1. Introduction
The fracture healing process consists of four overlapping phases, namely, inflammation, proliferation, callus formation, and bone remodeling. Immediately following fracture, the injury initiates an inflammatory response that is necessary to promote healing. The response induces the development of a hematoma, which consists of cells from both peripheral blood vessels and bone marrow. The hematoma coagulates between and around the fracture site and within the bone marrow, providing a template for callus formation [
1]. Vascularization supplies mesenchymal stem cells (MSCs), which differentiate into chondrocytes or osteoblasts simultaneously with cartilage tissue development (proliferation phase) [
2,
3]. The cartilage matrix begins to form at the fractured bone gap during the callus formation phase. Meanwhile, intramembranous ossification occurs internal to the periosteum adjacent to the fracture line and forms the bone matrix [
4]. MSCs directly differentiate into osteoblasts at the fracture site along the proximal and distal edges of fractured bone during intramembranous ossification. After cartilage tissue maturation, new bone formation is initiated as the cartilage tissue is resorbed and vascularization is induced to replace the cartilage tissue with bone. It has also been reported that primary bone formation is initiated peripheral to the newly formed cartilage region at the fractured bone site [
5]. The bone remodeling phase recapitulates embryonic bone development with a combination of cellular proliferation and differentiation, increasing the cellular volume and matrix deposition [
1]. Finally, remodeling of the hard callus into a lamellar bone structure occurs (bone remodeling phase).
The biological process occurring during bone fracture healing is regulated by several signaling molecules. Hedgehog (HH) proteins are among the signaling molecules required for endochondral bone formation during embryonic development, and they regulate bone homeostasis by controlling MSC proliferation [
6,
7]. HH signaling is also involved in the regulation of MSC proliferation in adult tissues. Aberrant activation of HH pathways has been linked to multiple types of human cancer [
7]. These pathways are also activated during intramembranous and endochondral ossification in the fracture healing process, but it is not clear if they are involved in the healing process [
5]. HH signaling pathways play critical roles in developmental processes and in the postnatal homeostasis of many tissues, including bone and cartilage. The HH family of intercellular signaling proteins plays important roles in regulating the development of many tissues and organs. Their name is derived from the observation of a hedgehog-like appearance in
Drosophila embryos with genetic mutations that block their action. Three types of HH proteins have been reported in mammals, namely Sonic HH (Shh), Indian HH (Ihh), and Desert HH (Dhh). Ihh is up-regulated during the initial stage of fracture repair, and it regulates differentiation indirectly by controlling cartilage development at the fracture site. Ihh regulates osteoblast differentiation indirectly by controlling cartilage development [
8]. In general, Shh acts in the early stages of development to regulate patterning and growth [
9]. Recently, several studies reported that Shh might be related to fracture healing [
10,
11]. Following the inactivation of HH signaling, the activity of Smo is inhibited by a receptor known as Patched (Ptch). Binding of the HH ligand Ptch relieves the inhibition of Smo, and activated Smo blocks the proteolysis of Gli proteins in the cytoplasm and promotes their dissociation from suppressor of fused (SuFu). Following dissociation from SuFu, activated Gli proteins translocate into the nucleus and promote the expression of Hh target genes, including
Gli1 [
9,
12]. Gli1 positivity has been identified as a marker for MSCs [
13]. Another study uncovered that Gli1 is involved in osteoblast differentiation [
14]. However, it is unclear that whether Shh proteins are involved in fracture healing. In this study, we demonstrated that Shh protein and the related proteins Smo and Gli1 were involved in osteoblast differentiation at the fracture healing site via immunohistochemical analysis.
