Fatty-Acid-Binding Protein 4 as a Novel Contributor to Mononuclear Cell Activation and Endothelial Cell Dysfunction in Atherosclerosis


Abstract
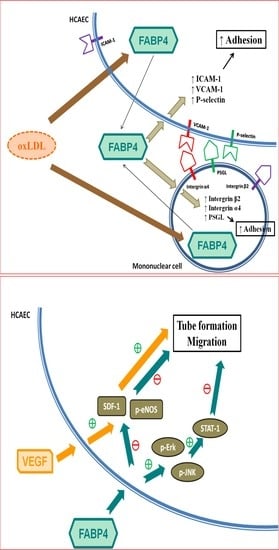
:
1. Introduction
2. Results
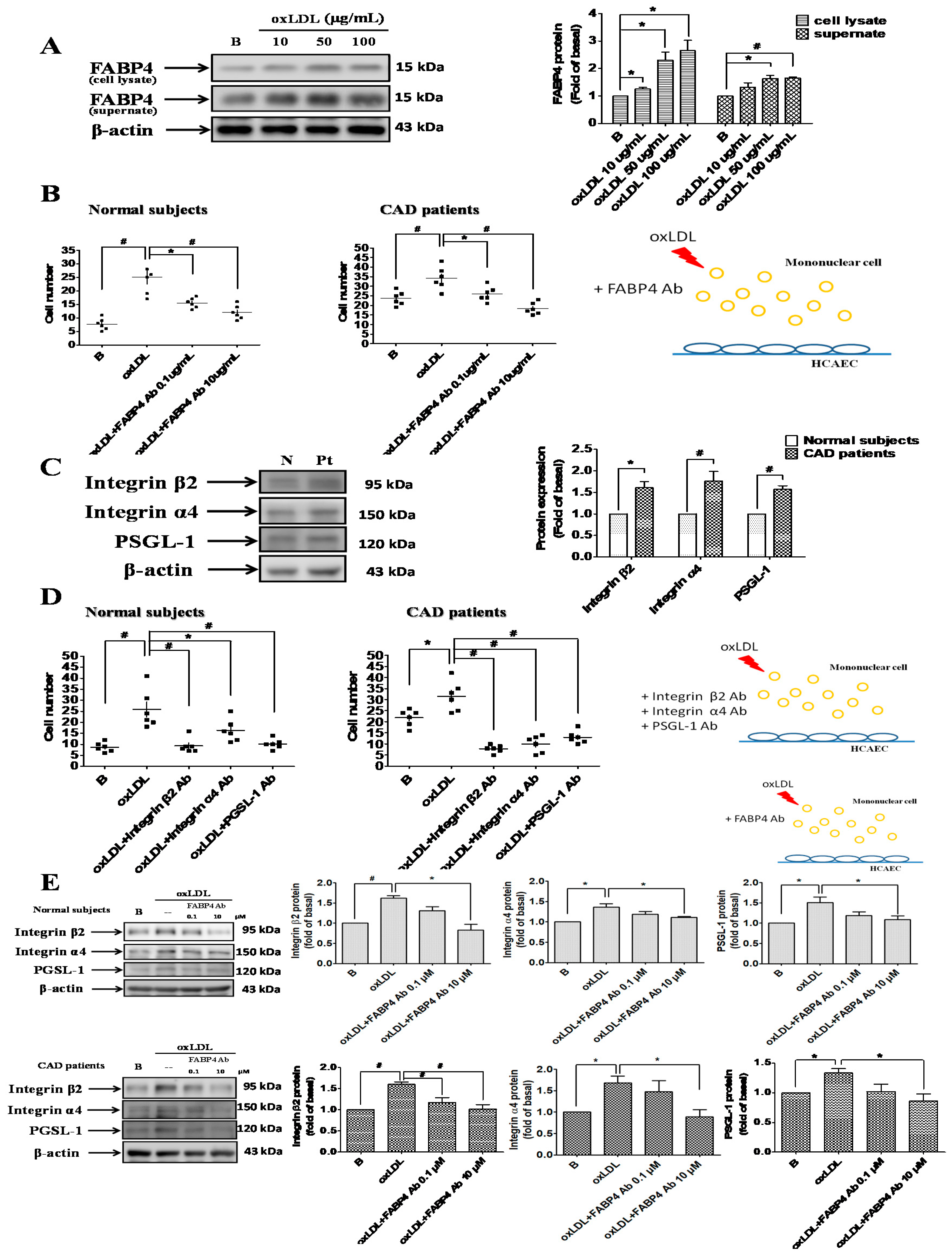
2.1. Up-Regulated FABP4 Secretion and Enhanced Endothelial Adhesiveness of MNCs from CAD Patients
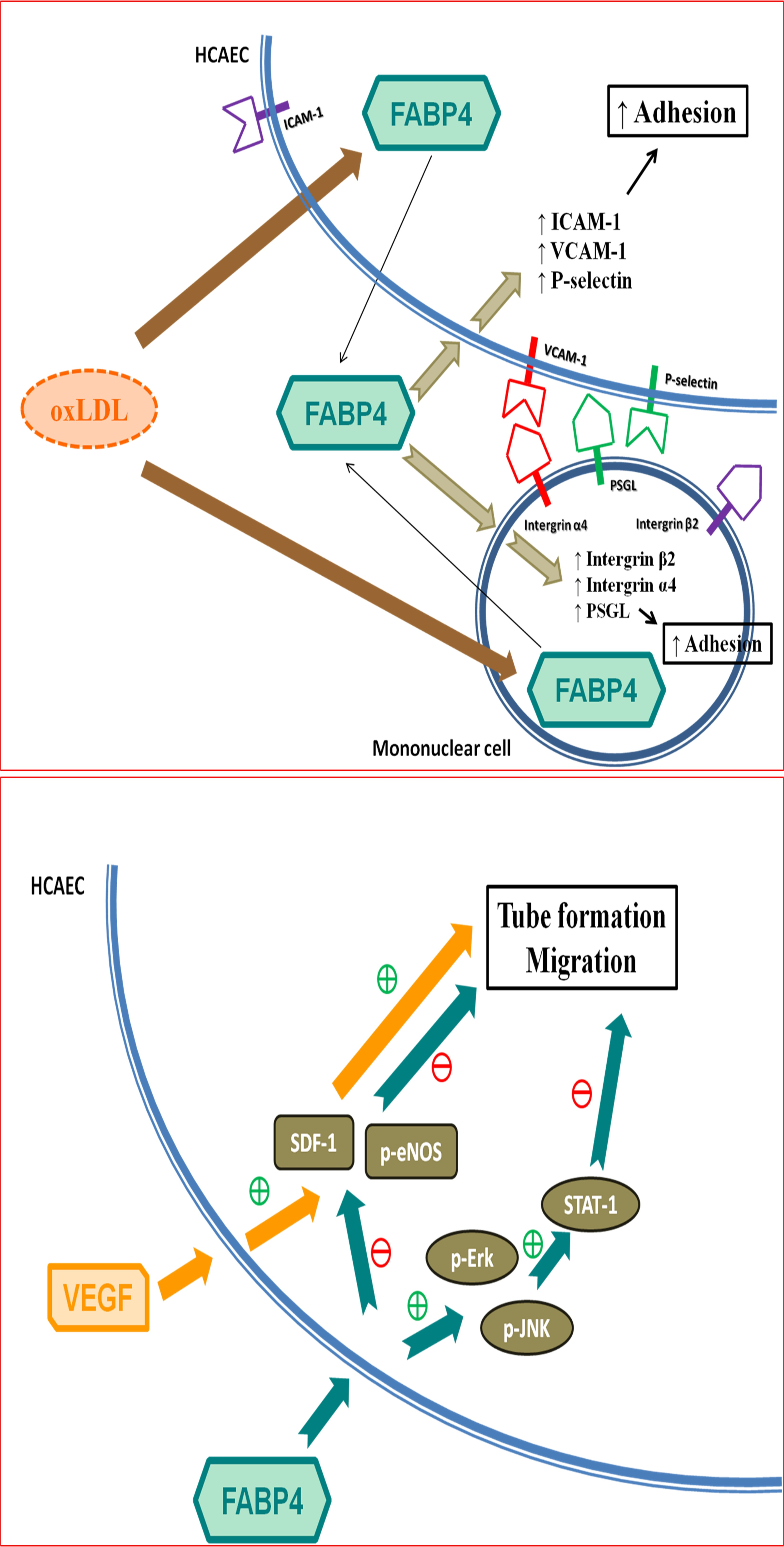
2.2. FABP4 Inhibition Decreased the Oxidized-LDL-Induced Adhesion of MNCs Via Down-Regulating the Integrin β2, Integrin α4, and PSGL-1 Expression
2.3. FABP4 Inhibition Decreased the Oxidized-LDL-Induced Adhesion of HCAECs Via Down-Regulating the ICAM-1, VCAM-1, and P-Selectin Expression
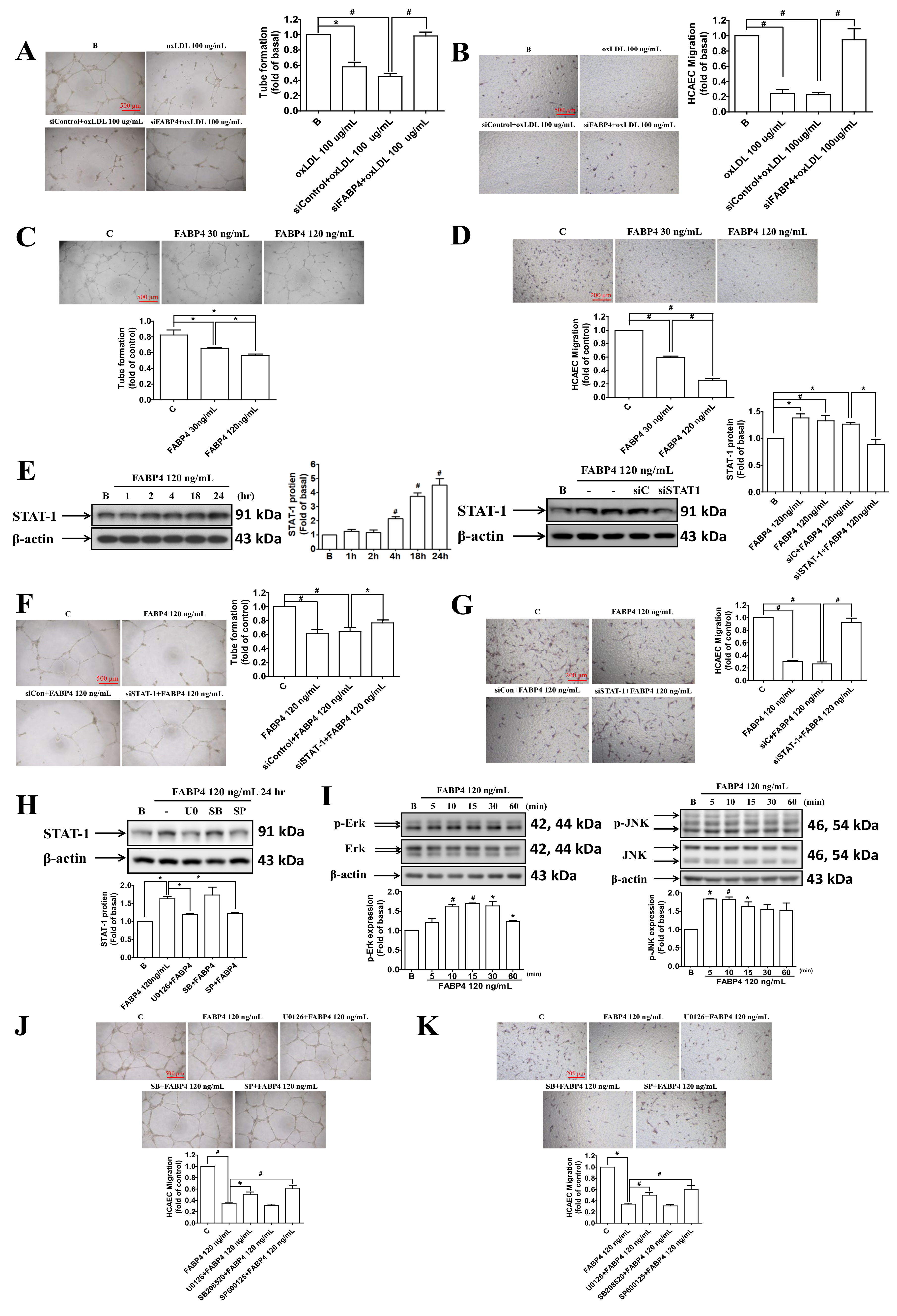
2.4. FABP4 Impaired the Functions of HCAECs via the ERK/JNK/STAT-1 Signaling Pathways
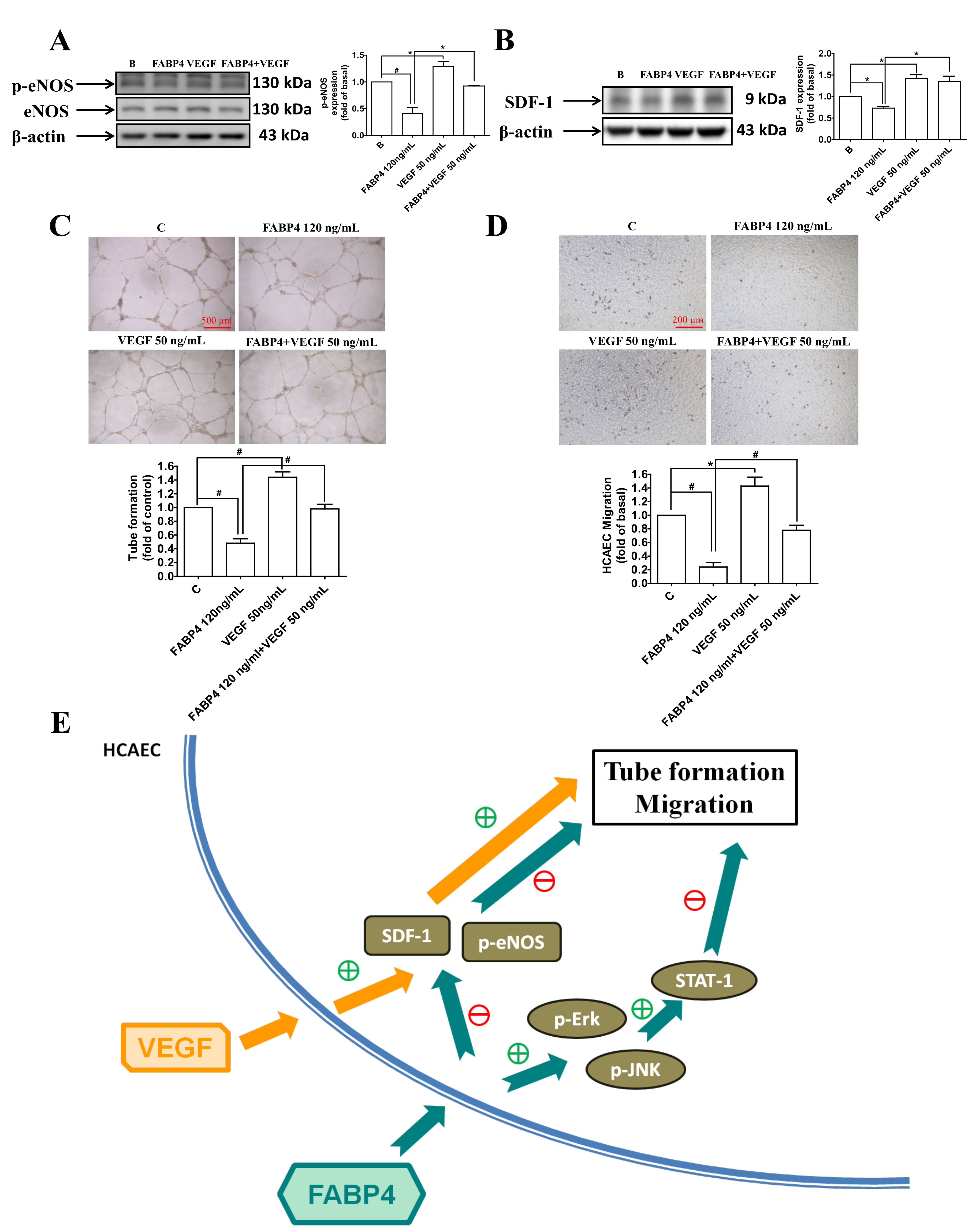
2.5. VEGF Reversed the FABP4-Imapired Functions of HCAECs via Up-Regulating the eNOS Phosphorylation and SDF-1 Expression
3. Discussion
4. Materials and Methods
4.1. Patients and Study Design
4.2. Characteristics of Study Population
4.3. Blood Sample Collections
4.4. Cell Culture
4.5. Measurement of FABP4 Concentrations in Plasma and in Culture Supernatants of Mononuclear Cells from Subjects
4.6. Western Blotting
4.7. Cellular Viability Assay
4.8. Tube Formation Assay
4.9. Migration Assay
4.10. The Transfection of FABP4 or STAT-1 siRNA
4.11. Adhesion Assay
4.12. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Xu, A.; Wang, Y.; Xu, J.Y.; Stejskal, D.; Tam, S.; Zhang, J.; Wat, N.M.; Wong, W.K.; Lam, K.S. Adipocyte fatty acid-binding protein is a plasma biomarker closely associated with obesity and metabolic syndrome. Clin. Chem. 2006, 52, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Terra, X.; Quintero, Y.; Auguet, T.; Porras, J.A.; Hernández, M.; Sabench, F.; Aguilar, C.; Luna, A.M.; Del Castillo, D.; Richart, C. FABP 4 is associated with inflammatory markers and metabolic syndrome in morbidly obese women. Eur. J. Endocrinol. 