The Effects of Dietary Interventions on DNA Methylation: Implications for Obesity Management


Abstract
:1. Introduction
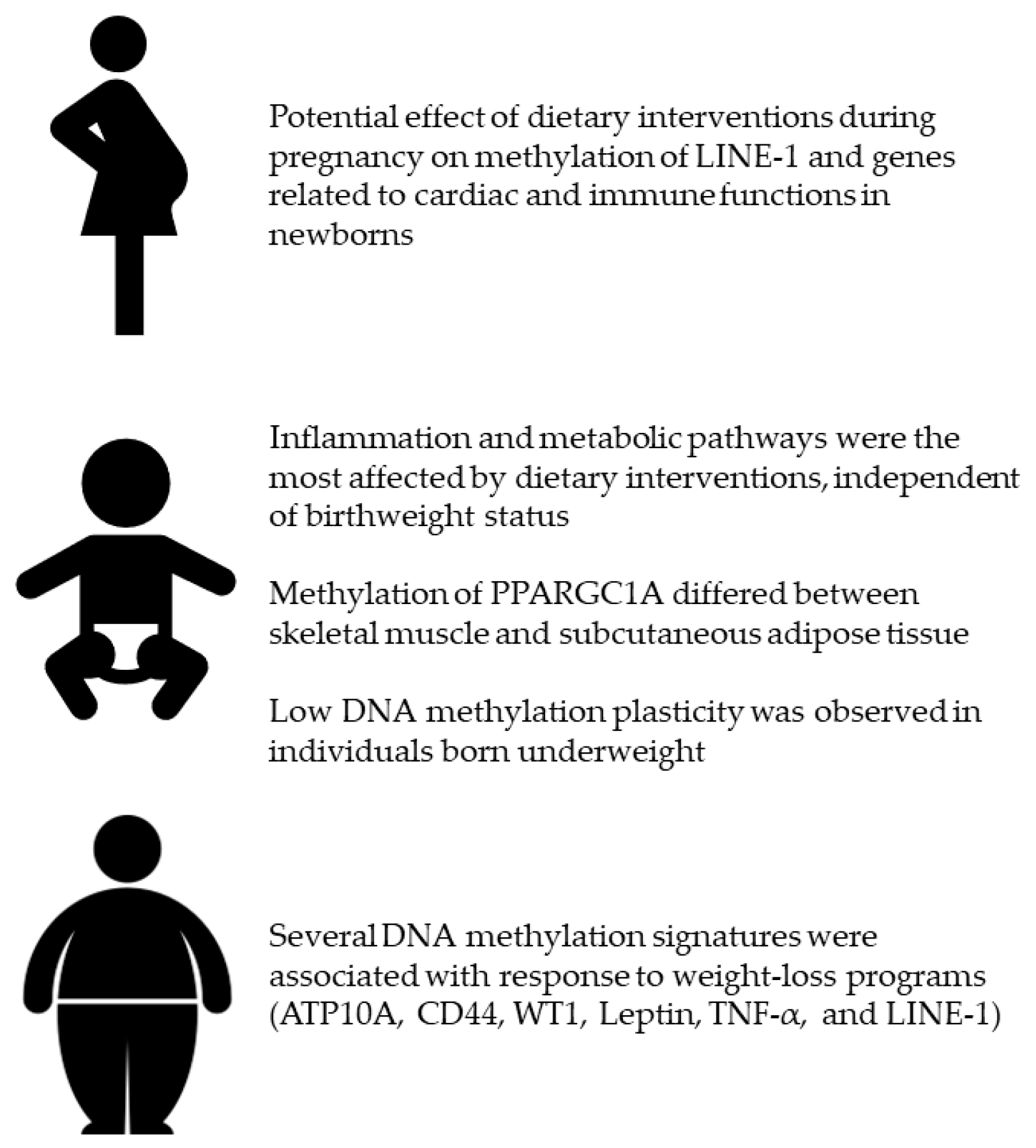
2. Dietary Interventions during Pregnancy
3. The Effect of Interventions in Adults According to Their Birthweight
4. Dietary Interventions against Obesity and Related Disorders
5. Discussion
Funding
Conflicts of Interest
Abbreviations
| WHO | World Health Organization |
| DNMTs | DNA methyltransferases |
| IGF2 | Insulin-like growth factor 2 |
| LINE-1 | Long interspersed nuclear elements 1 |
| BMI | Body Mass Index |
| PPARGC1A | Proliferator-activated receptor-γ, coactivator-1α |
| FADS2 | Fatty Acid Desaturase 2 |
| CPLX1 | Complexin 1 |
| IGFBP5 | Insulin-like growth factor-binding protein 5 |
| SLC2A4 | Solute Carrier Family 2 Member 4 |
| LEP | Leptin |
| ADIPQ | Adiponectin |
| MTHFR | Methylenetetrahydrofolate reductase |
| PREDIMED | Prevención con Dieta Mediterránea |
| GOLDN | Genetics of Lipid Lowering Drugs and Diet Network |
| LPP | Lipoma-preferred partner |
| APOA5 | Apolipoprotein A-5 |
| SREBF1 | Sterol regulatory element-binding transcription factor 1 |
| ABCG1 | ATP-binding cassette sub-family G member 1 |
| CPT1A | Carnitine palmitoyl-transferase 1-A |
| PUFA | Polyunsaturated fatty acids |
| SFA | Saturated fatty acids |
| FTO | Alpha-ketoglutarate dependent dioxygenase |
| IL6 | Interleukin 6 |
| INSR | Insulin receptor |
| NEGR1 | Neuronal growth regulator 1 |
| POMC | Proopiomelanocortin |
| ATP10A | ATPase Phospholipid Transporting 10A |
| WT1 | Wilms’ tumor 1 |
| TNF-α | Tumor Necrosis Factor Alpha |
| SERPINE-1 | Serpin Family E Member 1 |
| RESMENA | Metabolic Syndrome Reduction in Navarra |
| AHA | American Heart Association |
| BMAL1 | Brain and muscle aryl hydrocarbon receptor nuclear translocator–like protein 1 |
| NFATC2IP | Nuclear Factor Of Activated T Cells 2 Interacting Protein |
References
- González-Muniesa, P.; Mártinez-González, M.A.; Hu, F.B.; Després, J.P.; Matsuzawa, Y.; Loos, R.J.F.; Moreno, L.A.; Bray, G.A.; Martinez, J.A. Obesity. Nat. Rev. Dis. Primers 2017, 3, 17034. [Google Scholar] [CrossRef]
- Williams, E.P.; Mesidor, M.; Winters, K.; Dubbert, P.M.; Wyatt, S.B. Overweight and obesity: Prevalence, consequences, and causes of a growing public health problem. Curr. Obes. Rep. 2015, 4, 363–370. [Google Scholar] [CrossRef]
- Kroes, M.; Osei-Assibey, G.; Baker-Searle, R.; Huang, J. Impact of weight change on quality of life in adults with overweight/obesity in the United States: A systematic review. Curr. Med. Res. Opin. 2016, 32, 485–508. [Google Scholar] [CrossRef]
- Genné-Bacon, E.A. Thinking evolutionarily about obesity. Yale J. Biol. Med. 2014, 87, 99–112. [Google Scholar] [PubMed]
