis, H.D.; Guo, M.; et al. Protein arginine deiminase 2 binds calcium in an ordered fashion: Implications for inhibitor design. ACS Chem. Biol. 2015, 10, 1043–1053. [Google Scholar] [CrossRef] [Green Version]
- Saijo, S.; Nagai, A.; Kinjo, S.; Mashimo, R.; Akimoto, M.; Kizawa, K.; Yabe-Wada, T.; Shimizu, N.; Takahara, H.; Unno, M. Monomeric Form of Peptidylarginine Deiminase Type I Revealed by X-ray Crystallography and Small-Angle X-ray Scattering. J. Mol. Biol. 2016, 428, 3058–3073. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, M.; Al Ghamdi, K.A.; Khan, R.H.; Uversky, V.N.; Redwan, E.M. An interplay of structure and intrinsic disorder in the functionality of peptidylarginine deiminases, a family of key autoimmunity-related enzymes. Cell. Mol. Life Sci. 2019, 76, 4635–4662. [Google Scholar] [CrossRef]
- Méchin, M.-C.; Sebbag, M.; Arnaud, J.; Nachat, R.; Foulquier, C.; Adoue, V.; Coudane, F.; Duplan, H.; Schmitt, A.-M.; Chavanas, S.; et al. Update on peptidylarginine deiminases and deimination in skin physiology and severe human diseases. Int. J. Cosmet. Sci. 2007, 29, 147–168. [Google Scholar] [CrossRef]
- Liu, Y.L.; Lee, C.Y.; Huang, Y.N.; Chen, H.Y.; Liu, G.Y.; Hung, H.C. Probing the Roles of Calcium-Binding Sites during the Folding of Human Peptidylarginine Deiminase 4. Sci. Rep. 2017, 7, 2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuthbert, G.L.; Daujat, S.; Snowden, A.W.; Erdjument-Bromage, H.; Hagiwara, T.; Yamada, M.; Schneider, R.; Gregory, P.D.; Tempst, P.; Bannister, A.J.; et al. Histone deimination antagonizes arginine methylation. Cell 2004, 118, 545–553. [Google Scholar] [CrossRef] [Green Version]
- Méchin, M.C.; Coudane, F.; Adoue, V.; Arnaud, J.; Duplan, H.; Charveron, M.; Schmitt, A.M.; Takahara, H.; Serre, G.; Simon, M. Deimination is regulated at multiple levels including auto-deimination of peptidylarginine deiminases. Cell. Mol. Life Sci. 2010, 67, 1491–1503. [Google Scholar] [CrossRef]
- Agraz-Cibrian, J.M.; Giraldo, D.M.; Urcuqui-Inchima, S. 1,25-Dihydroxyvitamin D3 induces formation of neutrophil extracellular trap-like structures and modulates the transcription of genes whose products are neutrophil extracellular trap-associated proteins: A pilot study. Steroids 2019, 141, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Andrade, F.; Darrah, E.; Gucek, M.; Cole, R.N.; Rosen, A.; Zhu, X. Autocitrullination of human peptidyl arginine deiminase type 4 regulates protein citrullination during cell activation. Arthritis Rheum. 2010, 62, 1630–1640. [Google Scholar] [CrossRef] [Green Version]
- Slack, J.L.; Jones, L.E., Jr.; Bhatia, M.M.; Thompson, P.R. Autodeimination of protein arginine deiminase 4 alters protein-protein interactions but not activity. Biochemistry 2011, 50, 3997–4010. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.H.; Dwivedi, N.; Nicholas, A.P.; Ho, I.C. The W620 Polymorphism in PTPN22 Disrupts Its Interaction With Peptidylarginine Deiminase Type 4 and Enhances Citrullination and NETosis. Arthritis Rheumatol. 2015, 67, 2323–2334. [Google Scholar] [CrossRef]
- Chen, L.Y.; Lin, K.R.; Chen, Y.J.; Chiang, Y.J.; Ho, K.C.; Shen, L.F.; Song, I.W.; Liu, K.M.; Yang-Yen, H.F.; Chen, Y.J.; et al. Palmitoyl acyltransferase activity of ZDHHC13 regulates skin barrier development partly by controlling PADi3 and TGM1 protein stability. J. Invest. Dermatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Basmanav, Ü.F.B.; Cau, L.; Tafazzoli, A.; Méchin, M.C.; Wolf, S.; Romano, M.T.; Valentin, F.; Wiegmann, H.; Huchenq, A.; Kandil, R.; et al. Mutations in Three Genes Encoding Proteins Involved in Hair Shaft Formation Cause Uncombable Hair Syndrome. Am. J. Hum. Genet. 