Dimethyl Fumarate Attenuates Lung Inflammation and Oxidative Stress Induced by Chronic Exposure to Diesel Exhaust Particles in Mice

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Results
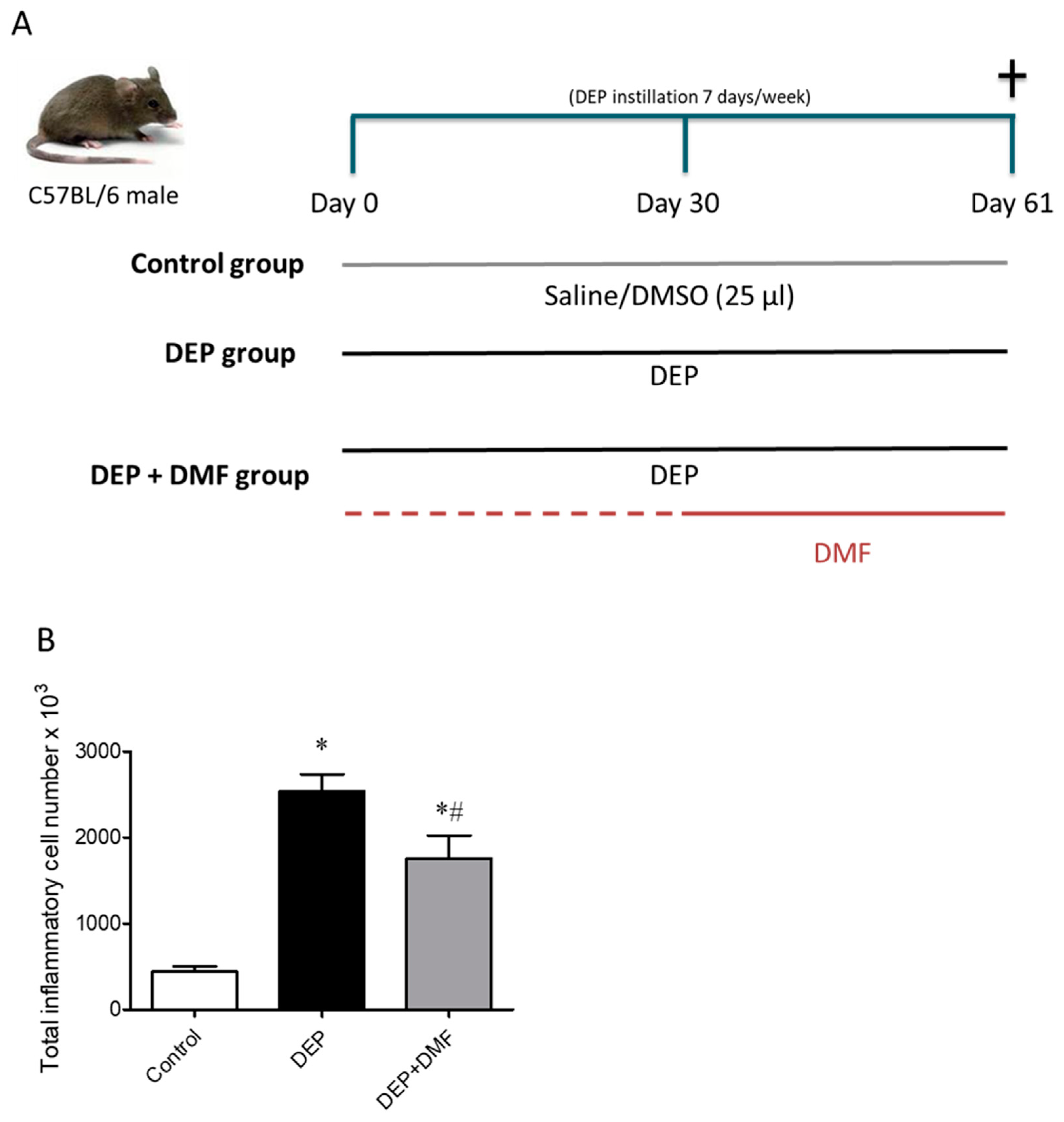
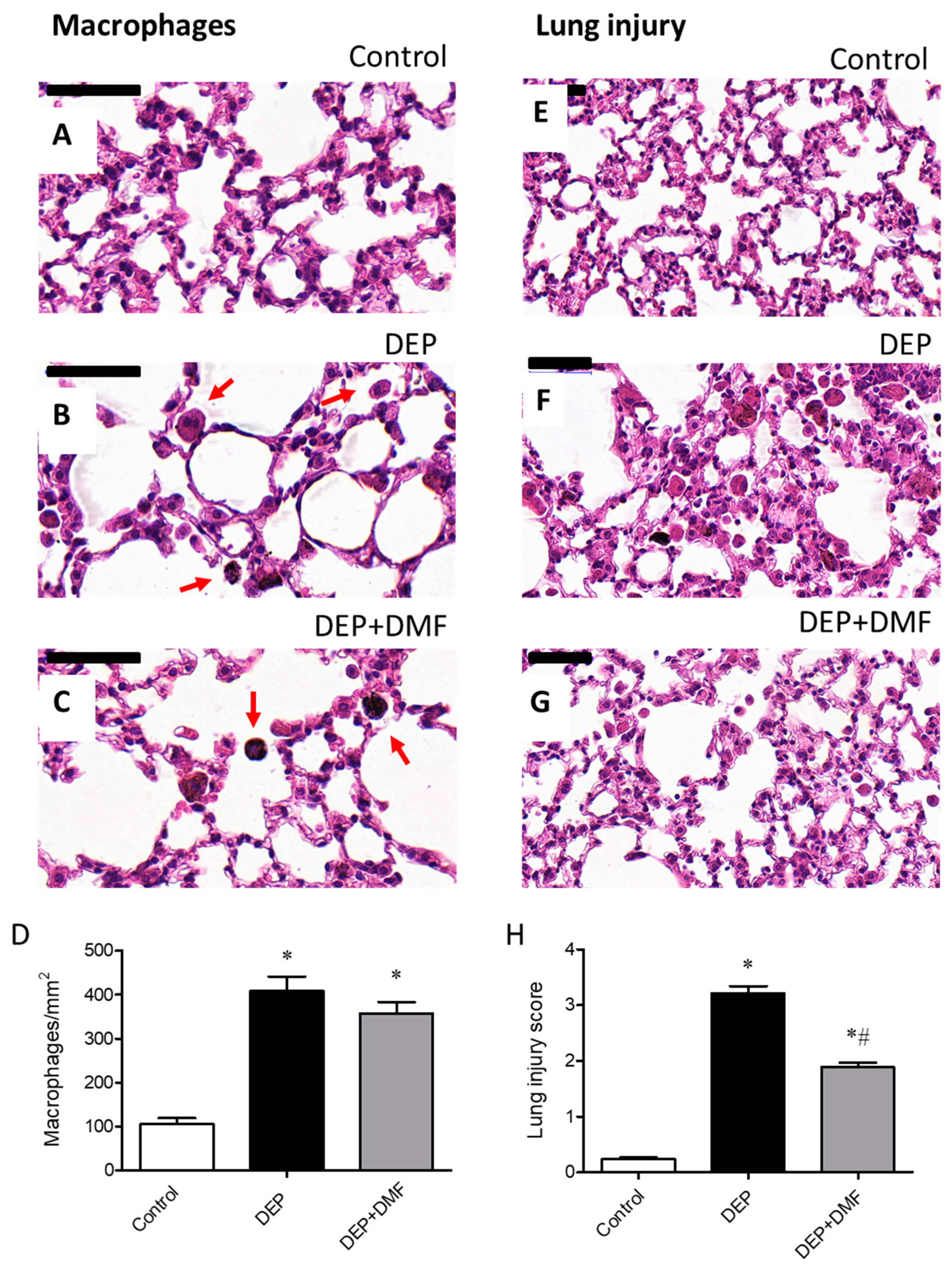
2.1. DMF Reduces Total Inflammatory Cell Number in BAL and Lung Injury Caused by DEP Exposure
2.2. DMF Reduces Oxidative and Nitrosative Stress after DEP Instillation
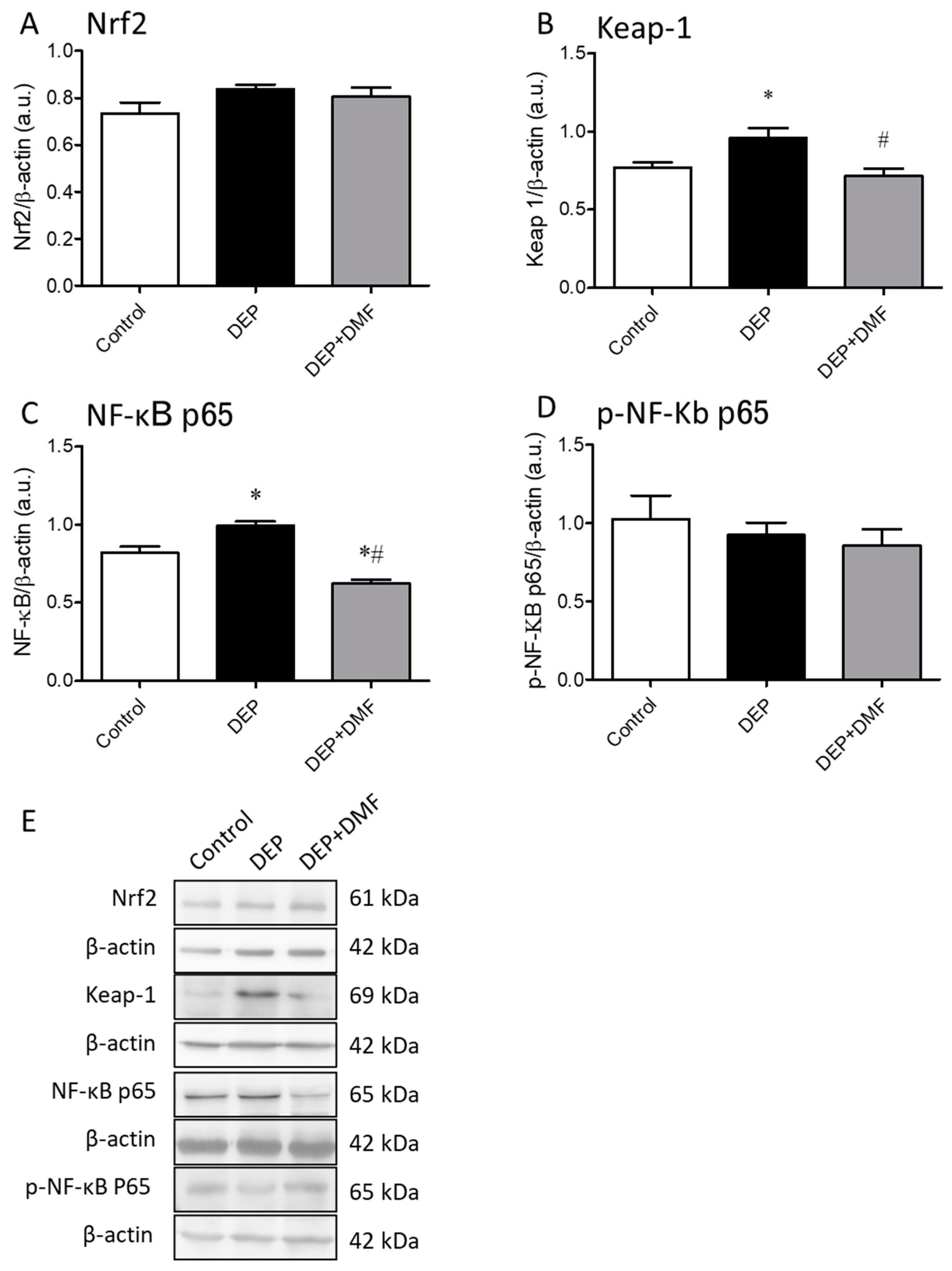
2.3. DMF Treatment Decreases DEP-Induced Keap-1 and NF-κB Expression in Mouse Lungs
2.4. DMF Attenuates the Effects of Chronic DEP Exposure on the Expression of Nitrotyrosine and Redox System-Associated Proteins in Mouse Lungs
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Diesel Exhaust Particles (DEPs)
4.3. Dimethyl Fumarate (DMF)
4.4. Experimental Design
4.5. Bronchoalveolar Lavage (BAL) and Lung Homogenates