- Sellayah, D.; Cagampang, F.R.; Cox, R.D. On the evolutionary origins of obesity: A new hypothesis. Endocrinology 2014, 155, 1573–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maugeri, A.; Kunzova, S.; Medina-Inojosa, J.R.; Agodi, A.; Barchitta, M.; Homolka, M.; Kiacova, N.; Bauerova, H.; Sochor, O.; Lopez-Jimenez, F.; et al. Association between eating time interval and frequency with ideal cardiovascular health: Results from a random sample Czech urban population. Nutr. Metab. Cardiovasc. Dis. 2018. [Google Scholar] [CrossRef]
- Agodi, A.; Maugeri, A.; Kunzova, S.; Sochor, O.; Bauerova, H.; Kiacova, N.; Barchitta, M.; Vinciguerra, M. Association of dietary patterns with metabolic syndrome: Results from the Kardiovize Brno 2030 Study. Nutrients 2018, 10, 898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maugeri, A.; Barchitta, M.; Fiore, V.; Rosta, G.; Favara, G.; La Mastra, C.; La Rosa, M.C.; Magnano San Lio, R.; Agodi, A. Determinants of adherence to the Mediterranean diet: Findings from a cross-sectional study in women from Southern Italy. Int. J. Environ. Res. Public Health 2019, 16, 2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunzova, S.; Maugeri, A.; Medina-Inojosa, J.; Lopez-Jimenez, F.; Vinciguerra, M.; Marques-Vidal, P. Determinants of metabolic health across body mass index categories in Central Europe: A comparison between Swiss and Czech populations. Front. Public Health 2020, 8, 108. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, A.; Barchitta, M.; Agrifoglio, O.; Favara, G.; La Mastra, C.; La Rosa, M.C.; Magnano San Lio, R.; Panella, M.; Cianci, A.; Agodi, A. The impact of social determinants and lifestyles on dietary patterns during pregnancy: Evidence from the “Mamma & Bambino” study. Ann. Ig. 2019, 31. [Google Scholar] [CrossRef]
- Maugeri, A.; Barchitta, M.; Favara, G.; La Rosa, M.C.; La Mastra, C.; Magnano San Lio, R.; Agodi, A. Maternal dietary patterns are associated with pre-pregnancy body mass index and gestational weight gain: Results from the “Mamma & Bambin” cohort. Nutrients 2019, 11, 1308. [Google Scholar] [CrossRef] [Green Version]
- Medina-Inojosa, J.R.; Vinciguerra, M.; Maugeri, A.; Kunzova, S.; Sochor, O.; Movsisyan, N.; Geda, Y.E.; Stokin, G.B.; Lopez-Jimenez, F. Prevalence of ideal cardiovascular health in a Central European community: Results from the Kardiovize Brno 2030 Project. Eur. J. Prev. Cardiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, A.; Medina-Inojosa, J.R.; Kunzova, S.; Agodi, A.; Barchitta, M.; Sochor, O.; Lopez-Jimenez, F.; Geda, Y.E.; Vinciguerra, M. Sleep duration and excessive daytime sleepiness are associated with obesity independent of diet and physical activity. Nutrients 2018, 10, 1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/en/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 10 September 2020).