2016, 99, 1292–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malki, L.; Sarig, O.; Romano, M.T.; Méchin, M.C.; Peled, A.; Pavlovsky, M.; Warshauer, E.; Samuelov, L.; Uwakwe, L.; Briskin, V.; et al. Variant PADI3 in Central Centrifugal Cicatricial Alopecia. N. Engl. J. Med. 2019, 380, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Guerrin, M.; Ishigami, A.; Méchin, M.C.; Nachat, R.; Valmary, S.; Sebbag, M.; Simon, M.; Senshu, T.; Serre, G. cDNA cloning, gene organization and expression analysis of human peptidylarginine deiminase type I. Biochem. J. 2003, 370, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Nachat, R.; Méchin, M.-C.; Takahara, H.; Chavanas, S.; Charveron, M.; Serre, G.; Simon, M. peptidylarginine deiminase isoforms 1–3 are expressed in the epidermis and involved in the deimination of K1 and filaggrin. J. Invest. Dermatol. 2005, 124, 384–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachat, R.; Méchin, M.C.; Charveron, M.; Serre, G.; Constans, J.; Simon, M. Peptidylarginine deiminase isoforms are differentially expressed in the anagen hair follicles and other human skin appendages. J. Invest. Dermatol. 2005, 125, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Kizawa, K.; Takahara, H.; Unno, M.; Heizmann, C.W. S100 and S100 fused-type protein families in epidermal maturation with special focus on S100A3 in mammalian hair cuticles. Biochimie 2011, 93, 2038–2047. [Google Scholar] [CrossRef]
- Yamakoshi, A.; Ono, H.; Nishijyo, T.; Shiraiwa, M.; Takahara, H. Cloning of cDNA encoding a novel isoform (type IV) of peptidylarginine deiminase from rat epidermis. Biochim. Biophys. Acta 1998, 1386, 227–232. [Google Scholar] [CrossRef]
- Senshu, T.; Kan, S.; Ogawa, H.; Manabe, M.; Asaga, H. Preferential Deimination of Keratin K1 and Filaggrin during the Terminal Differentiation of Human Epidermis. Biochem. Biophys. Res. Commun. 1996, 225, 712–719. [Google Scholar] [CrossRef]
- Harding, C.R.; Scott, I.R. Histidine-rich proteins (filaggrins): Structural and functional heterogeneity during epidermal differentiation. J. Mol. Biol. 1983, 170, 651–673. [Google Scholar] [CrossRef]
- Tarcsa, E.; Marekov, L.N.; Mei, G.; Melino, G.; Lee, S.-C.; Steinert, P.M. Protein unfolding by peptidylarginine deiminase substrate specificity and structural relationships of the natural substrates trichohyalin and filaggrin. J. Biol. Chem. 1996, 271, 30709–30716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.-Y.; Gasc, G.; Raymond, A.-A.; Burlet-Schiltz, O.; Takahara, H.; Serre, G.; Mechin, M.-C.; Simon, M. Deimination of human hornerin enhances its processing by calpain-1 and its cross-linking by transglutaminases. J. Invest. Dermatol 2017, 137, 422–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.-Y.; Henry, J.; Raymond, A.-A.; Méchin, M.-C.; Pendaries, V.; Nassar, D.; Hansmann, B.; Balica, S.; Burlet-Schiltz, O.; Schmitt, A.M.; et al. Deimination of human filaggrin-2 promotes its proteolysis by calpain 1. J. Biol. Chem. 2011, 286, 23222–23233. [Google Scholar] [CrossRef] [Green Version]
- Henry, J.; Toulza, E.; Hsu, C.Y.; Pellerin, L.; Balica, S.; Mazereeuw-Hautier, J.; Paul, C.; Serre, G.; Jonca, N.; Simon, M. Update on the epidermal differentiation complex. Front. Biosci. (Landmark Ed.) 2012, 17, 1517–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albérola, G.; Schröder, J.M.; Froment, C.; Simon, M. The Amino-Terminal Part of Human FLG2 Is a Component of Cornified Envelopes. J. Invest. Dermatol. 2019, 139, 1395–1397. [Google Scholar] [CrossRef] [PubMed]
