Effects of Tenascin C on the Integrity of Extracellular Matrix and Skin Aging

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
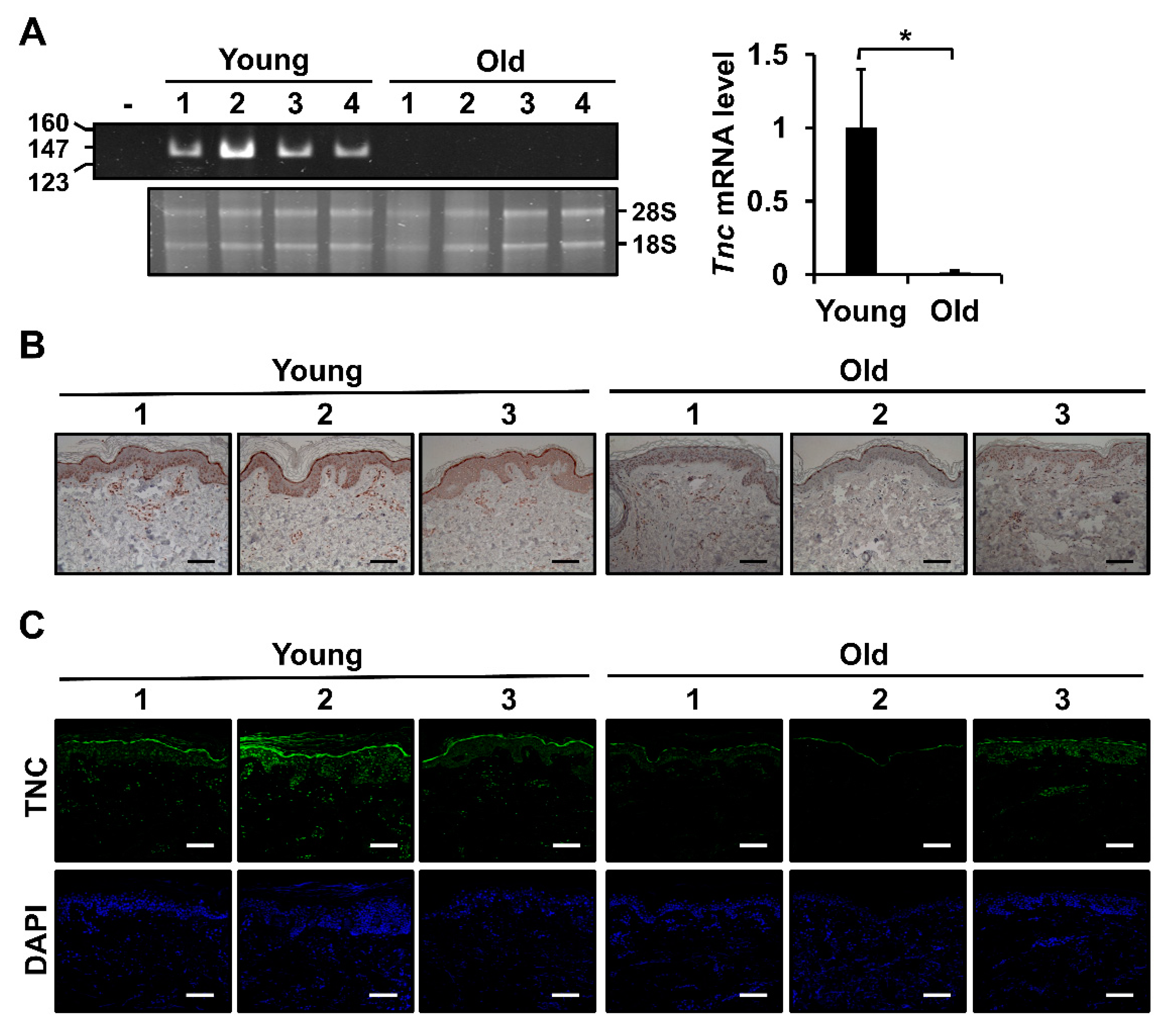
2.1. Expression of TNC is Downregulated during Intrinsic Skin Aging in Mouse and Human Skin Tissues
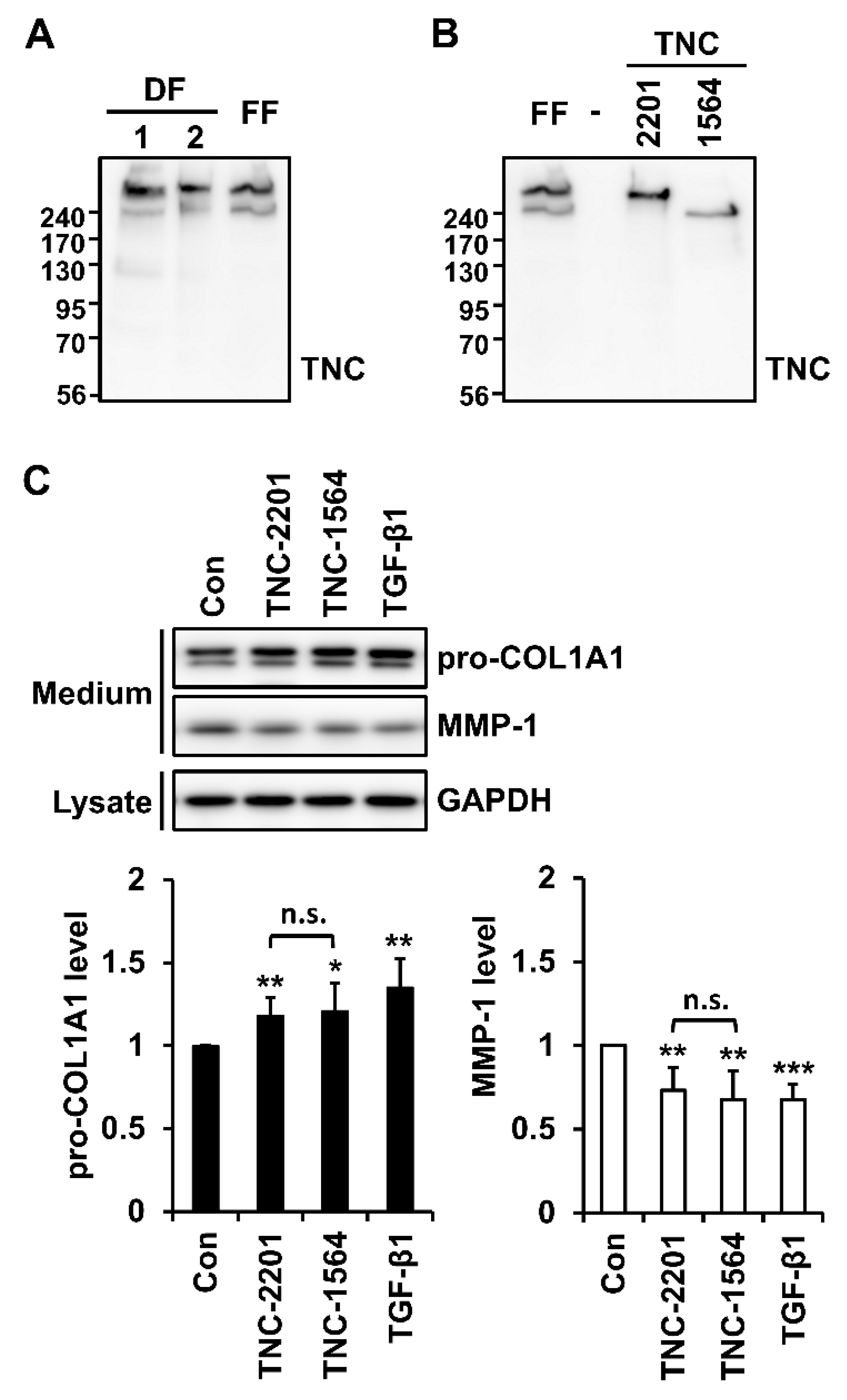
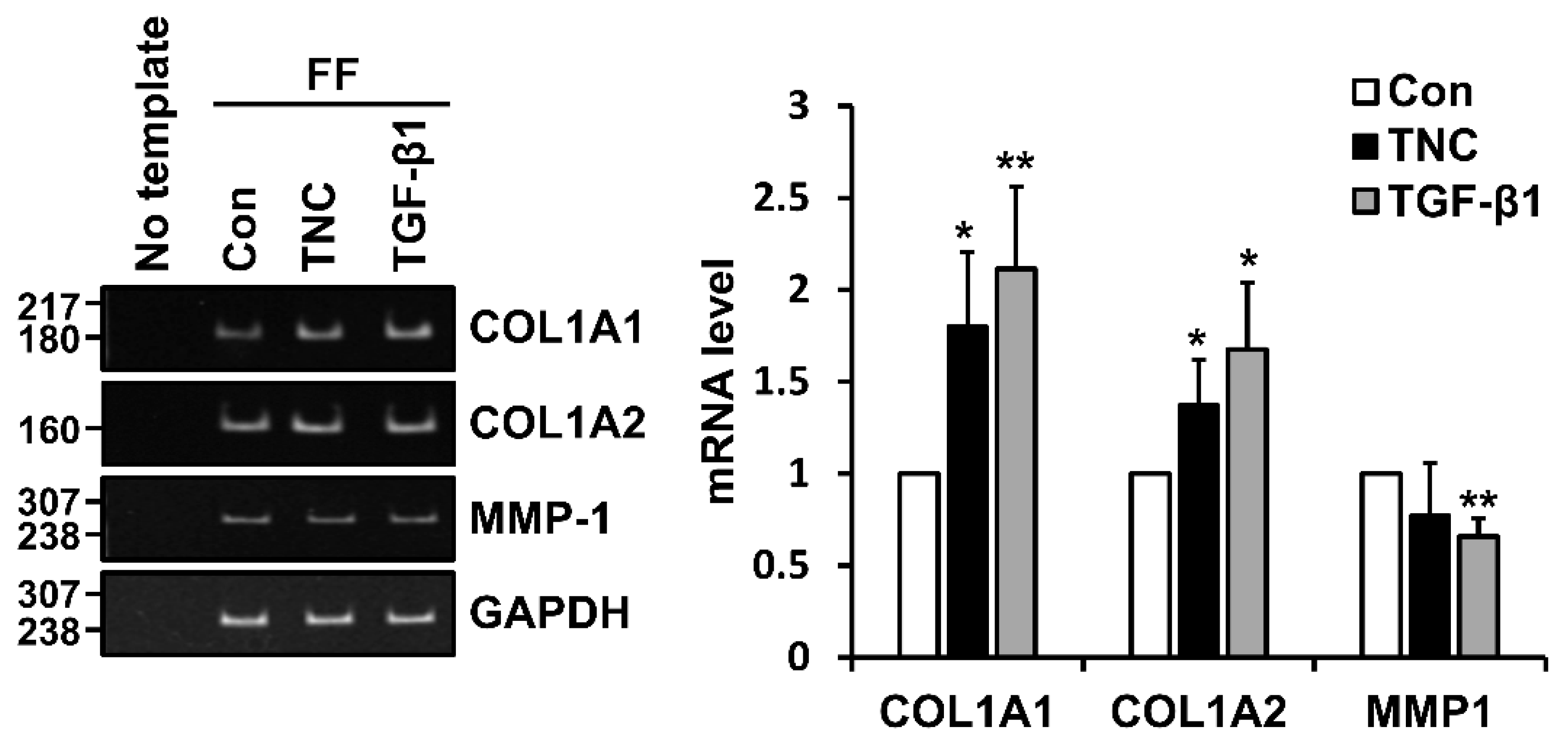
2.2. TNC Upregulates Type I Collagen Expression and Downregulates MMP-1 Expression in Fibroblasts
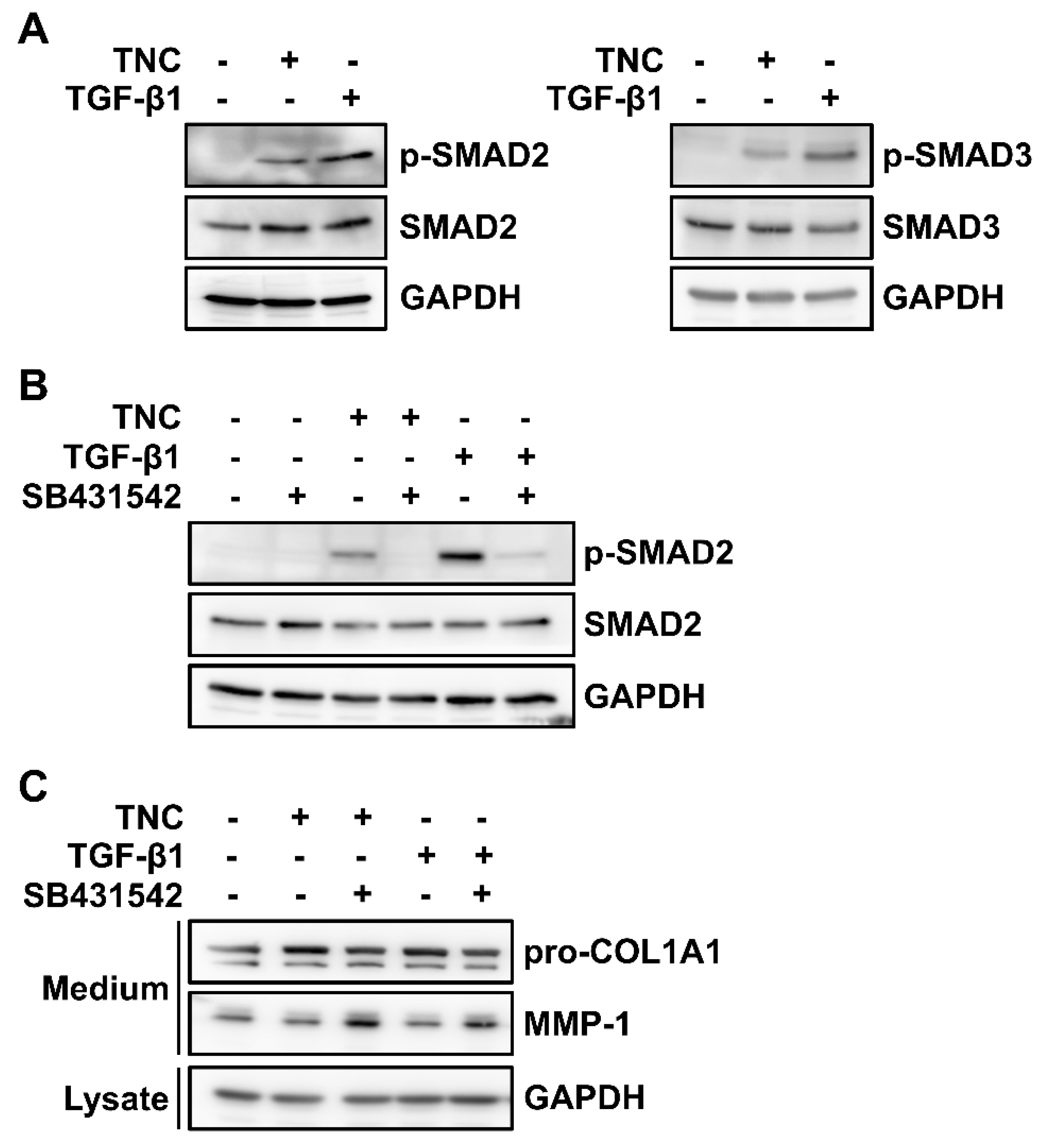
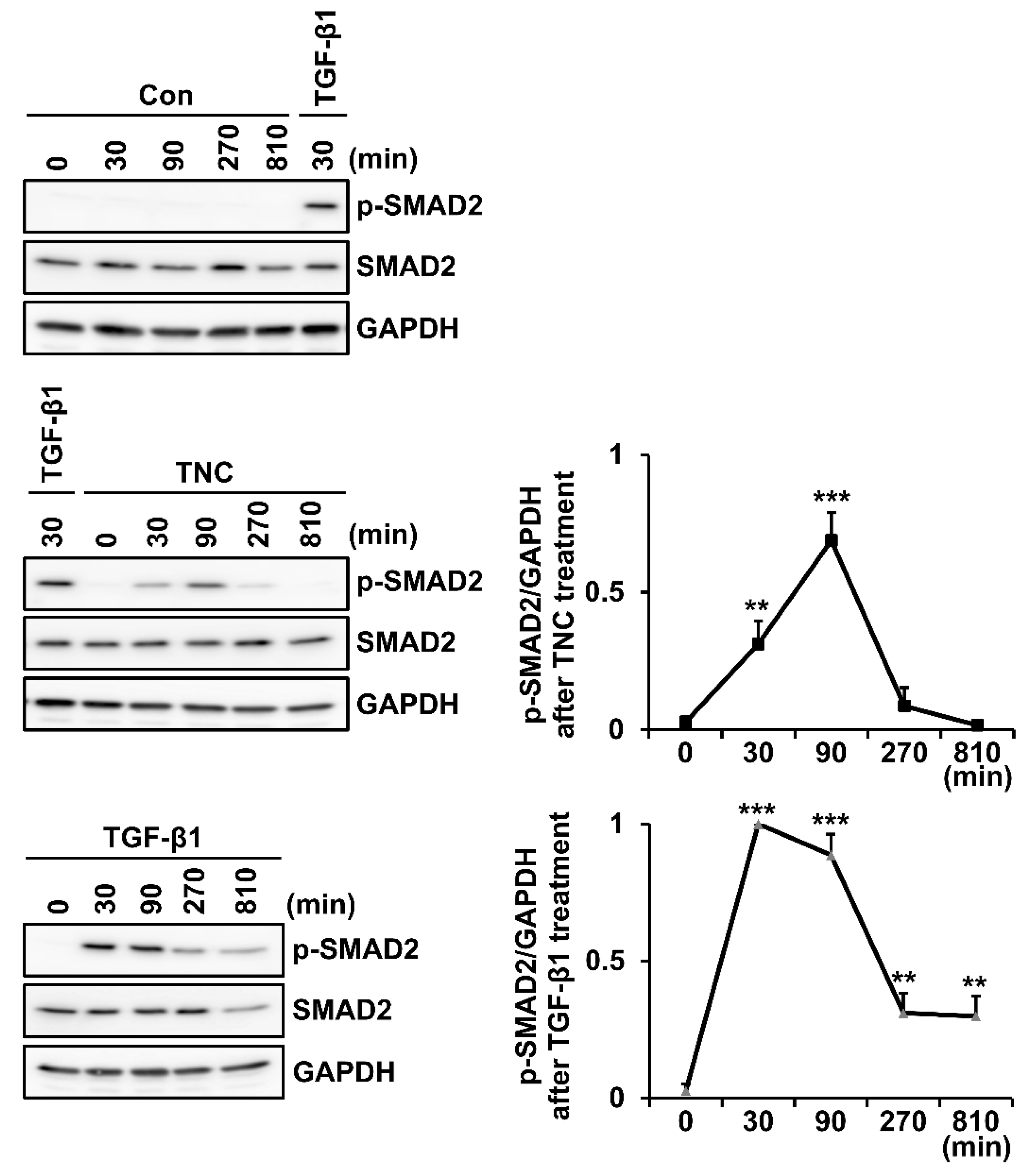
2.3. TNC Activates Receptor-Regulated SMADs (R-SMADs) and TGF-β Receptors in Fibroblasts
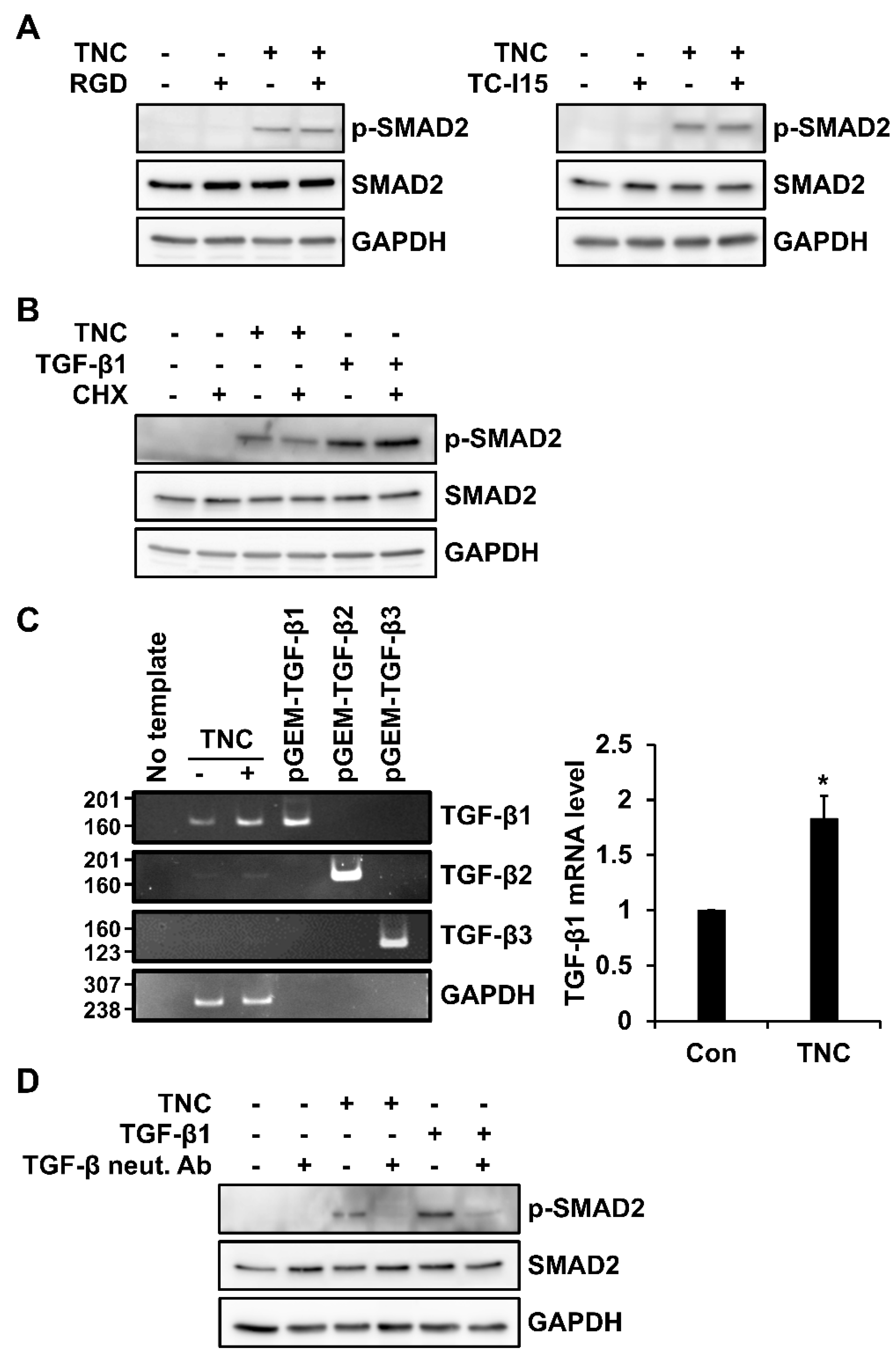
2.4. TNC Induces TGF-β Family Members in Fibroblasts
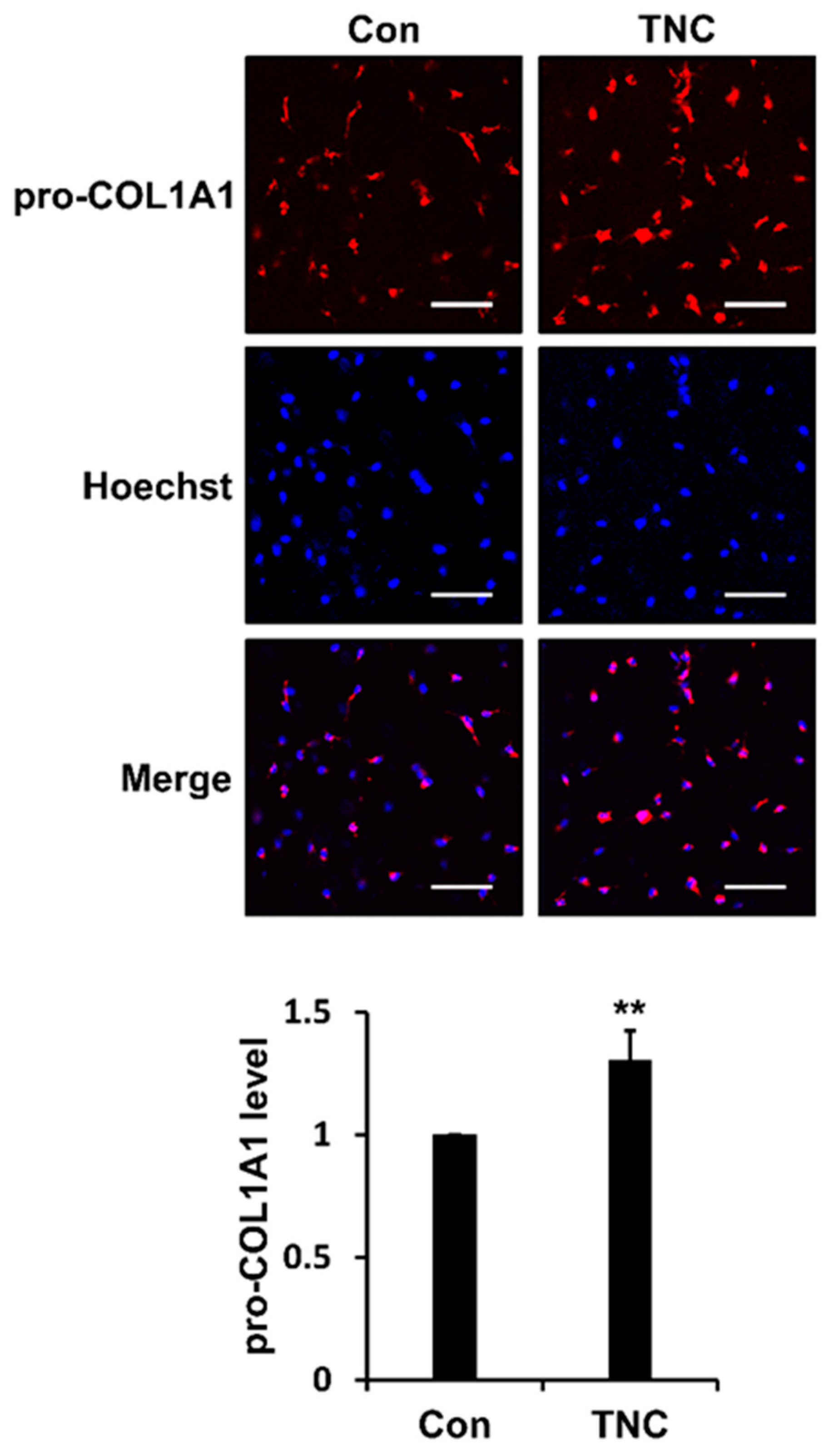
2.5. TNC Increases the Biosynthesis of Type I Collagen during 3D Culture of Fibroblasts
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture
4.3. Acquisition of Mouse and Human Skin Tissues
4.4. RNA Isolation and RT-PCR Analysis
4.5. Histological Analysis
4.6. Western Blot Analysis
4.7. Construction of TNC Expression Vectors
4.8. Purification of Recombinant Human TNC
4.9. Construction of pGEM-TGF-β1, 2, and 3 Vectors
4.10. Treatment of Fibroblasts with TNC or TGF-β1 in the Presence or Absence of Signaling Blockers
4.11. Synthesis of Type I Collagen in Fibroblasts Grown in 3D Culture
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | Three-dimensional |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMEM | Dulbecco’s modified Eagle’s medium |
| ECM | Extracellular matrix |
| EGF | Epidermal growth factor |
| MMP | Matrix metalloproteinase |
| IF | Immunofluorescence |
| IHC | Immunohistochemistry |
| KO | Knockout |
| R-SMAD | Receptor-regulated SMAD |
| RT-PCR | Reverse transcription-polymerase chain reaction |
| TGF-β | Transforming growth factor-β |
| TNC | Tenascin C |
| WT | Wild-type |
References