- Bray, G.A.; Siri-Tarino, P.W. The role of macronutrient content in the diet for weight management. Endocrinol. Metab. Clin. North. Am. 2016, 45, 581–604. [Google Scholar] [CrossRef]
- Martinez, J.A.; Navas-Carretero, S.; Saris, W.H.; Astrup, A. Personalized weight loss strategies—The role of macronutrient distribution. Nat. Rev. Endocrinol. 2014, 10, 749–760. [Google Scholar] [CrossRef] [Green Version]
- Mozaffarian, D. Dietary and policy priorities for cardiovascular disease, diabetes, and obesity: A comprehensive review. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef]
- Winkler, T.W.; Justice, A.E.; Graff, M.; Barata, L.; Feitosa, M.F.; Chu, S.; Czajkowski, J.; Esko, T.; Fall, T.; Kilpeläinen, T.O.; et al. The influence of age and sex on genetic associations with adult body size and shape: A large-scale genome-wide interaction study. PLoS Genet. 2015, 11, e1005378. [Google Scholar] [CrossRef] [Green Version]
- Lo Re, O.; Maugeri, A.; Hruskova, J.; Jakubik, J.; Kucera, J.; Bienertova-Vasku, J.; Oben, J.A.; Kubala, L.; Dvorakova, A.; Ciz, M.; et al. Obesity-induced nucleosome release predicts poor cardio-metabolic health. Clin. Epigenetics 2019, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Elks, C.E.; den Hoed, M.; Zhao, J.H.; Sharp, S.J.; Wareham, N.J.; Loos, R.J.; Ong, K.K. Variability in the heritability of body mass index: A systematic review and meta-regression. Front. Endocrinol. 2012, 3, 29. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.H.; Fei, J.; Lan, M.S.; Lu, Z.P.; Liu, M.; Fan, W.W.; Gao, X.; Lu, D.R. Hypermethylation of hepatic Gck promoter in ageing rats contributes to diabetogenic potential. Diabetologia 2008, 51, 1525–1533. [Google Scholar] [CrossRef] [Green Version]
- Milagro, F.I.; Campión, J.; Cordero, P.; Goyenechea, E.; Gómez-Uriz, A.M.; Abete, I.; Zulet, M.A.; Martínez, J.A. A dual epigenomic approach for the search of obesity biomarkers: DNA methylation in relation to diet-induced weight loss. FASEB J. 2011, 25, 1378–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, S.; Olek, A. Diabetes: A candidate disease for efficient DNA methylation profiling. J. Nutr. 2002, 132, 2440S–2443S. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Erickson, S.W.; MacLeod, S.L.; Cleves, M.A.; Hu, P.; Karim, M.A.; Hobbs, C.A. Maternal genome-wide DNA methylation patterns and congenital heart defects. PLoS ONE 2011, 6, e16506. [Google Scholar] [CrossRef] [Green Version]
- Maugeri, A.; Barchitta, M. How dietary factors affect DNA methylation: Lesson from epidemiological studies. Medicina 2020, 56, 374. [Google Scholar] [CrossRef] [PubMed]
- ElGendy, K.; Malcomson, F.C.; Lara, J.G.; Bradburn, D.M.; Mathers, J.C. Effects of dietary interventions on DNA methylation in adult humans: Systematic review and meta-analysis. Br. J. Nutr. 2018, 120, 961–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanley, B.; Dijane, J.; Fewtrell, M.; Grynberg, A.; Hummel, S.; Junien, C.; Koletzko, B.; Lewis, S.; Renz, H.; Symonds, M.; et al. Metabolic imprinting, programming and epigenetics-a review of present priorities and future opportunities. Br. J. Nutr. 2010, 104, S1–S25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, J.G. Developmental origins of health and disease-from a small body size at birth to epigenetics. Ann. Med. 2016, 48, 456–467. [Google Scholar] [CrossRef]
- Tielemans, M.J.; Garcia, A.H.; Peralta Santos, A.; Bramer, W.M.; Luksa, N.; Luvizotto, M.J.; Moreira, E.; Topi, G.; de Jonge, E.A.; Visser, T.L.; et al. Macronutrient composition and gestational weight gain: A systematic review. Am. J. Clin. Nutr. 2016, 103, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Dearden, L.; Ozanne, S.E. Early life origins of metabolic disease: Developmental programming of hypothalamic pathways controlling energy homeostasis. Front. Neuroendocrinol. 2015, 39, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Lim, I.Y.; Wu, Y.; Teh, A.L.; Chen, L.; Aris, I.M.; Soh, S.E.; Tint, M.T.; MacIsaac, J.L.; Morin, A.M.; et al. Developmental pathways to adiposity begin before birth and are influenced by genotype, prenatal environment and epigenome. BMC Med. 2017, 15, 50. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.B.; Han, S.P.; Zhu, G.Z.; Zhu, C.; Wang, X.J.; Cao, X.G.; Guo, X.R. Birth weight and subsequent risk of obesity: A systematic review and meta-analysis. Obes. Rev. 2011, 12, 525–542. [Google Scholar] [CrossRef] [PubMed]