2011, 164, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuhashi, M.; Ishimura, S.; Ota, H.; Hayashi, M.; Nishitani, T.; Tanaka, M.; Yoshida, H.; Shimamoto, K.; Hotamisligil, G.S.; Miura, T. Serum fatty acid-binding protein 4 is a predictor of cardiovascular events in end-stage renal disease. PLoS ONE 2011, 6, e27356. [Google Scholar] [CrossRef] [PubMed]
- Ota, H.; Furuhashi, M.; Ishimura, S.; Koyama, M.; Okazaki, Y.; Mita, T.; Fuseya, T.; Yamashita, T.; Tanaka, M.; Yoshida, H.; et al. Elevation of fatty acid-binding protein 4 is predisposed by family history of hypertension and contributes to blood pressure elevation. Am. J. Hypertens. 2012, 25, 1124–1130. [Google Scholar] [CrossRef] [Green Version]
- Tso, A.W.; Xu, A.; Sham, P.C.; Wat, N.M.; Wang, Y.; Fong, C.H.; Cheung, B.M.; Janus, E.D.; Lam, K.S. Serum adipocyte fatty acid binding protein as a new biomarker predicting the development of type 2 diabetes: A 10-year prospective study in a Chinese cohort. Diabetes Care 2007, 30, 2667–2672. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.S.; Wu, Y.W.; Huang, J.Y.; Hsu, P.Y.; Chen, M.F. Evaluation of circulating adipokines and abdominal obesity as predictors of significant myocardial ischemia using gated single-photon emission computed tomography. PLoS ONE 2014, 9, e97710. [Google Scholar] [CrossRef]
- Xu, A.; Tso, A.W.; Cheung, B.M.; Wang, Y.; Wat, N.M.; Fong, C.H.; Yeung, D.C.; Janus, E.D.; Sham, P.C.; Lam, K.S. Circulating adipocyte-fatty acid binding protein levels predict the development of the metabolic syndrome: A 5-year prospective study. Circulation 2007, 115, 1537–1543. [Google Scholar] [CrossRef]