2. Results and Discussion
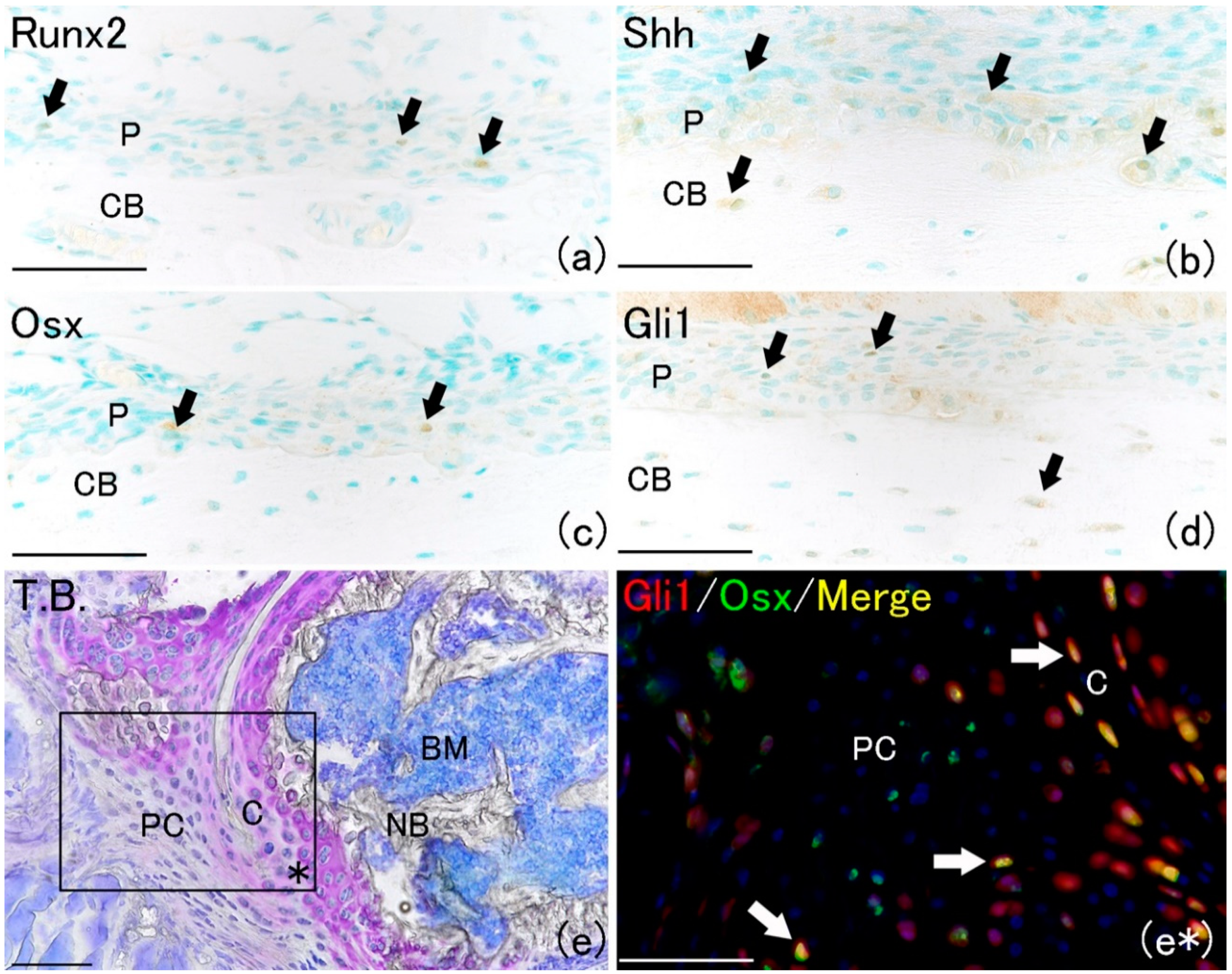
In this study, we hypothesized that Shh is related to the healing process of fractures and investigated and compared the positive localization of Runx2 and Osx, which appear during the fracture repair process, with that of Shh and its downstream factor Gli1. Runt-related transcription factor 2 (Runx2), which is an essential factor for bone formation, is expressed very early in skeletal development. Osterix (Osx) is activated downstream of Runx2 during osteoblastic lineage differentiation [
15,
16]. On the day of fracture (day 0), a few Runx2-positive and Osx-positive cells were observed on the bone surface in the periosteum (
Figure 1a,c). Shh-positive cells were rarely observed in the periosteum on day 0 (
Figure 1b). Furthermore, Gli1-positive cells were also rarely observed (
Figure 1d). However, Shh and Gli1 positivity were localized to osteocytes in the bone matrix. These results indicated that Shh signaling occurred in osteocytes but not in undifferentiated cells in the periosteum. Moreover, in this study, we traced the fate of Gli1-positive cells in Gli1-Cre recombinase-mutated estrogen receptors (CreER
T2); tdTomato mice on day seven after fracture by administering tamoxifen. Previous reports demonstrated that 3 days are required for Cre activation after tamoxifen administration [
8]. In our genetically modified mouse system, both Gli1-positive cells and their progeny were permanently marked by red fluorescent protein expression. Gli1-CreER
T2; tdTomato mice, which are transgenic for the
Gli1-CreERT2/Rosa26-loxP-stop-loxP-tdTomato gene, were used to generate Gli1-positive and progeny cells through lineage-tracing analysis. Gli1-positive cells expressed the CreER
T2. CreER
T2 has the function of specifically recognizing and removing the LoxP site. Moreover, CreER
T2 binds to tamoxifen but not to natural estrogens. Gli1-positive cells were found to express tomato red fluorescence after tamoxifen administration. Since tomato fluorescence is expressed permanently, not only Gli1-positive cells but also progeny cells were found to express tomato red fluorescence [
17].