- Labayen, I.; Ruiz, J.R.; Vicente-Rodríguez, G.; Turck, D.; Rodríguez, G.; Meirhaeghe, A.; Molnár, D.; Sjöström, M.; Castillo, M.J.; Gottrand, F.; et al. Early life programming of abdominal adiposity in adolescents: The HELENA Study. Diabetes Care 2009, 32, 2120–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravelli, G.P.; Stein, Z.A.; Susser, M.W. Obesity in young men after famine exposure in utero and early infancy. N. Engl. J. Med. 1976, 295, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Tobi, E.W.; Lumey, L.H.; Talens, R.P.; Kremer, D.; Putter, H.; Stein, A.D.; Slagboom, P.E.; Heijmans, B.T. DNA methylation differences after exposure to prenatal famine are common and timing- and sex-specific. Hum. Mol. Genet. 2009, 18, 4046–4053. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Barraza-Villarreal, A.; Hernandez-Vargas, H.; Sly, P.D.; Biessy, C.; Ramakrishnan, U.; Romieu, I.; Herceg, Z. Modulation of DNA methylation states and infant immune system by dietary supplementation with ω-3 PUFA during pregnancy in an intervention study. Am. J. Clin. Nutr. 2013, 98, 480–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraghty, A.A.; Sexton-Oates, A.; O’Brien, E.C.; Alberdi, G.; Fransquet, P.; Saffery, R.; McAuliffe, F.M. A Low glycaemic index diet in pregnancy induces DNA methylation variation in blood of newborns: Results from the ROLO randomised controlled trial. Nutrients 2018, 10, 455. [Google Scholar] [CrossRef] [Green Version]
- Brøns, C.; Jacobsen, S.; Nilsson, E.; Rönn, T.; Jensen, C.B.; Storgaard, H.; Poulsen, P.; Groop, L.; Ling, C.; Astrup, A.; et al. Deoxyribonucleic acid methylation and gene expression of PPARGC1A in human muscle is influenced by high-fat overfeeding in a birth-weight-dependent manner. J. Clin. Endocrinol. Metab. 2010, 95, 3048–3056. [Google Scholar] [CrossRef]
- Gillberg, L.; Jacobsen, S.C.; Rönn, T.; Brøns, C.; Vaag, A. PPARGC1A DNA methylation in subcutaneous adipose tissue in low birth weight subjects—impact of 5 days of high-fat overfeeding. Metabolism 2014, 63, 263–271. [Google Scholar] [CrossRef]
- Jacobsen, S.C.; Brøns, C.; Bork-Jensen, J.; Ribel-Madsen, R.; Yang, B.; Lara, E.; Hall, E.; Calvanese, V.; Nilsson, E.; Jørgensen, S.W.; et al. Effects of short-term high-fat overfeeding on genome-wide DNA methylation in the skeletal muscle of healthy young men. Diabetologia 2012, 55, 3341–3349. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, S.C.; Gillberg, L.; Bork-Jensen, J.; Ribel-Madsen, R.; Lara, E.; Calvanese, V.; Ling, C.; Fernandez, A.F.; Fraga, M.F.; Poulsen, P.; et al. Young men with low birthweight exhibit decreased plasticity of genome-wide muscle DNA methylation by high-fat overfeeding. Diabetologia 2014, 57, 1154–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillberg, L.; Perfilyev, A.; Brøns, C.; Thomasen, M.; Grunnet, L.G.; Volkov, P.; Rosqvist, F.; Iggman, D.; Dahlman, I.; Risérus, U.; et al. Adipose tissue transcriptomics and epigenomics in low birthweight men and controls: Role of high-fat overfeeding. Diabetologia 2016, 59, 799–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hjort, L.; Jørgensen, S.W.; Gillberg, L.; Hall, E.; Brøns, C.; Frystyk, J.; Vaag, A.A.; Ling, C. 36 h fasting of young men influences adipose tissue DNA methylation of. Clin. Epigenetics 2017, 9, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samblas, M.; Mansego, M.L.; Zulet, M.A.; Milagro, F.I.; Martinez, J.A. An integrated transcriptomic and epigenomic analysis identifies CD44 gene as a potential biomarker for weight loss within an energy-restricted program. Eur. J. Nutr. 2019, 58, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Cordero, P.; Campion, J.; Milagro, F.I.; Goyenechea, E.; Steemburgo, T.; Javierre, B.M.; Martinez, J.A. Leptin and TNF-alpha promoter methylation levels measured by MSP could predict the response to a low-calorie diet. J. Physiol. Biochem. 2011, 67, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Campión, J.; Milagro, F.I.; Goyenechea, E.; Martínez, J.A. TNF-alpha promoter methylation as a predictive biomarker for weight-loss response. Obesity 2009, 17, 1293–1297. [Google Scholar] [CrossRef]
- Nicoletti, C.F.; Nonino, C.B.; de Oliveira, B.A.; Pinhel, M.A.; Mansego, M.L.; Milagro, F.I.; Zulet, M.A.; Martinez, J.A. DNA methylation and hydroxymethylation levels in relation to two weight loss strategies: Energy-restricted diet or bariatric surgery. Obes. Surg. 2016, 26, 603–611. [Google Scholar] [CrossRef]
- Delgado-Cruzata, L.; Zhang, W.; McDonald, J.A.; Tsai, W.Y.; Valdovinos, C.; Falci, L.; Wang, Q.; Crew, K.D.; Santella, R.M.; Hershman, D.L.; et al. Dietary modifications, weight loss, and changes in metabolic markers affect global DNA methylation in Hispanic, African American, and Afro-Caribbean breast cancer survivors. J. Nutr. 2015, 145, 783–790. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Lacarte, M.; Milagro, F.I.; Zulet, M.A.; Martinez, J.A.; Mansego, M.L. LINE-1 methylation levels, a biomarker of weight loss in obese subjects, are influenced by dietary antioxidant capacity. Redox Rep. 2016, 21, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Martín-Núñez, G.M.; Cabrera-Mulero, R.; Rubio-Martín, E.; Rojo-Martínez, G.; Olveira, G.; Valdés, S.; Soriguer, F.; Castaño, L.; Morcillo, S. Methylation levels of the SCD1 gene promoter and LINE-1 repeat region are associated with weight change: An intervention study. Mol. Nutr. Food Res. 2014, 58, 1528–1536. [Google Scholar] [CrossRef]
- Duggan, C.; Xiao, L.; Terry, M.B.; McTiernan, A. No effect of weight loss on LINE-1 methylation levels in peripheral blood leukocytes from postmenopausal overweight women. Obesity 2014, 22, 2091–2096. [Google Scholar] [CrossRef] [Green Version]