- Midwood, K.S.; Chiquet, M.; Tucker, R.P.; Orend, G. Tenascin-C at a glance. J. Cell Sci. 2016, 129, 4321–4327. [Google Scholar] [CrossRef] [PubMed]
- Berndt, A.; Richter, P.; Kosmehl, H.; Franz, M. Tenascin-C and carcinoma cell invasion in oral and urinary bladder cancer. Cell Adhes. Migr. 2015, 9, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Niebroj-Dobosz, I. Tenascin-C in human cardiac pathology. Clin. Chim. Acta 2012, 413, 1516–1518. [Google Scholar] [CrossRef] [PubMed]
- Brellier, F.; Tucker, R.P.; Chiquet-Ehrismann, R. Tenascins and their implications in diseases and tissue mechanics. Scand. J. Med. Sci. Sports 2009, 19, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.Y.; Lee, K.W.; Shin, D.; An, S.; Cho, K.H.; Kim, S.H. A positive feedback loop bi-stably activates fibroblasts. Nat. Commun. 2018, 9, 3016. [Google Scholar] [CrossRef]
- Imanaka-Yoshida, K.; Tawara, I.; Yoshida, T. Tenascin-C in cardiac disease: A sophisticated controller of inflammation, repair, and fibrosis. Am. J. Physiol. Cell Physiol. 2020, 319, C781–C796. [Google Scholar] [CrossRef]
- Murphy-Ullrich, J.E.; Sage, E.H. Revisiting the matricellular concept. Matrix Biol. 2014, 37, 1–14. [Google Scholar] [CrossRef]
- Swindle, C.S.; Tran, K.T.; Johnson, T.D.; Banerjee, P.; Mayes, A.M.; Griffith, L.; Wells, A. Epidermal growth factor (EGF)-like repeats of human tenascin-C as ligands for EGF receptor. J. Cell Biol. 2001, 154, 459–468. [Google Scholar] [CrossRef]
- Yoshida, T.; Akatsuka, T.; Imanaka-Yoshida, K. Tenascin-C and integrins in cancer. Cell Adhes. Migr. 2015, 9, 96–104. [Google Scholar] [CrossRef]
- Tucker, R.P.; Chiquet-Ehrismann, R. Tenascin-C: Its functions as an integrin ligand. Int. J. Biochem. Cell Biol. 2015, 65, 165–168. [Google Scholar] [CrossRef]
- Midwood, K.S.; Hussenet, T.; Langlois, B.; Orend, G. Advances in tenascin-C biology. Cell. Mol. Life Sci. 2011, 68, 3175–3199. [Google Scholar] [CrossRef] [PubMed]