- Senshu, T.; Akiyama, K.; Nomura, K. Identification of citrulline residues in the V subdomains of keratin K1 derived from the cornified layer of newborn mouse epidermis. Exp. Dermatol. 1999, 8, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Pendaries, V.; Malaisse, J.; Pellerin, L.; Le Lamer, M.; Nachat, R.; Kezic, S.; Schmitt, A.M.; Paul, C.; Poumay, Y.; Serre, G.; et al. Knockdown of filaggrin in a three-dimensional reconstructed human epidermis impairs keratinocyte differentiation. J. Invest. Dermatol. 2014, 134, 2938–2946. [Google Scholar] [CrossRef] [Green Version]
- Le Lamer, M.; Pellerin, L.; Reynier, M.; Cau, L.; Pendaries, V.; Leprince, C.; Méchin, M.C.; Serre, G.; Paul, C.; Simon, M. Defects of corneocyte structural proteins and epidermal barrier in atopic dermatitis. Biol. Chem. 2015, 396, 1163–1179. [Google Scholar] [CrossRef]
- Pendaries, V.; Le Lamer, M.; Cau, L.; Hansmann, B.; Malaisse, J.; Kezic, S.; Serre, G.; Simon, M. In a three-dimensional reconstructed human epidermis filaggrin-2 is essential for proper cornification. Cell Death Dis. 2015, 6, e1656. [Google Scholar] [CrossRef] [Green Version]
- Kamata, Y.; Taniguchi, A.; Yamamoto, M.; Nomura, J.; Ishihara, K.; Takahara, H.; Hibino, T.; Takeda, A. Neutral cysteine protease bleomycin hydrolase is essential for the breakdown of deiminated filaggrin into amino acids. J. Biol. Chem. 2009, 284, 12829–12836. [Google Scholar] [CrossRef] [Green Version]
- Cau, L.; Méchin, M.C.; Simon, M. Peptidylarginine deiminases and deiminated proteins at the epidermal barrier. Exp. Dermatol. 2018, 27, 852–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cau, L.; Pendaries, V.; Lhuillier, E.; Thompson, P.R.; Serre, G.; Takahara, H.; Méchin, M.-C.; Simon, M. Lowering relative humidity level increases epidermal protein deimination and drives human filaggrin breakdown. J. Dermatol. Sci. 2017, 86, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arita, S.; Hatta, M.; Uchida, K.; Kita, T.; Okamura, K.; Ryu, T.; Murakami, H.; Sakagami, R.; Yamazaki, J. Peptidylarginine deiminase is involved in maintaining the cornified oral mucosa of rats. J. Periodontal Res. 2018, 53, 750–761. [Google Scholar] [CrossRef] [PubMed]
- Méchin, M.-C.; Cau, L.; Galliano, M.F.; Daunes-Marion, S.; Poigny, S.; Vidaluc, J.L.; Bessou-Touya, S.; Takahara, H.; Serre, G.; Duplan, H.; et al. Acefylline activates filaggrin deimination by peptidylarginine deiminases in the upper epidermis. J. Dermatol. Sci. 2016, 81, 101–106. [Google Scholar] [CrossRef]
- Akiyama, K.; Senshu, T. Dynamic aspects of protein deimination in developing mouse epidermis. Exp. Dermatol. 1999, 8, 177–186. [Google Scholar] [CrossRef]
- Scott, I.R.; Harding, C.R. Filaggrin breakdown to water binding compounds during development of the rat stratum corneum is controlled by the water activity of the environment. Dev. Biol. 1986, 115, 84–92. [Google Scholar] [CrossRef]
- Cau, L.; Takahara, H.; Thompson, P.R.; Serre, G.; Méchin, M.-C.; Simon, M. Peptidylarginine deiminase inhibitor cl-amidine attenuates cornification and interferes with the regulation of autophagy in reconstructed human epidermis. J. Invest. Dermatol. 2019, 139, 1889–1897.e4. [Google Scholar] [CrossRef]
- Wang, S.; Chen, X.A.; Hu, J.; Jiang, J.; Li, Y.; Chan-Salis, K.Y.; Gu, Y.; Chen, G.; Thomas, C.; Pugh, B.F.; et al. ATF4 Gene Network Mediates Cellular Response to the Anticancer PAD Inhibitor YW3-56 in Triple-Negative Breast Cancer Cells. Mol. Cancer Ther. 1995, 14, 877–888. [Google Scholar] [CrossRef] [Green Version]
- Sorice, M.; Iannuccelli, C.; Manganelli, V.; Capozzi, A.; Alessandri, C.; Lococo, E.; Garofalo, T.; Di Franco, M.; Bombardieri, M.; Nerviani, A.; et al. Autophagy generates citrullinated peptides in human synoviocytes: A possible trigger for anti-citrullinated peptide antibodies. Rheumatology (Oxford) 2016, 55, 1374–1385. [Google Scholar] [CrossRef] [Green Version]