New cartilage matrix formed around the fracture site (
Figure 1e). The localization of Gli1 progeny cells and Osx-positive cells was examined at the fracture healing site (
Figure 1e*). Gli1 positivity was observed in chondrocytes and the perichondrium around the cartilage matrix at the fracture site (
Figure 1e*). Osx positivity was localized in cells surrounding new cartilage matrix. The merged image of Gli1-positive and Osx-positive areas indicated that most Osx-positive cells were co-localized with Gli1-positive cells (
Figure 1e*). These results indicate that Gli-positive cells and their progeny cells might differentiate into osteoblasts after bone fracture. This result was consistent with another report that Gli1 marked a major skeletal progenitor pool contributing to both bone and cartilage formation during bone fracture healing in postnatal mice [
8].
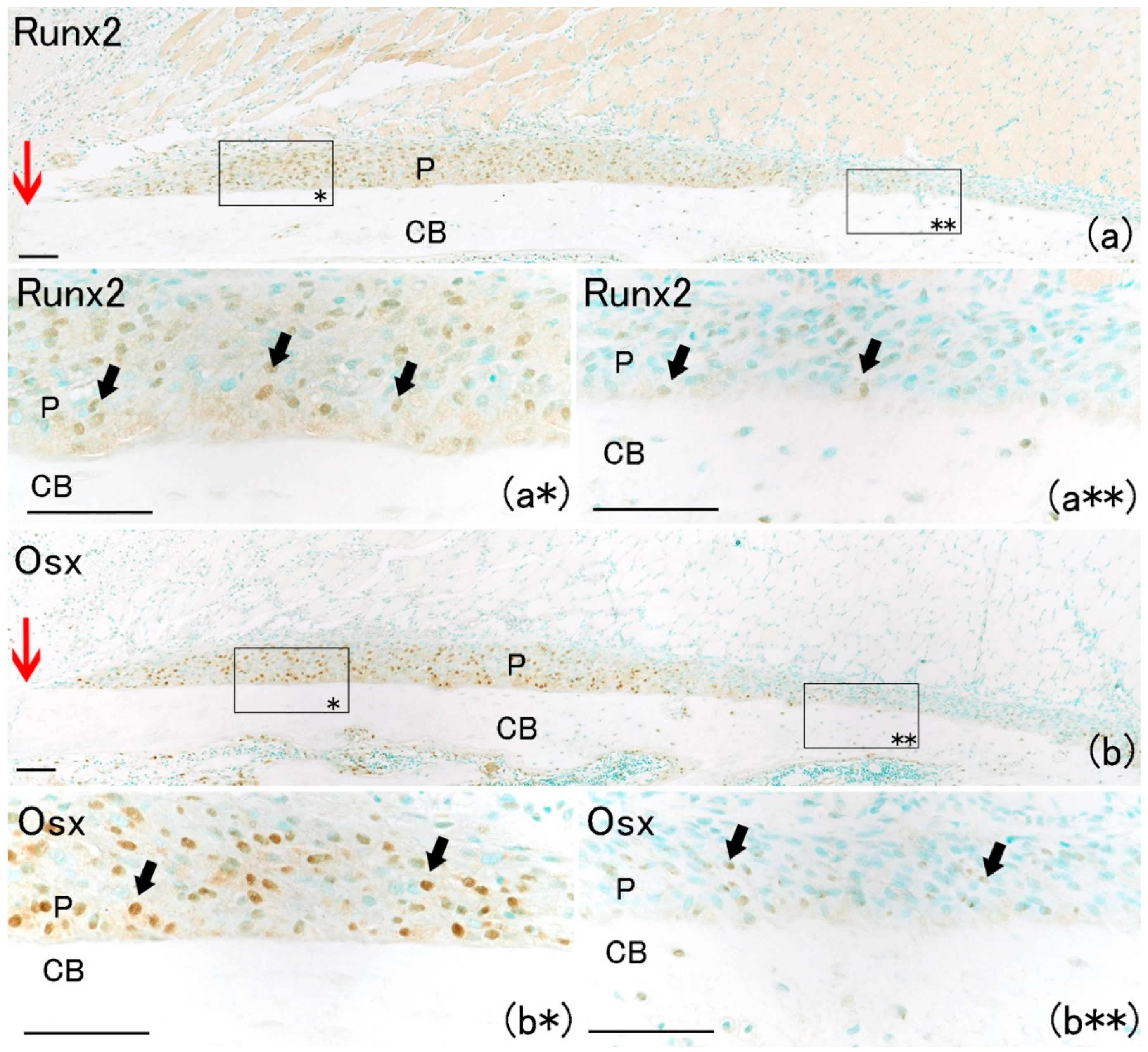
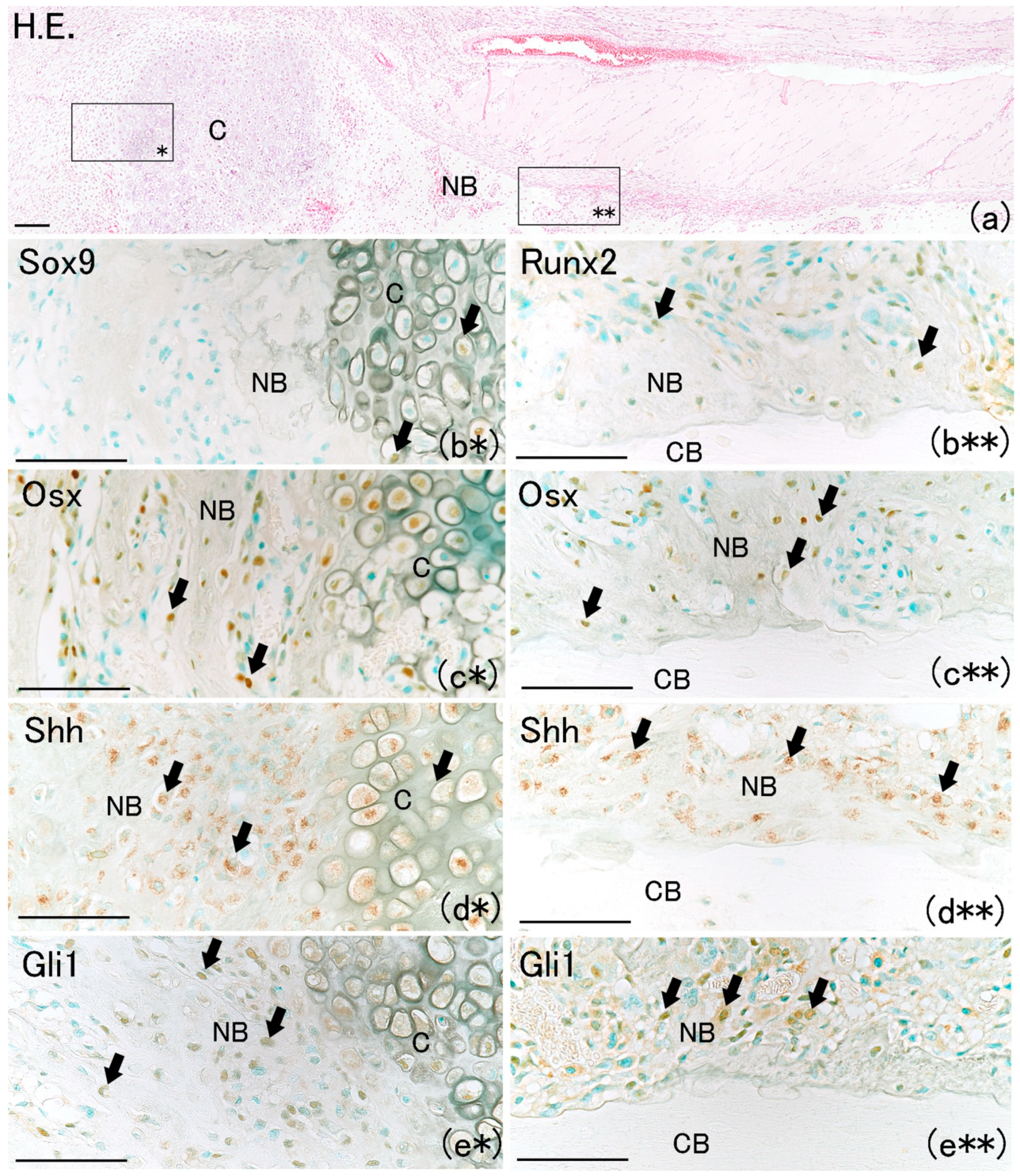
On day 1, hematoma and granulation tissue were observed at the bone fracture gap. Runx2-positive and Osx-positive cells were observed extensively in the remaining periosteum near the fracture site (