- Cabré, A.; Babio, N.; Lázaro, I.; Bulló, M.; Garcia-Arellano, A.; Masana, L.; Salas-Salvadó, J. FABP4 predicts atherogenic dyslipidemia development. The PREDIMED study. Atherosclerosis 2012, 222, 229–234. [Google Scholar] [CrossRef]
- Yeung, D.C.Y.; Xu, A.; Cheung, C.W.S.; Wat, N.M.S.; Yau, M.H.; Fong, C.H.Y.; Chau, M.T.; Lam, K.S.L. Serum adipocyte fatty acid-binding protein levels were independently associated with carotid atherosclerosis. Arter. Thromb. Vasc. Biol. 2007, 27, 1796–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, T.; Onoue, G.; Hirohata, A.; Hirohata, S.; Usui, S.; Hina, K.; Kawamura, H.; Doi, M.; Kusano, K.F.; Kusachi, S.; et al. Serum adipocyte fatty acid-binding protein is independently associated with coronary atherosclerotic burden measured by intravascular ultrasound. Atherosclerosis 2010, 211, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Gu, W.; Zhang, Y.; Yan, Q.; Dai, M.; Shi, J.; Zhai, Y.; Wang, W.; Li, X.; Ning, G. Different association of circulating levels of adipocyte and epidermal fatty acid-binding proteins with metabolic syndrome and coronary atherosclerosis in Chinese adults. Atherosclerosis 2011, 217, 194–200. [Google Scholar] [CrossRef]
- Lee, K.; Santibanez-Koref, M.; Polvikoski, T.; Birchall, D.; Mendelow, A.D.; Keavney, B. Increased expression of fatty acid binding protein 4 and leptin in resident macrophages characterises atherosclerotic plaque rupture. Atherosclerosis 2013, 226, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.L.; Wu, Y.W.; Wu, C.C.; Lin, L.; Wu, Y.C.; Hsu, P.Y.; Jong, Y.S.; Yang, W.S. Association between serum adipocyte fatty-acid binding protein concentrations, left ventricular function and myocardial perfusion abnormalities in patients with coronary artery disease. Cardiovasc. Diabetol. 2013, 12, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotamisligil, G.S.; Johnson, R.S.; Distel, R.J.; Ellis, R.; Papaioannou, V.E.; Spiegelman, B.M. Uncoupling of obesity from insulin resistance through a targeted mutation in aP2, the adipocyte fatty acid binding protein. Science 1996, 274, 1377–1379. [Google Scholar] [CrossRef] [PubMed]