- Paron, I.; Berchtold, S.; Voros, J.; Shamarla, M.; Erkan, M.; Hofler, H.; Esposito, I. Tenascin-C enhances pancreatic cancer cell growth and motility and affects cell adhesion through activation of the integrin pathway. PLoS ONE 2011, 6, e21684. [Google Scholar] [CrossRef] [PubMed]
- Katoh, D.; Nagaharu, K.; Shimojo, N.; Hanamura, N.; Yamashita, M.; Kozuka, Y.; Imanaka-Yoshida, K.; Yoshida, T. Binding of alphavbeta1 and alphavbeta6 integrins to tenascin-C induces epithelial-mesenchymal transition-like change of breast cancer cells. Oncogenesis 2013, 2, e65. [Google Scholar] [CrossRef] [PubMed]
- Lange, K.; Kammerer, M.; Hegi, M.E.; Grotegut, S.; Dittmann, A.; Huang, W.; Fluri, E.; Yip, G.W.; Gotte, M.; Ruiz, C.; et al. Endothelin receptor type B counteracts tenascin-C-induced endothelin receptor type A-dependent focal adhesion and actin stress fiber disorganization. Cancer Res. 2007, 67, 6163–6173. [Google Scholar] [CrossRef]
- Katoh, D.; Kozuka, Y.; Noro, A.; Ogawa, T.; Imanaka-Yoshida, K.; Yoshida, T. Tenascin-C induces phenotypic changes in fibroblasts to myofibroblasts with high contractility through the integrin alphavbeta1/transforming growth factor beta/SMAD signaling axis in human breast cancer. Am. J. Pathol. 2020, 190, 2123–2135. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Wang, W.; Morales-Nebreda, L.; Feng, G.; Wu, M.; Zhou, X.; Lafyatis, R.; Lee, J.; Hinchcliff, M.; Feghali-Bostwick, C.; et al. Tenascin-C drives persistence of organ fibrosis. Nat. Commun. 2016, 7, 11703. [Google Scholar] [CrossRef]
- Ma, J.C.; Huang, X.; Shen, Y.W.; Zheng, C.; Su, Q.H.; Xu, J.K.; Zhao, J. Tenascin-C promotes migration of hepatic stellate cells and production of type I collagen. Biosci. Biotechnol. Biochem. 2016, 80, 1470–1477. [Google Scholar] [CrossRef]
- Kasprzycka, M.; Hammarstrom, C.; Haraldsen, G. Tenascins in fibrotic disorders-from bench to bedside. Cell Adhes. Migr. 2015, 9, 83–89. [Google Scholar] [CrossRef]