Figure 2a*,b*). However, few Runx2-positive and Osx-positive cells were noted in the intact periosteum far from the fracture site (
Figure 2a**,b**). Runx2-positive and Osx-positive cell numbers in the periosteum near the fracture site were remarkably higher than those far from the fracture site (
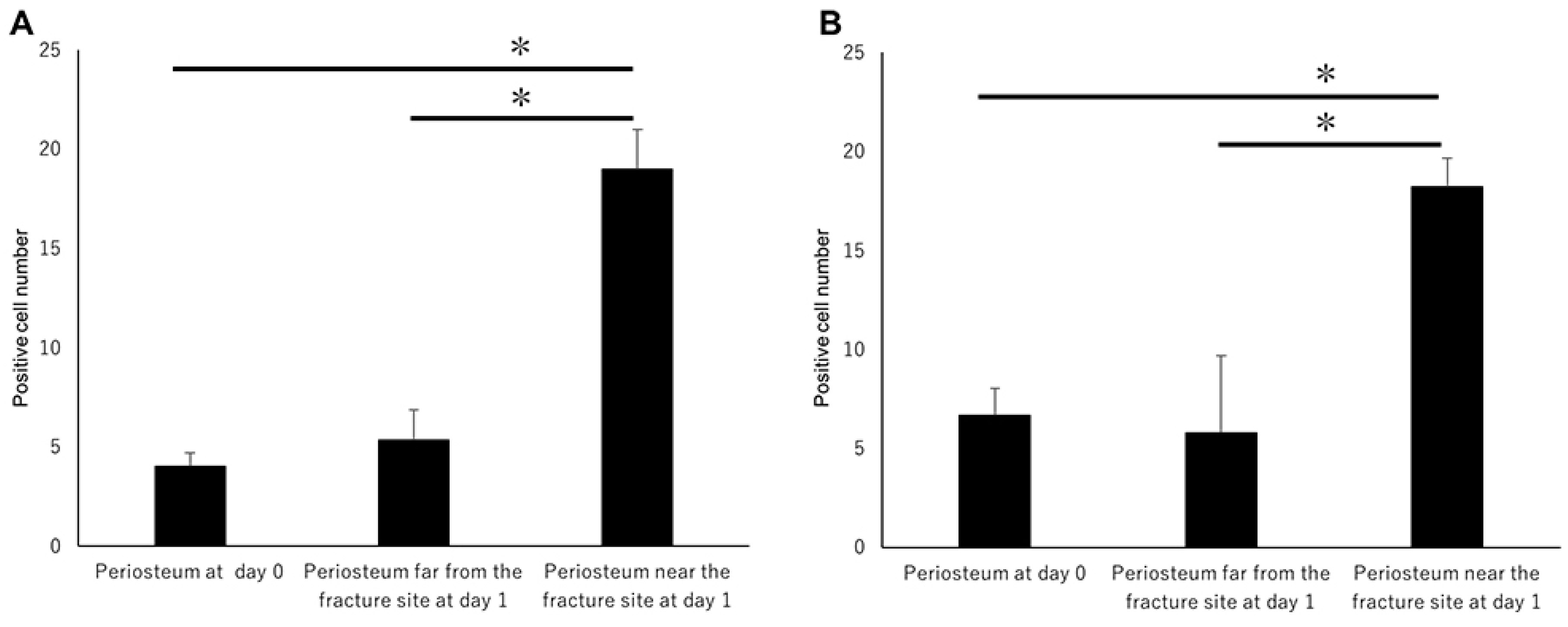
Figure 3A,B). These results indicated that MSCs committed to osteoblast or chondroblast differentiation participated in intramembranous and endochondral ossification only near the fracture site.