- Makowski, L.; Boord, J.B.; Maeda, K.; Babaev, V.R.; Uysal, K.T.; Morgan, M.A.; Parker, R.A.; Suttles, J.; Fazio, S.; Hotamisligil, G.S.; et al. Lack of macrophage fatty-acid-binding protein aP2 protects mice deficient in apolipoprotein E against atherosclerosis. Nat. Med. 2001, 7, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Boord, J.B.; Maeda, K.; Makowski, L.; Babaev, V.R.; Fazio, S.; Linton, M.F.; Hotamisligil, G.S. Combined adipocyte-macrophage fatty acid-binding protein deficiency improves metabolism, atherosclerosis, and survival in apolipoprotein E-deficient mice. Circulation 2004, 110, 1492–1498. [Google Scholar] [CrossRef] [Green Version]
- Furuhashi, M.; Fucho, R.; Görgün, C.Z.; Tuncman, G.; Cao, H.; Hotamisligil, G.S. Adipocyte/macrophage fatty acid-binding proteins contribute to metabolic deterioration through actions in both macrophages and adipocytes in mice. J. Clin. Investig. 2008, 118, 2640–2650. [Google Scholar] [CrossRef] [Green Version]
- Aragonès, G.; Saavedra, P.; Heras, M.; Cabré, A.; Girona, J.; Masana, L. Fatty acid-binding protein 4 impairs the insulin-dependent nitric oxide pathway in vascular endothelial cells. Cardiovasc. Diabetol. 2012, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- Fuseya, T.; Furuhashi, M.; Matsumoto, M.; Watanabe, Y.; Hoshina, K.; Mita, T.; Ishimura, S.; Tanaka, M.; Miura, T. Ectopic fatty acid-binding protein 4 expression in the vascular endothelium is involved in neointima formation after vascular injury. J. Am. Heart Assoc. 2017, 6, 9. [Google Scholar] [CrossRef]