- Samblas, M.; Milagro, F.I.; Gómez-Abellán, P.; Martínez, J.A.; Garaulet, M. Methylation on the circadian gene BMAL1 is associated with the effects of a weight loss intervention on serum lipid levels. J. Biol. Rhythm. 2016, 31, 308–317. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Heianza, Y.; Li, X.; Shang, X.; Smith, S.R.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetic, epigenetic and transcriptional variations at NFATC2IP locus with weight loss in response to diet interventions: The POUNDS Lost Trial. Diabetes Obes. Metab. 2018, 20, 2298–2303. [Google Scholar] [CrossRef]
- Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Vinciguerra, M.; Agodi, A. LINE-1 hypomethylation in blood and tissue samples as an epigenetic marker for cancer risk: A systematic review and meta-analysis. PLoS ONE 2014, 9, e109478. [Google Scholar] [CrossRef]
- Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Canto, C.; La Rosa, N.; Cantarella, M.A.; Spampinato, G.; Scalisi, A.; Agodi, A. LINE-1 hypermethylation in white blood cell DNA is associated with high-grade cervical intraepithelial neoplasia. BMC Cancer 2017, 17, 601. [Google Scholar] [CrossRef]
- Bollati, V.; Schwartz, J.; Wright, R.; Litonjua, A.; Tarantini, L.; Suh, H.; Sparrow, D.; Vokonas, P.; Baccarelli, A. Decline in genomic DNA methylation through aging in a cohort of elderly subjects. Mech. Ageing Dev. 2009, 130, 234–239. [Google Scholar] [CrossRef] [Green Version]
- Bollati, V.; Galimberti, D.; Pergoli, L.; Dalla Valle, E.; Barretta, F.; Cortini, F.; Scarpini, E.; Bertazzi, P.A.; Baccarelli, A. DNA methylation in repetitive elements and Alzheimer disease. Brain Behav. Immun. 2011, 25, 1078–1083. [Google Scholar] [CrossRef] [Green Version]
- Carraro, J.C.; Mansego, M.L.; Milagro, F.I.; Chaves, L.O.; Vidigal, F.C.; Bressan, J.; Martínez, J.A. LINE-1 and inflammatory gene methylation levels are early biomarkers of metabolic changes: Association with adiposity. Biomarkers 2016, 21, 625–632. [Google Scholar] [CrossRef]
- Choi, J.Y.; James, S.R.; Link, P.A.; McCann, S.E.; Hong, C.C.; Davis, W.; Nesline, M.K.; Ambrosone, C.B.; Karpf, A.R. Association between global DNA hypomethylation in leukocytes and risk of breast cancer. Carcinogenesis 2009, 30, 1889–1897. [Google Scholar] [CrossRef] [Green Version]
- Maugeri, A.; Barchitta, M.; Fallico, M.; Castellino, N.; Reibaldi, M.; Agodi, A. Characterization of SIRT1/DNMTs functions and LINE-1 methylation in patients with age-related macular degeneration. J. Clin. Med. 2019, 8, 159. [Google Scholar] [CrossRef] [Green Version]
- Carreira, P.E.; Richardson, S.R.; Faulkner, G.J. L1 retrotransposons, cancer stem cells and oncogenesis. FEBS J. 2014, 281, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Barchitta, M.; Maugeri, A.; Li Destri, G.; Basile, G.; Agodi, A. Epigenetic biomarkers in colorectal cancer patients receiving adjuvant or neoadjuvant therapy: A systematic review of epidemiological studies. Int. J. Mol. Sci. 2019, 20, 3842. [Google Scholar] [CrossRef] [Green Version]
- Agodi, A.; Barchitta, M.; Maugeri, A.; Basile, G.; Zamboni, M.; Bernardini, G.; Corona, D.; Veroux, M. Unveiling the role of DNA methylation in kidney transplantation: Novel perspectives toward biomarker identification. Biomed. Res. Int. 2019, 2019, 1602539. [Google Scholar] [CrossRef]
- Puigserver, P.; Wu, Z.; Park, C.W.; Graves, R.; Wright, M.; Spiegelman, B.M. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 1998, 92, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Oberkofler, H.; Krempler, F.; Patsch, W. Human peroxisome proliferator activated receptor gamma coactivator 1 (PPARGC1) gene: cDNA sequence, genomic organization, chromosomal localization, and tissue expression. Genomics 1999, 62, 98–102. [Google Scholar] [CrossRef]
- Mootha, V.K.; Handschin, C.; Arlow, D.; Xie, X.; St. Pierre, J.; Sihag, S.; Yang, W.; Altshuler, D.; Puigserver, P.; Patterson, N.; et al. Erralpha and Gabpa/b specify PGC-1alpha-dependent oxidative phosphorylation gene expression that is altered in diabetic muscle. Proc. Natl. Acad. Sci. USA 2004, 101, 6570–6575. [Google Scholar] [CrossRef] [Green Version]
- Koves, T.R.; Li, P.; An, J.; Akimoto, T.; Slentz, D.; Ilkayeva, O.; Dohm, G.L.; Yan, Z.; Newgard, C.B.; Muoio, D.M. Peroxisome proliferator-activated receptor-gamma co-activator 1alpha-mediated metabolic remodeling of skeletal myocytes mimics exercise training and reverses lipid-induced mitochondrial inefficiency. J. Biol. Chem. 2005, 280, 33588–33598. [Google Scholar] [CrossRef] [Green Version]
- St-Pierre, J.; Drori, S.; Uldry, M.; Silvaggi, J.M.; Rhee, J.; Jäger, S.; Handschin, C.; Zheng, K.; Lin, J.; Yang, W.; et al. Suppression of reactive oxygen species and neurodegeneration by the PGC-1 transcriptional coactivators. Cell 2006, 127, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Yao, M.; Li, J.; Xie, T.; He, T.; Fang, L.; Shi, Y.; Hou, L.; Lian, K.; Wang, R.; Jiang, L. Polymorphisms of rs174616 in the FADS1-FADS2 gene cluster is associated with a reduced risk of type 2 diabetes mellitus in northern Han Chinese people. Diabetes Res. Clin. Pract. 2015, 109, 206–212. [Google Scholar] [CrossRef]