- Coudane, F.; Méchin, M.-C.; Huchenq, A.; Henry, J.; Nachat, R.; Ishigami, A.; Adoue, V.; Sebbag, M.; Serre, G.; Simon, M. Deimination and expression of peptidylarginine deiminases during cutaneous wound healing in mice. Eur. J. Dermatol. 2011, 21, 376–384. [Google Scholar] [CrossRef]
- Medland, S.E.; Nyholt, D.R.; Painter, J.N.; McEvoy, B.P.; McRae, A.F.; Zhu, G.; Gordon, S.D.; Ferreira, M.A.R.; Wright, M.J.; Henders, A.K.; et al. Common Variants in the Trichohyalin Gene Are Associated with Straight Hair in Europeans. Am. J. Hum. Genet. 2009, 85, 750–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pośpiech, E.; Karłowska-Pik, J.; Marcińska, M.; Abidi, S.; Andersen, J.D.; Berge, M.V.D.; Carracedo, Á.; Eduardoff, M.; Freire-Aradas, A.; Morling, N.; et al. Evaluation of the predictive capacity of DNA variants associated with straight hair in Europeans. Forensic Sci. Int. Genet. 2015, 19, 280–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, R.H.; Rocke, D.M.; Tsai, H.S.; Silva, K.A.; Lee, Y.J.; Sundberg, J.P. Distinguishing mouse strains by proteomic analysis of pelage hair. J. Invest. Dermatol. 2009, 129, 2120–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinert, P.M.; Parry, D.A.D.; Marekov, L.N. Trichohyalin mechanically strengthens the hair follicle: Multiple cross-bridging roles in the inner root sheath. J. Biol. Chem. 2003, 278, 41409–41419. [Google Scholar] [CrossRef] [PubMed]
- Kizawa, K.; Fujimori, T.; Kawai, T. Arachidonate 12-Lipoxygenase Inhibitors Promote S100A3 Citrullination in Cultured SW480 Cells and Isolated Hair Follicles. Biol. Pharm. Bull. 2017, 40, 516–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unno, M.; Kawasaki, T.; Takahara, H.; Heizmann, C.W.; Kizawa, K. Refined crystal structures of human Ca2+/Zn2+-binding S100A3 protein characterized by two disulfide bridges. J. Mol. Biol. 2011, 408, 477–490. [Google Scholar] [CrossRef]
- Ishida-Yamamoto, A.; Senshu, T.; Takahashi, H.; Akiyama, K.; Nomura, K.; Iizuka, H. Decreased deiminated keratin K1 in psoriatic hyperproliferative epidermis. J. Invest. Dermatol. 2000, 114, 701–705. [Google Scholar] [CrossRef] [Green Version]
- Ishida-Yamamoto, A.; Senshu, T.; Eady, R.A.; Takahashi, H.; Shimizu, H.; Akiyama, M.; Iizuka, H. Sequential reorganization of cornified cell keratin filaments involving filaggrin-mediated compaction and keratin 1 deimination. J. Invest. Dermatol. 2002, 118, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Daunes-Marion, S.; Duplan, H.; Méchin, M.-C.; Poigny, S.; Serre, G.; Simon, M. Peptidylarginine Deiminase 1 and/or 3 Activator Compounds in the Epidermis and Uses Thereof. U.S. Patent 9566222, 14 February 2011. [Google Scholar]
- Urano, Y.; Watanabe, K.; Sakaki, A.; Arase, S.; Watanabe, Y.; Shigemi, F.; Takeda, K.; Akiyama, K.; Senshu, T. Immunohistochemical demonstration of peptidylarginine deiminase in human sweat glands. Am. J. Dermatopathol. 1990, 12, 249–255. [Google Scholar] [CrossRef]
- Chang, X.; Han, J.; Pang, L.; Zhao, Y.; Yang, Y.; Shen, Z. Increased PADI4 expression in blood and tissues of patients with malignant tumors. BMC Cancer 2009, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Wang, D.; Yao, H.; Doret, P.; Hao, G.; Shen, Q.; Qiu, H.; Zhang, X.; Wang, Y.; Chen, G.; et al. Coordination of PAD4 and HDAC2 in the regulation of p53-target gene expression. Oncogene 2010, 29, 3153–3162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, H.; Li, P.; Venters, B.J.; Zheng, S.; Thompson, P.R.; Pugh, B.F.; Wang, Y. Histone Arg modifications and p53 regulate the expression of OKL38, a mediator of apoptosis. J. Biol. Chem. 2008, 283, 20060–20068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stacey, S.N.; Gudbjartsson, D.F.; Sulem, P.; Bergthorsson, J.T.; Kumar, R.; Thorleifsson, G.; Sigurdsson, A.; Jakobsdottir, M.; Sigurgeirsson, B.; Benediktsdottir, K.R.; et al. Common variants on 1p36 and 1q42 are associated with cutaneous basal cell carcinoma but not with melanoma or pigmentation traits. Nat. Genet. 2008, 40, 1313–1318. [Google Scholar] [CrossRef] [PubMed]