- Elmasri, H.; Karaaslan, C.; Teper, Y.; Ghelfi, E.; Weng, M.; Ince, T.A.; Kozakewich, H.; Bischoff, J.; Cataltepe, S. Fatty acid binding protein 4 is a target of VEGF and a regulator of cell proliferation in endothelial cells. FASEB J. 2009, 23, 3865–3873. [Google Scholar] [CrossRef] [Green Version]
- Harjes, U.; Bridges, E.; McIntyre, A.; Fielding, B.A.; Harris, A.L. Fatty acid-binding protein 4, a point of convergence for angiogenic and metabolic signaling pathways in endothelial cells. J. Biol. Chem. 2014, 289, 23168–23176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmasri, H.; Ghelfi, E.; Yu, C.W.; Traphagen, S.; Cernadas, M.; Cao, H.; Shi, G.P.; Plutzky, J.; Sahin, M.; Hotamisligil, G.; et al. Endothelial cell-fatty acid binding protein 4 promotes angiogenesis: Role of stem cell factor/c-kit pathway. Angiogenesis 2012, 15, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girona, J.; Rosales, R.; Plana, N.; Saavedra, P.; Masana, L.; Vallvé, J.C. FABP4 induces vascular smooth muscle cell proliferation and migration through a MAPK-dependent pathway. PLoS ONE 2013, 8, e81914. [Google Scholar] [CrossRef] [PubMed]
- Karpisek, M.; Stejskal, D.; Kotolova, H.; Kollar, P.; Janoutova, G.; Ochmanova, R.; Cizek, L.; Horakova, D.; Yahia, R.B.; Lichnovska, R.; et al. Treatment with atorvastatin reduces serum adipocyte-fatty acid binding protein value in patients with hyperlipidaemia. Eur. J. Clin. Investig. 2007, 37, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.W.; Kao, H.L.; Huang, C.L.; Chen, M.F.; Lin, L.Y.; Wang, Y.C.; Lin, Y.H.; Lin, H.J.; Tzen, K.Y.; Yen, R.F.; et al. The effects of 3-month atorvastatin therapy on arterial inflammation, calcification, abdominal adipose tissue and circulating biomarkers. Eur. J. Nucl. Med. Mol. Imaging 2012, 39, 399–407. [Google Scholar] [CrossRef]