- Abderrahmani, A.; Niederhauser, G.; Plaisance, V.; Roehrich, M.E.; Lenain, V.; Coppola, T.; Regazzi, R.; Waeber, G. Complexin I regulates glucose-induced secretion in pancreatic beta-cells. J. Cell Sci. 2004, 117, 2239–2247. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.M. Leptin and the regulation of body weigh. Keio J. Med. 2011, 60, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Matsuzawa, Y. Adiponectin: A key player in obesity related disorders. Curr. Pharm. Des. 2010, 16, 1896–1901. [Google Scholar] [CrossRef]
- DuBose, A.J.; Johnstone, K.A.; Smith, E.Y.; Hallett, R.A.; Resnick, J.L. Atp10a, a gene adjacent to the PWS/AS gene cluster, is not imprinted in mouse and is insensitive to the PWS-IC. Neurogenetics 2010, 11, 145–151. [Google Scholar] [CrossRef]
- Dhar, M.S.; Sommardahl, C.S.; Kirkland, T.; Nelson, S.; Donnell, R.; Johnson, D.K.; Castellani, L.W. Mice heterozygous for Atp10c, a putative amphipath, represent a novel model of obesity and type 2 diabetes. J. Nutr. 2004, 134, 799–805. [Google Scholar] [CrossRef]
- Satoh, T.; Ichida, T.; Matsuda, Y.; Sugiyama, M.; Yonekura, K.; Ishikawa, T.; Asakura, H. Interaction between hyaluronan and CD44 in the development of dimethylnitrosamine-induced liver cirrhosis. J. Gastroenterol. Hepatol. 2000, 15, 402–411. [Google Scholar] [CrossRef]
- Bertola, A.; Deveaux, V.; Bonnafous, S.; Rousseau, D.; Anty, R.; Wakkach, A.; Dahman, M.; Tordjman, J.; Clément, K.; McQuaid, S.E.; et al. Elevated expression of osteopontin may be related to adipose tissue macrophage accumulation and liver steatosis in morbid obesity. Diabetes 2009, 58, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Han, Y.; Suarez Saiz, F.; Saurez Saiz, F.; Minden, M.D. A tumor suppressor and oncogene: The WT1 story. Leukemia 2007, 21, 868–876. [Google Scholar] [CrossRef]
- Sullivan, K.E.; Reddy, A.B.; Dietzmann, K.; Suriano, A.R.; Kocieda, V.P.; Stewart, M.; Bhatia, M. Epigenetic regulation of tumor necrosis factor alpha. Mol. Cell. Biol. 2007, 27, 5147–5160. [Google Scholar] [CrossRef] [Green Version]
- Tonet, A.C.; Karnikowski, M.; Moraes, C.F.; Gomes, L.; Karnikowski, M.G.; Córdova, C.; Nóbrega, O.T. Association between the -174 G/C promoter polymorphism of the interleukin-6 gene and cardiovascular disease risk factors in Brazilian older women. Braz. J. Med. Biol. Res. 2008, 41, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Moleres, A.; Rendo-Urteaga, T.; Azcona, C.; Martínez, J.A.; Gómez-Martínez, S.; Ruiz, J.R.; Moreno, L.A.; Marcos, A.; Marti, A.; AVENA group. Il6 gene promoter polymorphism (-174G/C) influences the association between fat mass and cardiovascular risk factors. J. Physiol. Biochem. 2009, 65, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.L.; Kratz, M.; Ralston, M.M.; Reinehr, T. Changes in adipose-derived inflammatory cytokines and chemokines after successful lifestyle intervention in obese children. Metabolism 2011, 60, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Bocca, G.; Corpeleijn, E.; Stolk, R.P.; Wolffenbuttel, B.H.; Sauer, P.J. Effect of obesity intervention programs on adipokines, insulin resistance, lipid profile, and low-grade inflammation in 3- to 5-y-old children. Pediatr. Res. 2014, 75, 352–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.F.; Santella, R.M.; Wolff, M.; Kappil, M.A.; Markowitz, S.B.; Morabia, A. White blood cell global methylation and IL-6 promoter methylation in association with diet and lifestyle risk factors in a cancer-free population. Epigenetics 2012, 7, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Pearce, M.S.; McConnell, J.C.; Potter, C.; Barrett, L.M.; Parker, L.; Mathers, J.C.; Relton, C.L. Global LINE-1 DNA methylation is associated with blood glycaemic and lipid profiles. Int. J. Epidemiol. 2012, 41, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Barchitta, M.; Maugeri, A.; Magnano San Lio, R.; Favara, G.; La Rosa, M.C.; La Mastra, C.; Quattrocchi, A.; Agodi, A. Dietary patterns are associated with leukocyte LINE-1 methylation in women: A cross-sectional study in Southern Italy. Nutrients 2019, 11, 1843. [Google Scholar] [CrossRef] [Green Version]
- Barchitta, M.; Maugeri, A.; Quattrocchi, A.; Barone, G.; Mazzoleni, P.; Catalfo, A.; De Guidi, G.; Iemmolo, M.G.; Crimi, N.; Agodi, A. Mediterranean diet and particulate matter exposure are associated with LINE-1 methylation: Results from a cross-sectional study in women. Front. Genet. 2018, 9, 514. [Google Scholar] [CrossRef] [Green Version]
- Maugeri, A.; Barchitta, M.; Magnano San Lio, R.; Favara, G.; La Rosa, M.C.; La Mastra, C.; Basile, G.; Agodi, A. Adherence to the Mediterranean diet partially mediates socioeconomic differences in leukocyte LINE-1 methylation: Evidence from a cross-sectional study in Italian women. Sci. Rep. 2020, 10, 14360. [Google Scholar] [CrossRef]
- Maugeri, A.; Mazzone, M.G.; Giuliano, F.; Vinciguerra, M.; Basile, G.; Barchitta, M.; Agodi, A. Curcumin modulates DNA methyltransferase functions in a cellular model of diabetic retinopathy. Oxid. Med. Cell. Longev. 2018, 2018, 5407482. [Google Scholar] [CrossRef]