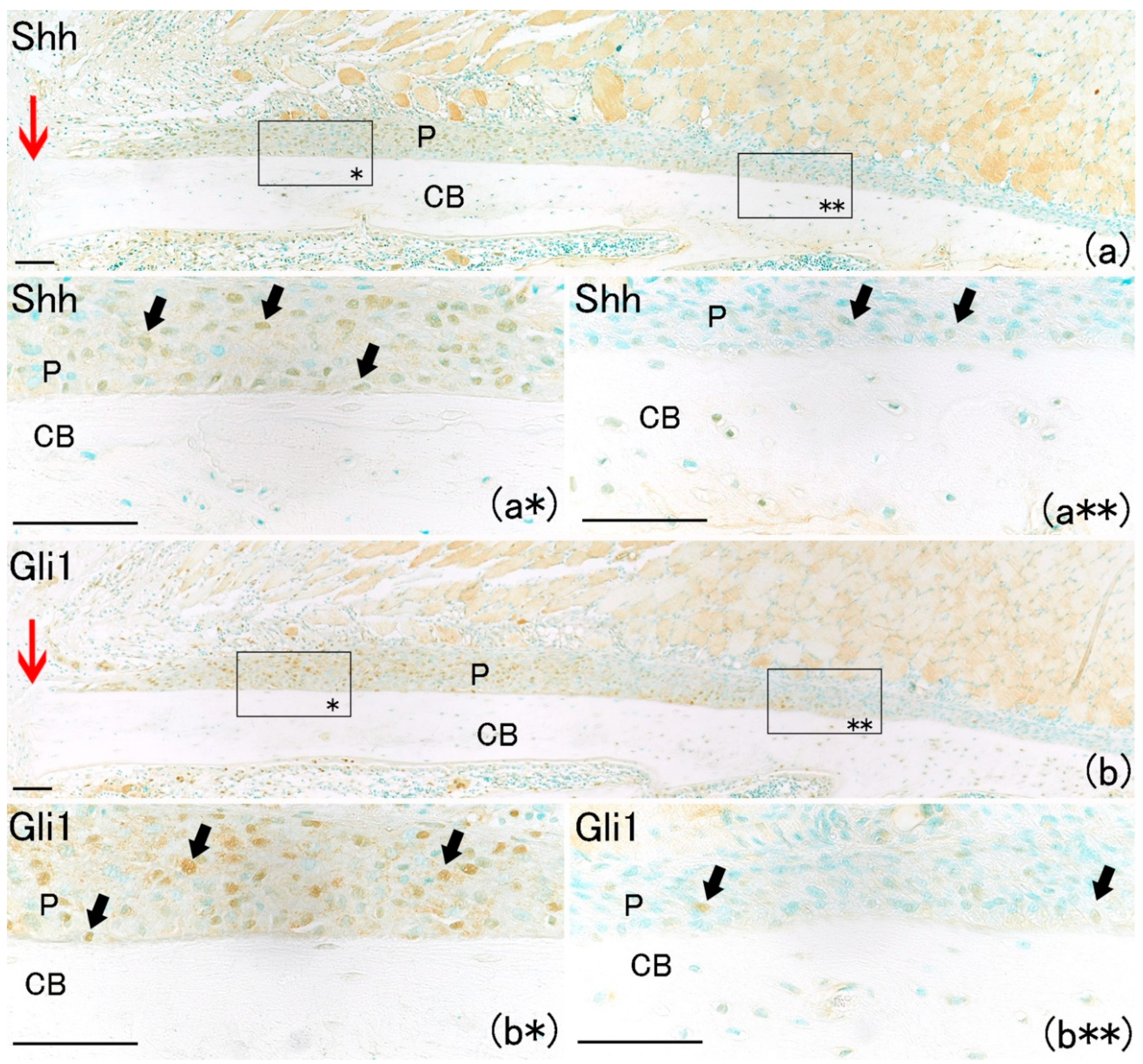
Large numbers of Shh-positive and Gli1-positive cells were also found in the periosteum only near the fracture site (
Figure 4a*,b*). Their numbers increased on day one in the periosteum adjacent to the fracture site compared with the number of Shh-positive and Gli1-positive cells on day 0 (
Figure 5A,B). Shh-positive and Gli1-positive cell numbers in the periosteum near the fracture site were remarkably higher than those far from the fracture site (
Figure 5A,B). These results indicate that Shh and Gli1, which emerge after bone fracture in the periosteum, might be associated with osteoblast differentiation.
On day 7, a newly formed cartilage matrix was observed in the fracture gap (
Figure 6a). It has been reported that endochondral ossification is observed in the fracture gap at that time [
18,
19]. Sox9 positivity was observed in chondrocytes as well as on the bone matrix surface (
Figure 6b*). Runx2-positive and Osx-positive cells were observed on the surfaces of newly formed bone matrix and chondrocytes (
Figure 6b**,c*,c**). Shh-positive and Gli1-positive cells were also observed on the surfaces of newly formed bone matrix and chondrocytes (
Figure 6d*,d**,e*,e**). In addition, new bone matrix extending from the proximal and distal edges of the fractured bone surface, which is termed intramembranous ossification, was observed (
Figure 6a). A large number of Osx-positive cells were also observed on the surface of newly formed bone extending from the proximal and distal edges. Osx-positive cells were localized on the new bone surface around the newly formed cartilage matrix (
Figure 6c*). This result is consistent with another report in which MSCs directly differentiated into osteoblasts in the perichondrium around the cartilage matrix after bone fracture, resulting in bone formation [
20]. Shh-positive and Gli1-positive cells localized along the surface of newly formed bone (
Figure 6d*,d**,e*,e**). These results indicate that the Shh–Gli1 signaling pathway might regulate intramembranous and endochondral ossification at the fracture site.