- Bognar, P.; Nemeth, I.; Mayer, B.; Haluszka, D.; Wikonkal, N.; Ostorhazi, E.; John, S.; Paulsson, M.; Smyth, N.; Pasztoi, M.; et al. Reduced inflammatory threshold indicates skin barrier defect in transglutaminase 3 knockout mice. J. Invest. Dermatol. 2014, 134, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Ogunleye, T.A.; McMichael, A.; Olsen, E.A. Central centrifugal cicatricial alopecia: What has been achieved, current clues for future research. Dermatol. Clin. 2014, 32, 173–181. [Google Scholar] [CrossRef]
- Dlova, N.C.; Salkey, K.S.; Callender, V.D.; McMichael, A.J. Central Centrifugal Cicatricial Alopecia: New Insights and a Call for Action. J. Investig. Dermatol. Symp. Proc. 2017, 18, S54–S56. [Google Scholar] [CrossRef] [Green Version]
- Subash, J.; Alexander, T.; Beamer, V.; McMichael, A. A proposed mechanism for central centrifugal cicatricial alopecia. Exp. Dermatol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Jordan, C.S.; Chapman, C.; Kolivras, A.; Roberts, J.L.; Thompson, N.B.; Thompson, C.T. Clinicopathologic and immunophenotypic characterization of lichen planopilaris and central centrifugal cicatricial alopecia: A comparative study of 51 cases. J. Cutan. Pathol. 2019. [Google Scholar] [CrossRef]
- Mallet, A.; Kypriotou, M.; George, K.; Leclerc, E.; Rivero, D.; Mazereeuw-Hautier, J.; Serre, G.; Huber, M.; Jonca, N.; Hohl, D. Identification of the first nonsense CDSN mutation with expression of a truncated protein and causing Peeling skin syndrome type B. Br. J. Dermatol. 2013, 169, 1322–1325. [Google Scholar] [CrossRef]
- Caubet, C.; Bousset, L.; Clemmensen, O.; Sourigues, Y.; Bygum, A.; Chavanas, S.; Coudane, F.; Hsu, C.Y.; Betz, R.C.; Melki, R.; et al. A new amyloidosis caused by fibrillar aggregates of mutated corneodesmosin. FASEB J. 2010, 24, 3416–3426. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Identifier 1 | Protein Name (Abbreviation) | Deiminated Arg Number |
|---|---|---|
| P04264 | Keratin 1 (KRT1) | R65, R82 |
| P35908 | Keratin 2 (KRT2) | R603 |
| P04259 | Keratin 6B (KRT6B) | R59, R86 |
| O43290 | Keratin 14 (KRT14) | R510 |
| P08779 | Keratin 16 (KRT16) | R41 |
| Q04695 | Keratin 17 (KRT17) | R41, R191 |
| P20930 | Filaggrin (FLG) | R1026 or R3295, R1567 |
| Q5D862 | Filaggrin-2 (FLG2) | R637 or R789 |
| Q86YZ3 | Hornerin (HRNR) | R1953 |
| Q5T749 | Keratinocyte proline-rich protein (KPRP) | R409 |
| P53611 | Geranylgeranyltransferase (GGT) | R184 |
| Q7Z4L5 | Tetratricopeptide repeat protein 21B (TTC21B) | R297, R301 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méchin, M.-C.; Takahara, H.; Simon, M. Deimination and Peptidylarginine Deiminases in Skin Physiology and Diseases. Int. J. Mol. Sci. 2020, 21, 566. https://doi.org/10.3390/ijms21020566
Méchin M-C, Takahara H, Simon M. Deimination and Peptidylarginine Deiminases in Skin Physiology and Diseases. International Journal of Molecular Sciences. 2020; 21(2):566. https://doi.org/10.3390/ijms21020566
Chicago/Turabian StyleMéchin, Marie-Claire, Hidenari Takahara, and Michel Simon. 2020. "Deimination and Peptidylarginine Deiminases in Skin Physiology and Diseases" International Journal of Molecular Sciences 21, no. 2: 566. https://doi.org/10.3390/ijms21020566
APA StyleMéchin, M.-C., Takahara, H., & Simon, M. (2020). Deimination and Peptidylarginine Deiminases in Skin Physiology and Diseases. International Journal of Molecular Sciences, 21(2), 566. https://doi.org/10.3390/ijms21020566