4.6. Reactive Oxygen Species (ROS) and Probe Analysis for Intracellular Total ROS, Peroxynitrite (OONO) and Nitric Oxide (NO)
4.7. Tissue Preparation and Microscopic Analyses
4.8. Immunoblotting
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johannson, K.A.; Balmes, J.R.; Collard, H.R. Air pollution exposure: A novel environmental risk factor for interstitial lung disease? Chest 2015, 147, 1161–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prüss-Üstün, A.; Wolf, J.; Corvalán, C.F.; Bos, R.; Neira, M.P. Preventing Disease Through Healthy Environments: A Global Assessment of the Burden of Disease from Environmental Risks; World Health Organization: Geneva, Switzerland, 2016; p. 176. [Google Scholar]
- Shin, S.; Bai, L.; Burnett, R.T.; Kwong, J.C.; Hystad, P.; van Donkelaar, A.; Lavigne, E.; Weichenthal, S.; Copes, R.; Martin, R.V.; et al. Air Pollution as a Risk Factor for Incident COPD and Asthma: 15-Year Population-Based Cohort Study. Am. J. Respir. Crit. Care Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Annesi-Maesano, I. Air Pollution and Chronic Obstructive Pulmonary Disease Exacerbations: When Prevention Becomes Feasible. Am. J. Respir. Crit. Care Med. 2019, 199, 547–548. [Google Scholar] [CrossRef] [PubMed]
- Steiner, S.; Bisig, C.; Petri-Fink, A.; Rothen-Rutishauser, B. Diesel exhaust: Current knowledge of adverse effects and underlying cellular mechanisms. Arch. Toxicol. 2016, 90, 1541–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EPA. Quantitative Health Risk Assessment for Particulate Matter. In Particulate Matter (PM) Standards—Documents from Review Completed in 2012—Risk and Exposure Assessments, 1st ed.; McCarthy, G., Ed.; United States Environmental Protection Agency: Washington, DC, USA, 2010; pp. 1–596. [Google Scholar]
- Choi, J.; Oh, J.Y.; Lee, Y.S.; Min, K.H.; Hur, G.Y.; Lee, S.Y.; Kang, K.H.; Shim, J.J. Harmful impact of air pollution on severe acute exacerbation of chronic obstructive pulmonary disease: Particulate matter is hazardous. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 1053–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pothirat, C.; Chaiwong, W.; Liwsrisakun, C.; Bumroongkit, C.; Deesomchok, A.; Theerakittikul, T.; Limsukon, A.; Tajarernmuang, P.; Phetsuk, N. Acute effects of air pollutants on daily mortality and hospitalizations due to cardiovascular and respiratory diseases. J. Thorac. Dis. 2019, 11, 3070–3083. [Google Scholar] [CrossRef]
- GOLD. Global Initiative for Chronic Obstructive Lung Disease. In Global Strategy for the Diagnosis, Management and Prevention of COPD; GOLD: Fontana, WI, USA, 2020; Available online: https://goldcopd.org (accessed on 9 October 2020).