- Quan, T.; Shao, Y.; He, T.; Voorhees, J.J.; Fisher, G.J. Reduced expression of connective tissue growth factor (CTGF/CCN2) mediates collagen loss in chronologically aged human skin. J. Investig. Dermatol. 2010, 130, 415–424. [Google Scholar] [CrossRef]
- El-Karef, A.; Yoshida, T.; Gabazza, E.C.; Nishioka, T.; Inada, H.; Sakakura, T.; Imanaka-Yoshida, K. Deficiency of tenascin-C attenuates liver fibrosis in immune-mediated chronic hepatitis in mice. J. Pathol. 2007, 211, 86–94. [Google Scholar] [CrossRef]
- Carey, W.A.; Taylor, G.D.; Dean, W.B.; Bristow, J.D. Tenascin-C deficiency attenuates TGF-ss-mediated fibrosis following murine lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 299, L785–L793. [Google Scholar] [CrossRef] [PubMed]
- Jinnin, M.; Ihn, H.; Asano, Y.; Yamane, K.; Trojanowska, M.; Tamaki, K. Upregulation of tenascin-C expression by IL-13 in human dermal fibroblasts via the phosphoinositide 3-kinase/Akt and the protein kinase C signaling pathways. J. Investig. Dermatol. 2006, 126, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Aukhil, I.; Joshi, P.; Yan, Y.; Erickson, H.P. Cell- and heparin-binding domains of the hexabrachion arm identified by tenascin expression proteins. J. Biol. Chem. 1993, 268, 2542–2553. [Google Scholar] [PubMed]
- Khan, Z.; Marshall, J.F. The role of integrins in TGFbeta activation in the tumour stroma. Cell Tissue Res. 2016, 365, 657–673. [Google Scholar] [CrossRef]
- Ogawa, K.; Ito, M.; Takeuchi, K.; Nakada, A.; Heishi, M.; Suto, H.; Mitsuishi, K.; Sugita, Y.; Ogawa, H.; Ra, C. Tenascin-C is upregulated in the skin lesions of patients with atopic dermatitis. J. Dermatol. Sci. 2005, 40, 35–41. [Google Scholar] [CrossRef]
- Latijnhouwers, M.A.; de Jongh, G.J.; Bergers, M.; de Rooij, M.J.; Schalkwijk, J. Expression of tenascin-C splice variants by human skin cells. Arch. Dermatol. Res. 2000, 292, 446–454. [Google Scholar] [CrossRef]