- Fu, Y.; Luo, N.; Lopes-Virella, M.F.; Garvey, W.T. The adipocyte lipid binding protein (ALBP/aP2) gene facilitates foam cell formation in human THP-1 macrophages. Atherosclerosis 2002, 165, 259–269. [Google Scholar] [CrossRef]
- Fu, Y.; Luo, L.; Luo, N.; Garvey, W.T. Lipid metabolism mediated by adipocyte lipid binding protein (ALBP/aP2) gene expression in human THP-1 macrophages. Atherosclerosis 2006, 188, 102–111. [Google Scholar] [CrossRef]
- Aragonès, G.; Ferré, R.; Lázaro, I.; Cabré, A.; Plana, N.; Merino, J.; Heras, M.; Girona, J.; Masana, L. Fatty acid-binding protein 4 is associated with endothelial dysfunction in patients with type 2 diabetes. Atherosclerosis 2010, 213, 329–331. [Google Scholar] [CrossRef]
- Lee, M.Y.; Li, H.; Xiao, Y.; Zhou, Z.; Xu, A.; Vanhoutte, P.M. Chronic administration of BMS309403 improves endothelial function in apolipoprotein E-deficient mice and in cultured human endothelial cells. Br. J. Pharmacol. 2011, 162, 1564–1576. [Google Scholar] [CrossRef] [Green Version]
- Makowski, L.; Brittingham, K.C.; Reynolds, J.M.; Suttles, J.; Hotamisligil, G.S. The fatty acid-binding protein, aP2, coordinates macrophage cholesterol trafficking and inflammatory activity. Macrophage expression of aP2 impacts peroxisome proliferator-activated receptor gamma and IkappaB kinase activities. J. Biol. Chem. 2005, 280, 12888–12895. [Google Scholar] [CrossRef] [Green Version]
- Hui, X.; Li, H.; Zhou, Z.; Lam, K.S.; Xiao, Y.; Wu, D.; Ding, K.; Wang, Y.; Vanhoutte, P.M.; Xu, A. Adipocyte fatty acid-binding protein modulates inflammatory responses in macrophages through a positive feedback loop involving c-Jun NH2-terminal kinases and activator protein-1. J. Biol. Chem. 2010, 285, 10273–10280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikorski, K.; Czerwoniec, A.; Bujnicki, J.M.; Wesoly, J.; Bluyssen, H.A. STAT1 as a novel therapeutical