- Reddel, H.K.; FitzGerald, J.M.; Bateman, E.D.; Bacharier, L.B.; Becker, A.; Brusselle, G.; Buhl, R.; Cruz, A.A.; Fleming, L.; Inoue, H.; et al. GINA 2019: A fundamental change in asthma management: Treatment of asthma with short-acting bronchodilators alone is no longer recommended for adults and adolescents. Eur. Respir. J. 2019, 53. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Air Quality Guidelines for Particulate Matter, Ozone, Nitrogen Dioxide and Sulfur Dioxide. Global Update 2005; WHO: Geneva, Switzerland, 2006; pp. 1–22. [Google Scholar]
- Larcombe, A.N.; Phan, J.A.; Kicic, A.; Perks, K.L.; Mead-Hunter, R.; Mullins, B.J. Route of exposure alters inflammation and lung function responses to diesel exhaust. Inhal. Toxicol. 2014, 26, 409–418. [Google Scholar] [CrossRef]
- Nemmar, A.; Subramaniyan, D.; Zia, S.; Yasin, J.; Ali, B.H. Airway resistance, inflammation and oxidative stress following exposure to diesel exhaust particle in angiotensin II-induced hypertension in mice. Toxicology 2012, 292, 162–168. [Google Scholar] [CrossRef]
- Hart, J.E.; Eisen, E.A.; Laden, F. Occupational diesel exhaust exposure as a risk factor for chronic obstructive pulmonary disease. Curr. Opin. Pulm. Med. 2012, 18, 151–154. [Google Scholar] [CrossRef]
- Xu, Y.; Barregard, L.; Nielsen, J.; Gudmundsson, A.; Wierzbicka, A.; Axmon, A.; Jonsson, B.A.; Karedal, M.; Albin, M. Effects of diesel exposure on lung function and inflammation biomarkers from airway and peripheral blood of healthy volunteers in a chamber study. Part. Fibre Toxicol. 2013, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Li, Q.; Liu, H.; Lin, L.; Han, W.; Hao, W. Role of C/EBPalpha hypermethylation in diesel engine exhaust exposure-induced lung inflammation. Ecotoxicol. Environ. Saf. 2019, 183, 109500. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, K.; Azuma, A.; Kudoh, S.; Desaki, M.; Takizawa, H.; Sugawara, I. Inhalation of diesel exhaust for three months affects major cytokine expression and induces bronchus-associated lymphoid tissue formation in murine lungs. Exp. Lung Res. 2003, 29, 607–622. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, K.; Brito, J.M.; Toledo, A.C.; Nakagawa, N.K.; Piccin, V.S.; Junqueira, M.S.; Negri, E.M.; Carvalho, A.L.; Oliveira, A.P.; Lima, W.T.; et al. Subchronic effects of nasally instilled diesel exhaust particulates on the nasal and airway epithelia in mice. Inhal. Toxicol. 2010, 22, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Yuvaraju, P.; Hamadi, N.; Ali, B.H. In Vivo Protective Effects of Nootkatone against Particles-Induced Lung Injury Caused by Diesel Exhaust Is Mediated via the NF-kappaB Pathway. Nutrients 2018, 10, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.I.; Song, M.K.; Kim, S.H.; Park, C.Y.; Lee, K. TF-343 Alleviates Diesel Exhaust Particulate-Induced Lung Inflammation via Modulation of Nuclear Factor-kappaB Signaling. J. Immunol. Res. 2019, 2019, 8315845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.P.; Zhang, X.; Duan, H.W.; Meng, T.; Niu, Y.; Huang, C.F.; Gao, W.M.; Yu, S.F.; Zheng, Y.X. Long-term exposure to diesel engine exhaust induced lung function decline in a cross sectional study. Ind. Health 2017, 55, 13–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, G.; Bin, P.; Meng, T.; Niu, Y.; Yang, M.; Zhang, L.; Duan, H.; Yu, T.; Dai, Y.; et al. Time-course effects of antioxidants and phase II enzymes on diesel exhaust particles-induced oxidative damage in the mouse lung. Toxicol. Appl. Pharmacol. 2019, 366, 25–34. [Google Scholar] [CrossRef]