- Chung, J.H.; Seo, J.Y.; Choi, H.R.; Lee, M.K.; Youn, C.S.; Rhie, G.; Cho, K.H.; Kim, K.H.; Park, K.C.; Eun, H.C. Modulation of skin collagen metabolism in aged and photoaged human skin in vivo. J. Investig. Dermatol. 2001, 117, 1218–1224. [Google Scholar] [CrossRef]
- Latijnhouwers, M.; Bergers, M.; Ponec, M.; Dijkman, H.; Andriessen, M.; Schalkwijk, J. Human epidermal keratinocytes are a source of tenascin-C during wound healing. J. Investig. Dermatol. 1997, 108, 776–783. [Google Scholar] [CrossRef][Green Version]
- Chung, C.Y.; Murphy-Ullrich, J.E.; Erickson, H.P. Mitogenesis, cell migration, and loss of focal adhesions induced by tenascin-C interacting with its cell surface receptor, annexin II. Mol. Biol. Cell 1996, 7, 883–892. [Google Scholar] [CrossRef]
- Orend, G.; Huang, W.; Olayioye, M.A.; Hynes, N.E.; Chiquet-Ehrismann, R. Tenascin-C blocks cell-cycle progression of anchorage-dependent fibroblasts on fibronectin through inhibition of syndecan-4. Oncogene 2003, 22, 3917–3926. [Google Scholar] [CrossRef]
- Chiquet-Ehrismann, R.; Matsuoka, Y.; Hofer, U.; Spring, J.; Bernasconi, C.; Chiquet, M. Tenascin variants: Differential binding to fibronectin and distinct distribution in cell cultures and tissues. Cell Regul. 1991, 2, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Ghert, M.A.; Qi, W.N.; Erickson, H.P.; Block, J.A.; Scully, S.P. Tenascin-C splice variant adhesive/anti-adhesive effects on chondrosarcoma cell attachment to fibronectin. Cell Struct. Funct. 2001, 26, 179–187. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Prieto, A.L.; Andersson-Fisone, C.; Crossin, K.L. Characterization of multiple adhesive and counteradhesive domains in the extracellular matrix protein cytotactin. J. Cell Biol. 1992, 119, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Saga, Y.; Yagi, T.; Ikawa, Y.; Sakakura, T.; Aizawa, S. Mice develop normally without tenascin. Genes Dev. 1992, 6, 1821–1831. [Google Scholar] [CrossRef]