target in pro-atherogenic signal integration of IFNγ, TLR4 and IL-6 in vascular disease. Cytokine Growth Factor Rev. 2011, 22, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.; Döring, Y.; Noels, H. Potential cell-specific functions of CXCR4 in atherosclerosis. Hamostaseologie 2016, 36, 97–102. [Google Scholar] [PubMed] [Green Version]
- Perrotta, P.; Van der Veken, B.; Van Der Veken, P.; Pintelon, I.; Roosens, L.; Adriaenssens, E.; Timmerman, V.; Guns, P.J.; De Meyer, G.R.Y.; Martinet, W. Partial Inhibition of Glycolysis Reduces Atherogenesis Independent of Intraplaque Neovascularization in Mice. Arter. Thromb. Vasc. Biol. 2020, 40, 1168–1181. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAD | Controls | p | |
|---|---|---|---|
| n = 22 | n = 40 | ||
| Age (year) | 66 ± 10 | 34 ± 7 | <0.0001 |
| Male gender (n, %) | 86% | 40% | 0.0022 |
| Waist (cm) | 85 ± 10 | 80 ± 13 | 0.15 |
| Hip (cm) | 85 ± 9 | 91 ± 10 | 0.0367 |
| WHR | 1.00 ± 0.09 | 0.89 ± 0.11 | 0.0002 |
| BH (cm) | 164 ± 10 | 167 ± 9 | 0.22 |
| BW (kg) | 69 ± 12 | 68 ± 14 | 0.81 |
| BMI | 25.5 ± 3.7 | 24.1 ± 3.6 | 0.16 |
| Systolic BP (mmHg) | 140 ± 17 | 126 ± 15 | 0.0013 |
| Diastolic BP (mmHg) | 78 ± 12 | 75 ± 11 | 0.32 |
| HR (bpm) | 76 ± 13 | 72 ± 8 | 0.09 |
| Family History | |||
| MI/CVA/SD (%) | 23% | 18% | 0.63 |
| DM (%) | 18% | 15% | 0.75 |
| Clinical History | |||
| Hypertension (%) | 82% | 23% | <0.0001 |
| Type 2 DM (%) | 41% | 0% | <0.0001 |
| CVA (ischemic/TIA) (%) | 5% | 0% | 0.18 |
| PAOD (%) | 5% | 0% | 0.18 |
| Smoking (%) | 41% | 13% | 0.0099 |
| Alcohol consumption (%) | 27% | 10% | 0.08 |
| Physical Activity (≥30 min) (%) | 9% | 48% | 0.0018 |
| Medication | <0.0001 | ||
| Beta-blockers (%) | 64% | 5% | |
| ACEi/ARB (%) | 36% | 3% | |
| CCB (%) | 27% | 3% | |
| Statin (%) | 64% | 3% | |
| Lab data | |||