- Cattani-Cavalieri, I.; Valenca, S.S.; Lanzetti, M.; Carvalho, G.M.C.; Zin, W.A.; Monte-Alto-Costa, A.; Porto, L.C.; Romana-Souza, B. Acute Exposure to Diesel-Biodiesel Particulate Matter Promotes Murine Lung Oxidative Stress by Nrf2/HO-1 and Inflammation Through the NF-kB/TNF-alpha Pathways. Inflammation 2019, 42, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Saidu, N.E.B.; Kavian, N.; Leroy, K.; Jacob, C.; Nicco, C.; Batteux, F.; Alexandre, J. Dimethyl fumarate, a two-edged drug: Current status and future directions. Med. Res. Rev. 2019, 39, 1923–1952. [Google Scholar] [CrossRef] [PubMed]
- Seidel, P.; Merfort, I.; Hughes, J.M.; Oliver, B.G.; Tamm, M.; Roth, M. Dimethylfumarate inhibits NF-{kappa}B function at multiple levels to limit airway smooth muscle cell cytokine secretion. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L326–L339. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, R.S.; Abdel-Rahman, N. Dimethyl fumarate ameliorates acetaminophen-induced hepatic injury in mice dependent of Nrf-2/HO-1 pathway. Life Sci. 2019, 217, 251–260. [Google Scholar] [CrossRef]
- Belcher, J.D.; Chen, C.; Nguyen, J.; Zhang, P.; Abdulla, F.; Nguyen, P.; Killeen, T.; Xu, P.; O’Sullivan, G.; Nath, K.A.; et al. Control of Oxidative Stress and Inflammation in Sickle Cell Disease with the Nrf2 Activator Dimethyl Fumarate. Antioxid. Redox Signal. 2017, 26, 748–762. [Google Scholar] [CrossRef] [PubMed]
- Campolo, M.; Casili, G.; Biundo, F.; Crupi, R.; Cordaro, M.; Cuzzocrea, S.; Esposito, E. The Neuroprotective Effect of Dimethyl Fumarate in an MPTP-Mouse Model of Parkinson’s Disease: Involvement of Reactive Oxygen Species/Nuclear Factor-kappaB/Nuclear Transcription Factor Related to NF-E2. Antioxid. Redox Signal. 2017, 27, 453–471. [Google Scholar] [CrossRef] [Green Version]
- Casili, G.; Campolo, M.; Paterniti, I.; Lanza, M.; Filippone, A.; Cuzzocrea, S.; Esposito, E. Dimethyl Fumarate Attenuates Neuroinflammation and Neurobehavioral Deficits Induced by Experimental Traumatic Brain Injury. J. Neurotrauma 2018, 35, 1437–1451. [Google Scholar] [CrossRef]
- Motterlini, R.; Nikam, A.; Manin, S.; Ollivier, A.; Wilson, J.L.; Djouadi, S.; Muchova, L.; Martens, T.; Rivard, M.; Foresti, R. HYCO-3, a dual CO-releaser/Nrf2 activator, reduces tissue inflammation in mice challenged with lipopolysaccharide. Redox Biol. 2019, 20, 334–348. [Google Scholar] [CrossRef]
- Mrowietz, U.; Asadullah, K. Dimethylfumarate for psoriasis: More than a dietary curiosity. Trends Mol. Med. 2005, 11, 43–48. [Google Scholar] [CrossRef]
- Linker, R.A.; Lee, D.H.; Ryan, S.; van Dam, A.M.; Conrad, R.; Bista, P.; Zeng, W.; Hronowsky, X.; Buko, A.; Chollate, S.; et al. Fumaric acid esters exert neuroprotective effects in neuroinflammation via activation of the Nrf2 antioxidant pathway. Brain 2011, 134, 678–692. [Google Scholar] [CrossRef] [Green Version]