- Forsberg, E.; Hirsch, E.; Frohlich, L.; Meyer, M.; Ekblom, P.; Aszodi, A.; Werner, S.; Fassler, R. Skin wounds and severed nerves heal normally in mice lacking tenascin-C. Proc. Natl. Acad. Sci. USA 1996, 93, 6594–6599. [Google Scholar] [CrossRef]
- Song, L.; Wang, L.; Li, F.; Yukht, A.; Qin, M.; Ruther, H.; Yang, M.; Chaux, A.; Shah, P.K.; Sharifi, B.G. Bone marrow-derived tenascin-C attenuates cardiac hypertrophy by controlling inflammation. J. Am. Coll. Cardiol. 2017, 70, 1601–1615. [Google Scholar] [CrossRef]
- Xie, K.; Liu, Y.; Hao, W.; Walter, S.; Penke, B.; Hartmann, T.; Schachner, M.; Fassbender, K. Tenascin-C deficiency ameliorates Alzheimer’s disease-related pathology in mice. Neurobiol. Aging 2013, 34, 2389–2398. [Google Scholar] [CrossRef]
- Shimojo, N.; Hashizume, R.; Kanayama, K.; Hara, M.; Suzuki, Y.; Nishioka, T.; Hiroe, M.; Yoshida, T.; Imanaka-Yoshida, K. Tenascin-C may accelerate cardiac fibrosis by activating macrophages via the integrin alphaVbeta3/nuclear factor-kappaB/interleukin-6 axis. Hypertension 2015, 66, 757–766. [Google Scholar] [CrossRef]
- Munger, J.S.; Huang, X.; Kawakatsu, H.; Griffiths, M.J.; Dalton, S.L.; Wu, J.; Pittet, J.F.; Kaminski, N.; Garat, C.; Matthay, M.A.; et al. The integrin alpha v beta 6 binds and activates latent TGF beta 1: A mechanism for regulating pulmonary inflammation and fibrosis. Cell 1999, 96, 319–328. [Google Scholar] [CrossRef]
- Hayashida, T.; Wu, M.H.; Pierce, A.; Poncelet, A.C.; Varga, J.; Schnaper, H.W. MAP-kinase activity necessary for TGFbeta1-stimulated mesangial cell type I collagen expression requires adhesion-dependent phosphorylation of FAK tyrosine 397. J. Cell Sci. 2007, 120, 4230–4240. [Google Scholar] [CrossRef]