| T-CHO (mg/dl) | 144 ± 28 | 179 ± 29 | <0.0001 |
| HDL-C (mg/dl) | 47 ± 18 | 64 ± 18 | 0.0036 |
| LDL-C (mg/dl) | 85 ± 22 | 105 ± 31 | 0.002 |
| Triglyceride (mg/dl) | 118 ± 65 | 100 ± 55 | 0.27 |
| Fasting glucose (mg/dl) | 114 ± 41 | 92 ± 16 | 0.0069 |
| ALT (IU/L) | 29 ± 24 | 22 ± 14 | 0.20 |
| Cre (mg/dl) | 1.6 ± 2.1 | 0.8 ± 0.2 | 0.0256 |
| UA (mg/dl) | 5.6 ± 1.1 | 6.2 ± 1.6 | 0.14 |
| Mean Adhesion Cell No. (n) | 22.6 ± 21.2 | 38 ± 29 | 0.0306 |
| FABP4 concentration | |||
| Plasma (pg/mL) | 5240 ± 1019 | 5052 ± 1037 | 0.5 |
| Supernatant (pg/mL) | 3365 ± 1500 | 2061 ± 829 | 0.0001 |
| oxLDL (U/L) | 50.2 ± 16.8 | 52.7 ± 14.3 | 0.54 |
| Univariate | Multivariate | |||
|---|---|---|---|---|
| r | p | ß | p | |
| Age | 0.5167 | <0.0001 | ||
| WHR | 0.5853 | 0.0002 | ||
| HTN | 0.4642 | <0.0001 | 727.6 | 0.008 |
| DM | 0.3942 | 0.0002 | ||
| Physical Activity | −0.514 | <0.0001 | ||
| HDL-C | −0.3321 | 0.036 | ||
| Cre | 0.4839 | <0.0001 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.-W.; Chang, T.-T.; Chang, C.-C.; Chen, J.-W. Fatty-Acid-Binding Protein 4 as a Novel Contributor to Mononuclear Cell Activation and Endothelial Cell Dysfunction in Atherosclerosis. Int. J. Mol. Sci. 2020, 21, 9245. https://doi.org/10.3390/ijms21239245
Wu Y-W, Chang T-T, Chang C-C, Chen J-W. Fatty-Acid-Binding Protein 4 as a Novel Contributor to Mononuclear Cell Activation and Endothelial Cell Dysfunction in Atherosclerosis. International Journal of Molecular Sciences. 2020; 21(23):9245. https://doi.org/10.3390/ijms21239245
Chicago/Turabian StyleWu, Yen-Wen, Ting-Ting Chang, Chia-Chi Chang, and Jaw-Wen Chen. 2020. "Fatty-Acid-Binding Protein 4 as a Novel Contributor to Mononuclear Cell Activation and Endothelial Cell Dysfunction in Atherosclerosis" International Journal of Molecular Sciences 21, no. 23: 9245. https://doi.org/10.3390/ijms21239245