- Maugeri, A.; Barchitta, M.; Mazzone, M.G.; Giuliano, F.; Basile, G.; Agodi, A. Resveratrol modulates SIRT1 and DNMT functions and restores LINE-1 methylation levels in ARPE-19 cells under oxidative stress and inflammation. Int. J. Mol. Sci. 2018, 19, 2118. [Google Scholar] [CrossRef] [Green Version]
- Agodi, A.; Barchitta, M.; Quattrocchi, A.; Maugeri, A.; Canto, C.; Marchese, A.E.; Vinciguerra, M. Low fruit consumption and folate deficiency are associated with LINE-1 hypomethylation in women of a cancer-free population. Genes Nutr. 2015, 10, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axume, J.; Smith, S.S.; Pogribny, I.P.; Moriarty, D.J.; Caudill, M.A. Global leukocyte DNA methylation is similar in African American and Caucasian women under conditions of controlled folate intake. Epigenetics 2007, 2, 66–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, W.; Yan, J.; Abratte, C.M.; Vermeylen, F.; Caudill, M.A. Choline intake exceeding current dietary recommendations preserves markers of cellular methylation in a genetic subgroup of folate-compromised men. J. Nutr. 2010, 140, 975–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arpón, A.; Riezu-Boj, J.I.; Milagro, F.I.; Marti, A.; Razquin, C.; Martínez-González, M.A.; Corella, D.; Estruch, R.; Casas, R.; Fitó, M.; et al. Adherence to Mediterranean diet is associated with methylation changes in inflammation-related genes in peripheral blood cells. J. Physiol. Biochem. 2016, 73, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arpón, A.; Milagro, F.I.; Razquin, C.; Corella, D.; Estruch, R.; Fitó, M.; Marti, A.; Martínez-González, M.A.; Ros, E.; Salas-Salvadó, J.; et al. Impact of consuming extra-virgin olive oil or nuts within a Mediterranean diet on DNA methylation in peripheral white blood cells within the PREDIMED-Navarra randomized controlled trial: A Role for dietary lipids. Nutrients 2017, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Druet, C.; Stettler, N.; Sharp, S.; Simmons, R.K.; Cooper, C.; Smith, G.D.; Ekelund, U.; Lévy-Marchal, C.; Jarvelin, M.R.; Kuh, D.; et al. Prediction of childhood obesity by infancy weight gain: An individual-level meta-analysis. Paediatr. Perinat. Epidemiol. 2012, 26, 19–26. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| First Author and Year of Publication | Country | Study Design | Study Population | Age (years) | Dietary Intervention | DNA Methylation Marker | Method | Samples |
|---|---|---|---|---|---|---|---|---|
| Lee 2013 [37] | Mexico | RCT | Pregnant women at 18–22 weeks of gestation | 18–35 | Daily supplementation of 400 mg DHA or a placebo | LINE-1, IFNγ-1, IFNγ-2, TNF-alpha, GATA3, IL10, IL13, STAT3, FOXP3 | Pyrosequencing | Cord blood |
| Geraghty 2018 [38] | Ireland | RCT | The discovery cohort included 60 sex-matched mother-child pairs (30 participants in the intervention arm and 30 participants in the control arm). The replication cohort consisted of different 60 sex-matched mother-child pairs | Mean = 32.8 (SD = 4.5) in the intervention group; 33.9 (SD = 4.2) in the control group | Eucaloric diet but replacing high glycemic index foods with low glycemic index alternatives | Genome-wide methylation profile | Illumina Infinium MethylationEPIC BeadChip Array and Sequenom MassARRAY | Cord blood |
| Brøns 2010 [39] | Denmark | Randomized crossover study | 20 low birthweight men and 26 normal birthweight controls | Mean = 24.6 (SD = 1) and 24.2 (SD = 0.5) in normal birthweight and low birthweight, respectively | Five-day high-fat overfeeding diet and a control diet in a randomized order separated by six to eight weeks | PPARGC1A | Bisulfite sequencing | Skeletal muscle biopsy |
| Gillberg 2014 [40] | Denmark | Randomized crossover study | 19 low birthweight men and 26 normal birthweight controls | 23–27 | Five-day high-fat overfeeding diet and a control diet in a randomized order separated by six to eight weeks | PPARGC1A | Bisulfite sequencing | Subcutaneous adipose tissue |
| Jacobsen 2012 [41] | Denmark | Randomized crossover study | 25 young men with normal birthweight | Mean = 24.6 (SD = 1.1) | Three-day weight-maintaining diet followed by five-day high-fat overfeeding diet | Genomewide methylation profile | Infinium HumanMethylation27K bead chip, Sequenom’s MassARRAY EpiTYPER and pyrosequencing | Skeletal muscle biopsy |
| Jacobsen 2014 [42] | Denmark | Randomized crossover study | 17 low birthweight men and 23 normal birthweight controls | Mean = 24.6 (SD = 1) and 24.2 (SD = 0.5) in normal birthweight and low birthweight, respectively | Five-day high-fat overfeeding diet and a control diet in a randomized order separated by six to eight weeks | Genomewide methylation profile | Infinium HumanMethylation27K bead chip | Skeletal muscle biopsy |