- White, L.A.; Mitchell, T.I.; Brinckerhoff, C.E. Transforming growth factor beta inhibitory element in the rabbit matrix metalloproteinase-1 (collagenase-1) gene functions as a repressor of constitutive transcription. Biochim. Biophys. Acta 2000, 1490, 259–268. [Google Scholar] [CrossRef]
- Galoian, K.A.; Garamszegi, N.; Garamszegi, S.P.; Scully, S.P. Molecular mechanism of tenascin-C action on matrix metalloproteinase-1 invasive potential. Exp. Biol. Med. 2007, 232, 515–522. [Google Scholar]
- Rogers, N.K.; Clements, D.; Dongre, A.; Harrison, T.W.; Shaw, D.; Johnson, S.R. Extra-cellular matrix proteins induce matrix metalloproteinase-1 (MMP-1) activity and increase airway smooth muscle contraction in asthma. PLoS ONE 2014, 9, e90565. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J. Introduction to skin aging. J. Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell Biol. 2012, 212, 1–115. [Google Scholar]
- Bocheva, G.; Slominski, R.M.; Slominski, A.T. Neuroendocrine aspects of skin aging. Int. J. Mol. Sci. 2019, 20, 2798. [Google Scholar] [CrossRef]
- Sumioka, T.; Kitano, A.; Flanders, K.C.; Okada, Y.; Yamanaka, O.; Fujita, N.; Iwanishi, H.; Kao, W.W.; Saika, S. Impaired cornea wound healing in a tenascin C-deficient mouse model. Lab. Investig. 2013, 93, 207–217. [Google Scholar] [CrossRef]
- Brellier, F.; Martina, E.; Chiquet, M.; Ferralli, J.; van der Heyden, M.; Orend, G.; Schittny, J.C.; Chiquet-Ehrismann, R.; Tucker, R.P. The adhesion modulating properties of tenascin-W. Int. J. Biol. Sci. 2012, 8, 187–194. [Google Scholar] [CrossRef]
- Kim, E.J.; Kim, Y.K.; Kim, M.K.; Kim, S.; Kim, J.Y.; Lee, D.H.; Chung, J.H. UV-induced inhibition of adipokine production in subcutaneous fat aggravates dermal matrix degradation in human skin. Sci. Rep. 2016, 6, 25616. [Google Scholar] [CrossRef]
- Shin, J.E.; Oh, J.H.; Kim, Y.K.; Jung, J.Y.; Chung, J.H. Transcriptional regulation of proteoglycans and glycosaminoglycan chain-synthesizing glycosyltransferases by UV irradiation in cultured human dermal fibroblasts. J. Korean Med. Sci. 2011, 26, 417–424. [Google Scholar] [CrossRef]