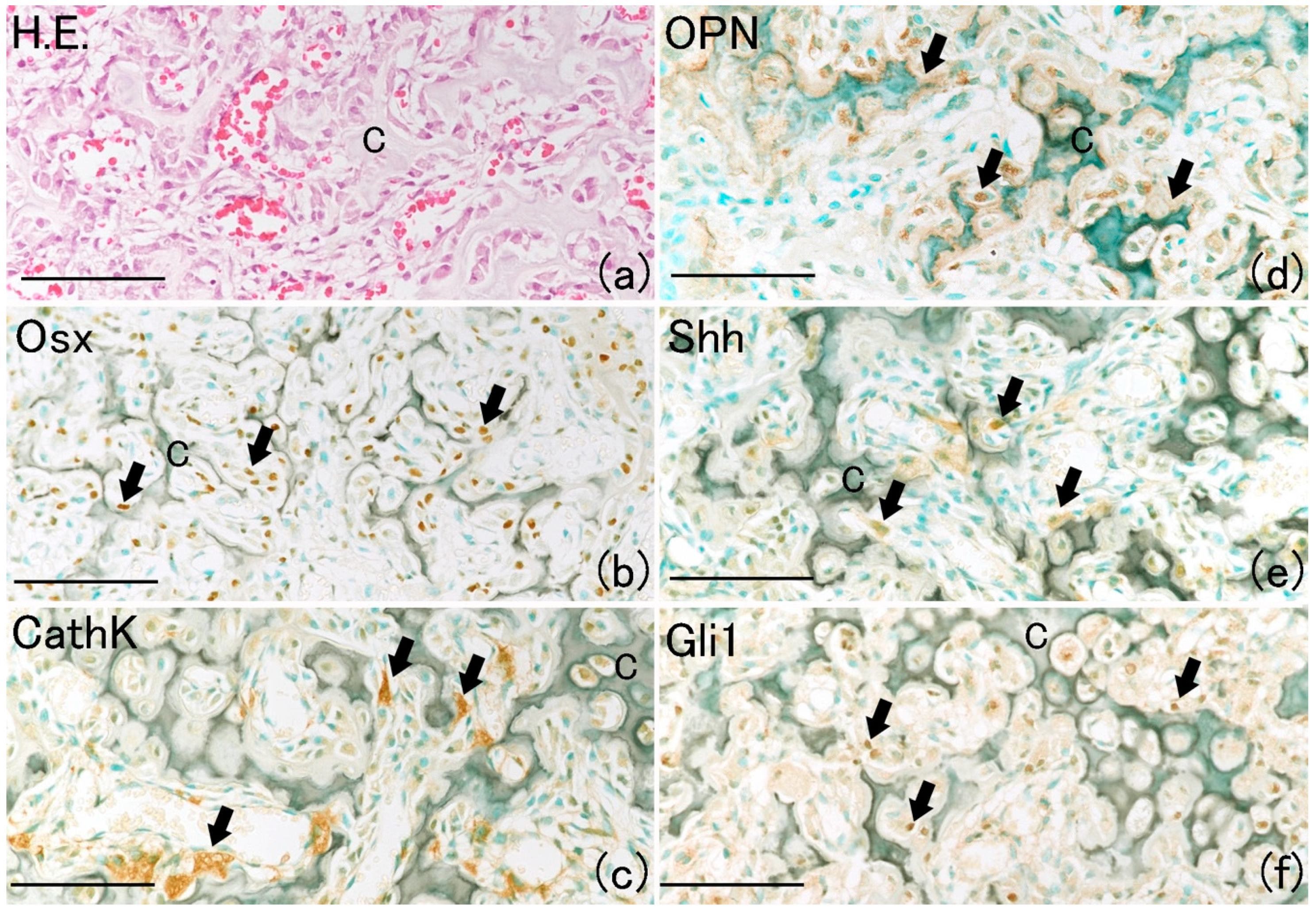
On day 14, newly formed cartilage matrix at the fracture site began to resorb, and it was replaced by newly formed bone known as primary bone (
Figure 7a) [
21]. In the resorbed cartilage area, many Osx-positive and cathepsin K (CathK)-positive cells were observed (
Figure 7b,c). Positivity for osteopontin (OPN), which is a bone matrix component, was observed around the resorbed cartilage matrix (
Figure 7d) [
22]. Shh and Gli1 positivity localized around the resorbed cartilage matrix (
Figure 7e,f). These results indicate that the Shh–Gli1 signaling pathway participates in new bone formation by osteoblasts.