- Scannevin, R.H.; Chollate, S.; Jung, M.Y.; Shackett, M.; Patel, H.; Bista, P.; Zeng, W.; Ryan, S.; Yamamoto, M.; Lukashev, M.; et al. Fumarates promote cytoprotection of central nervous system cells against oxidative stress via the nuclear factor (erythroid-derived 2)-like 2 pathway. J. Pharmacol. Exp. Ther. 2012, 341, 274–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzoli-Rocha, F.; Magalhaes, C.B.; Malm, O.; Saldiva, P.H.; Zin, W.A.; Faffe, D.S. Comparative respiratory toxicity of particles produced by traffic and sugar cane burning. Environ. Res. 2008, 108, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Mukae, H.; Hogg, J.C.; English, D.; Vincent, R.; van Eeden, S.F. Phagocytosis of particulate air pollutants by human alveolar macrophages stimulates the bone marrow. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L924–L931. [Google Scholar] [CrossRef]
- Santana, F.P.R.; Pinheiro, N.M.; Bittencourt-Mernak, M.I.; Perini, A.; Yoshizaki, K.; Macchione, M.; Saldiva, P.H.N.; Martins, M.A.; Tiberio, I.; Prado, M.A.M.; et al. Vesicular acetylcholine transport deficiency potentiates some inflammatory responses induced by diesel exhaust particles. Ecotoxicol. Environ. Saf. 2019, 167, 494–504. [Google Scholar] [CrossRef]
- Villeneuve, N.F.; Lau, A.; Zhang, D.D. Regulation of the Nrf2-Keap1 antioxidant response by the ubiquitin proteasome system: An insight into cullin-ring ubiquitin ligases. Antioxid. Redox Signal. 2010, 13, 1699–1712. [Google Scholar] [CrossRef] [Green Version]
- Yoshizaki, K.; Brito, J.M.; Moriya, H.T.; Toledo, A.C.; Ferzilan, S.; Ligeiro de Oliveira, A.P.; Machado, I.D.; Farsky, S.H.; Silva, L.F.; Martins, M.A.; et al. Chronic exposure of diesel exhaust particles induces alveolar enlargement in mice. Respir. Res. 2015, 16, 18. [Google Scholar] [CrossRef] [Green Version]
- Budinger, G.R.; McKell, J.L.; Urich, D.; Foiles, N.; Weiss, I.; Chiarella, S.E.; Gonzalez, A.; Soberanes, S.; Ghio, A.J.; Nigdelioglu, R.; et al. Particulate matter-induced lung inflammation increases systemic levels of PAI-1 and activates coagulation through distinct mechanisms. PLoS ONE 2011, 6, e18525. [Google Scholar] [CrossRef] [Green Version]
- Soodaeva, S.; Kubysheva, N.; Klimanov, I.; Nikitina, L.; Batyrshin, I. Features of Oxidative and Nitrosative Metabolism in Lung Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 1689861. [Google Scholar] [CrossRef] [Green Version]
- Pardo, M.; Porat, Z.; Rudich, A.; Schauer, J.J.; Rudich, Y. Repeated exposures to roadside particulate matter extracts suppresses pulmonary defense mechanisms, resulting in lipid and protein oxidative damage. Environ. Pollut. 2016, 210, 227–237. [Google Scholar] [CrossRef]
- Kaminsky, D.A.; Mitchell, J.; Carroll, N.; James, A.; Soultanakis, R.; Janssen, Y. Nitrotyrosine formation in the airways and lung parenchyma of patients with asthma. J. Allergy Clin. Immunol. 1999, 104, 747–754. [Google Scholar] [CrossRef]
- Ribeiro Junior, G.; de Souza Xavier Costa, N.; Belotti, L.; Dos Santos Alemany, A.A.; Amato-Lourenco, L.F.; da Cunha, P.G.; de Oliveira Duro, S.; Ribeiro, S.P.; Veras, M.M.; Quirino Dos Santos Lopes, F.D.T.; et al. Diesel exhaust exposure intensifies inflammatory and structural changes associated with lung aging in mice. Ecotoxicol. Environ. Saf. 2019, 170, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Dean, J.M.; Lovely, M.E.; Wang, H. Foreign Direct Investment and Pollution Havens: Evaluating the Evidence From China; Office of Economics, US International Trade Commission: Washington, DC, USA, 2004.