- Shin, W.S.; Park, M.K.; Lee, Y.H.; Kim, K.W.; Lee, H.; Lee, S.-T. The catalytically defective receptor protein tyrosine kinase EphA10 promotes tumorigenesis in pancreatic cancer cells. Cancer Sci. 2020, 111, 3292–3302. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.S.; Lee, H.W.; Lee, S.-T. Catalytically inactive receptor tyrosine kinase PTK7 activates FGFR1 independent of FGF. FASEB J. 2019, 33, 12960–12971. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Seo, E.K.; Lee, S.-T. Skullcapflavone II inhibits degradation of type I collagen by suppressing MMP-1 transcription in human skin fibroblasts. Int. J. Mol. Sci. 2019, 20, 2734. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.E.; Song, M.J.; Hara, M.; Imanaka-Yoshida, K.; Lee, D.H.; Chung, J.H.; Lee, S.-T. Effects of Tenascin C on the Integrity of Extracellular Matrix and Skin Aging. Int. J. Mol. Sci. 2020, 21, 8693. https://doi.org/10.3390/ijms21228693
Choi YE, Song MJ, Hara M, Imanaka-Yoshida K, Lee DH, Chung JH, Lee S-T. Effects of Tenascin C on the Integrity of Extracellular Matrix and Skin Aging. International Journal of Molecular Sciences. 2020; 21(22):8693. https://doi.org/10.3390/ijms21228693
Chicago/Turabian StyleChoi, Young Eun, Min Ji Song, Mari Hara, Kyoko Imanaka-Yoshida, Dong Hun Lee, Jin Ho Chung, and Seung-Taek Lee. 2020. "Effects of Tenascin C on the Integrity of Extracellular Matrix and Skin Aging" International Journal of Molecular Sciences 21, no. 22: 8693. https://doi.org/10.3390/ijms21228693
APA StyleChoi, Y. E., Song, M. J., Hara, M., Imanaka-Yoshida, K., Lee, D. H., Chung, J. H., & Lee, S.-T. (2020). Effects of Tenascin C on the Integrity of Extracellular Matrix and Skin Aging. International Journal of Molecular Sciences, 21(22), 8693. https://doi.org/10.3390/ijms21228693