3. Materials and Methods
3.1. Experimental Animals
Twenty-six-week-old male Sprague–Dawley rats (Hokudo, Sapporo, Japan) and three Gli-CreERT2; tdTomato male mice (Jackson Laboratory, Bar Harbor, ME, USA) were used. All experimental animals were maintained in a specific pathogen-free facility. All experiments were approved and performed according to guidelines set forth by the Animal Ethics Committee of the Health Sciences University of Hokkaido (The ethical permission code and permission date: 19-028, 8 March, 2019 and 19-045, 29 March 2019).
3.2. Tamoxifen Administration
Gli1-CreERT2; tdTomato mice were injected intraperitoneally with tamoxifen (Sigma-Aldrich, St. Louis, MO, USA) once daily for 3 consecutive days (40 mg/mL, dissolved in corn oil).
3.3. Fracture Experiment
The right eighth rib of each experimental animal was fractured as previously described [
10]. Briefly, each experimental animal was anesthetized, and the eighth rib on the right side was exposed and cut vertical to the axis with scissors. As a control, the right eighth rib of select animals was similarly exposed but not fractured.
3.4. Tissue Preparation
The animals were anesthetized subcutaneously with pentobarbital sodium (40 mg/kg) and killed via cervical dislocation. The ribs of Gli1-CreERT2; tdTomato mice were collected 7 days after fracture and immediately frozen at −80 °C. Each sample was embedded in 5 % carboxymethyl cellulose (CMC) gel (Section-Lab Co. Ltd., Tokyo, Japan). Each frozen CMC sample was covered with polyvinylidene chloride film (Section-Lab Co. Ltd.) and sagittally sectioned at a thickness of 5 µm. The ribs of rats were collected 0, 1, 7, and 14 days after fracture and fixed in 4.0% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) overnight at 4 °C. Specimens were demineralized via immersion in 10% ethylenediaminetetraacetic acid (pH 7.4) for 4 weeks at 4 °C. Following demineralization, the specimens were embedded in paraffin and sectioned at a thickness of 5 µm.
3.5. Immunohistochemistry
For immunohistochemistry, the dehydrated sections were treated with 0.3% H2O2 in phosphate-buffered saline (PBS; pH 7.4) for 30 min at room temperature to inactivate endogenous peroxidase. Sections were pretreated with 3% bovine serum albumin in PBS for 30 min at room temperature, followed by incubation with primary antibodies against Shh (1:100, Bioss, Woburn, MA, USA), Gli1 (1:100, Novus Biologicals, Centennial, CO, USA), Osx (1:1000, Abcam, Cambridge, MA, USA), Runx2 (1:1000, MBL, Nagoya, Japan), and OPN (1:100, antibody was kindly provided by Dr. Hiroaki Nakamura, Matsumoto Dental University, Japan) overnight at 4 °C. Sections were reacted with Histofine Simple Stain rat MAX-PO (MULTI; Nichirei, Tokyo, Japan) for 1 h at room temperature. Color was developed using liquid diaminobenzidine substrate-chromogen system (Dako, Carpinteria, CA, USA). Immunostained sections were then counterstained with methylene green.
3.6. Image Analysis
The 500 μm portion from the fracture line of the cortical bone to the midshaft was defined as the periosteum near the fracture site, and the 2000 μm portion from the fracture line of the cortical bone to the midshaft was defined as the periosteum far from the fracture site. The number of positive cells of Runx2, Osx, Shh, and Gli1 present in the periosteum near and far from the fracture were counted by defining a square (100 × 100 μm2).
3.7. Statistical Analysis
The statistical analysis of the data gathered from Runx2-, Osx-, Shh-, and Gli1-positive cell counting was performed using SPSS version 23 (SPSS Inc., Chicago, IL, USA). Analyses of variance were followed by the t-test and Tukey’s test to determine significance.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