- Choi, H.S.; Kim, J.W.; Cha, Y.N.; Kim, C. A quantitative nitroblue tetrazolium assay for determining intracellular superoxide anion production in phagocytic cells. J. Immunoass. Immunochem. 2006, 27, 31–44. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Frevert, C.W.; Kajikawa, O.; Skerrett, S.J.; Goodman, R.B.; Park, D.R.; Martin, T.R. Septic shock and acute lung injury in rabbits with peritonitis: Failure of the neutrophil response to localized infection. Am. J. Respir. Crit. Care Med. 2001, 163, 234–243. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Groups | ||
|---|---|---|---|
| Control | DEP | DEP + DMF | |
| Total ROS (µg formazan/103 cell) | 44.81 ± 11.22 | 771.5 ± 111.1 * | 407.9 ± 60.62 *,# |
| ROS (H2DCFDA) (% to control) | 1.00 ± 0.22 | 3.02 ± 0.59 * | 1.44 ± 0.27 # |
| NO (DAF) (% to control) | 1.00 ± 0.32 | 2.48 ± 0.40 * | 1.21 ± 0.25 # |
| OONO (APF) (% to control) | 1.00 ± 0.22 | 3.00 ± 0.73 * | 1.31 ± 0.26 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cattani-Cavalieri, I.; da Maia Valença, H.; Moraes, J.A.; Brito-Gitirana, L.; Romana-Souza, B.; Schmidt, M.; Valença, S.S. Dimethyl Fumarate Attenuates Lung Inflammation and Oxidative Stress Induced by Chronic Exposure to Diesel Exhaust Particles in Mice. Int. J. Mol. Sci. 2020, 21, 9658. https://doi.org/10.3390/ijms21249658
Cattani-Cavalieri I, da Maia Valença H, Moraes JA, Brito-Gitirana L, Romana-Souza B, Schmidt M, Valença SS. Dimethyl Fumarate Attenuates Lung Inflammation and Oxidative Stress Induced by Chronic Exposure to Diesel Exhaust Particles in Mice. International Journal of Molecular Sciences. 2020; 21(24):9658. https://doi.org/10.3390/ijms21249658
Chicago/Turabian StyleCattani-Cavalieri, Isabella, Helber da Maia Valença, João Alfredo Moraes, Lycia Brito-Gitirana, Bruna Romana-Souza, Martina Schmidt, and Samuel Santos Valença. 2020. "Dimethyl Fumarate Attenuates Lung Inflammation and Oxidative Stress Induced by Chronic Exposure to Diesel Exhaust Particles in Mice" International Journal of Molecular Sciences 21, no. 24: 9658. https://doi.org/10.3390/ijms21249658
APA StyleCattani-Cavalieri, I., da Maia Valença, H., Moraes, J. A., Brito-Gitirana, L., Romana-Souza, B., Schmidt, M., & Valença, S. S. (2020). Dimethyl Fumarate Attenuates Lung Inflammation and Oxidative Stress Induced by Chronic Exposure to Diesel Exhaust Particles in Mice. International Journal of Molecular Sciences, 21(24), 9658. https://doi.org/10.3390/ijms21249658