| Gillberg 2016 [43] | Denmark | Randomized crossover study | The discovery cohort included 16 normal birthweight men and 24 age- and BMI-matched control men with normal birthweight. Two replication cohorts consisted of 142 elderly monozygotic and dizygotic twins and 17 healthy young individuals, respectively | NA | Three-day weight-maintaining diet followed by 5-day high-fat overfeeding diet | Genomewide methylation profile | Infinium HumanMethylation450K bead chip | Subcutaneous adipose tissue |
| Hjort 2017 [44] | Denmark | Non-RCT | 21 low birthweight men and 18 normal birthweight controls | Mean = 24.6 (SD = 1.2) and 24.8 (SD = 1.4) in normal birthweight and low birthweight, respectively | 72 h control diet of precooked meals followed by 36 h of fasting with ad libitum water | LEP and ADIPOQ | Sequenom MassARRAY | Subcutaneous adipose tissue |
| Milagro 2011 [22] | Spain | Non-RCT | 25 overweight or obese men | NA | Eight-week energy-restricted diet with 53% of energy from carbohydrates, 17% from proteins and 30% from fats | Genomewide methylation profile | Infinium HumanMethylation27K bead chip and MALDI-TOF mass spectrometry | Blood |
| Samblas 2018 [45] | Spain | RCT | 47 adults with metabolic syndrome randomized to an energy-restricted dietary intervention | NA | Seven meals per day with a macronutrient distribution of 40% total caloric value from carbohydrates, 30% from proteins and 30% from lipids | Genomewide methylation profile | Infinium HumanMethylation27K bead chip and Sequenom’s MassARRAY EpiTYPER | Blood |
| Cordero 2011 [46] | Spain | Non-RCT | 27 obese women | 32–50 | Eight-week energy-restricted diet with 55% of energy from carbohydrates, 15% from proteins and 30% from fats | Leptin and TNF-alpha | Methylation-specific PCR | Adipose tissue |
| Campion 2009 [47] | Spain | Non-RCT | 24 obese individuals | Mean = 34 (SD = 4) | Eight-week energy-restricted diet | TNF-alpha | Bisulfite sequencing | Blood |
| Nicoletti 2015 [48] | Brazil and Spain | Non-RCT | 45 women randomized to three different intervention groups | Mean = 31.7 (SD = 8.6) in the control group; 52.6 (SD = 9.9) in the energy restriction group; 35.5 (SD = 10.1) in the bariatric surgery group | Six-month energy restriction program; hypocaloric dietary treatment followed by bariatric surgery; control group | LINE-1, IL-6, and SERPINE-1 | Methylation-sensitive high-resolution melting analysis | Blood |
| Delgado-Cruzata 2015 [49] | USA | Non-RCT | 24 overweight and sedentary female breast cancer survivors | Mean = 52.2 (SD = 8.7) | Six-month weight loss program aimed to increase physical activity to 90 minutes per week, to reduce caloric intake, and to distribute caloric intake as 45% from protein, 30% from carbohydrates, and 25% from fats | Global DNA methylation and LINE-1 | LUMA, pyrosequencing and MethyLight assay | Blood |
| Garcia-Lacarte 2016 [50] | Spain | RCT | 96 adults with metabolic syndrome randomized to two energy-restricted dietary interventions | NA | Seven meals per day with a macronutrient distribution of 40% total caloric value from carbohydrates, 30% from proteins and 30% from lipids; the control group followed the American Heart Association guidelines | LINE-1 | Methylation-sensitive high-resolution melting analysis | Blood |
| Martin-Nunez 2014 [51] | Spain | Non-RCT | 310 obese participants | Mean = 53.6 and 54.6 in the control and intervention group, respectively | 12-month intervention program based on the promotion of Mediterranean diet and exercise | LINE-1 and SCD1 | Pyrosequencing | Blood |
| Duggan 2014 [52] | USA | RCT | 298 overweight women randomized to four groups | Mean = 57.9 (SD = 4.9) | A control group and 12-month interventions aimed to reduce energy intake, to increase physical activity, or both | LINE-1 | Pyrosequencing | Blood |
| Samblas 2016 [53] | Spain | Non-RCT | 61 overweight or obese women | Mean = 42.2 (SD = 11.4) | Nine-month program to promote Mediterranean diet, physical activity, nutritional and behavioral education | BMAL1, NR1D1, and CLOCK | Sequenom’s MassARRAY EpiTyper | Blood |
| Sun 2018 [54] | USA | RCT | 692 individuals randomized to four energy-reduced diets varying in macronutrients | Mean = 51.4 | Two-year interventions, consisting of two high-fat diets low in carbohydrate and two low-fat diets high in carbohydrate diets | Genomewide methylation profile | Infinium HumanMethylation450 BeadChip | Blood |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maugeri, A. The Effects of Dietary Interventions on DNA Methylation: Implications for Obesity Management. Int. J. Mol. Sci. 2020, 21, 8670. https://doi.org/10.3390/ijms21228670
Maugeri A. The Effects of Dietary Interventions on DNA Methylation: Implications for Obesity Management. International Journal of Molecular Sciences. 2020; 21(22):8670. https://doi.org/10.3390/ijms21228670
Chicago/Turabian StyleMaugeri, Andrea. 2020. "The Effects of Dietary Interventions on DNA Methylation: Implications for Obesity Management" International Journal of Molecular Sciences 21, no. 22: 8670. https://doi.org/10.3390/ijms21228670
APA StyleMaugeri, A. (2020). The Effects of Dietary Interventions on DNA Methylation: Implications for Obesity Management. International Journal of Molecular Sciences, 21(22), 8670. https://doi.org/10.3390/ijms21228670