Systems Biology Analysis Reveals Eight SLC22 Transporter Subgroups, Including OATs, OCTs, and OCTNs

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Analysis of Substrate Specificity and Selectivity Helps Categorize Mono-, Oligo-, and Multi-Specificity of SLC22 Members
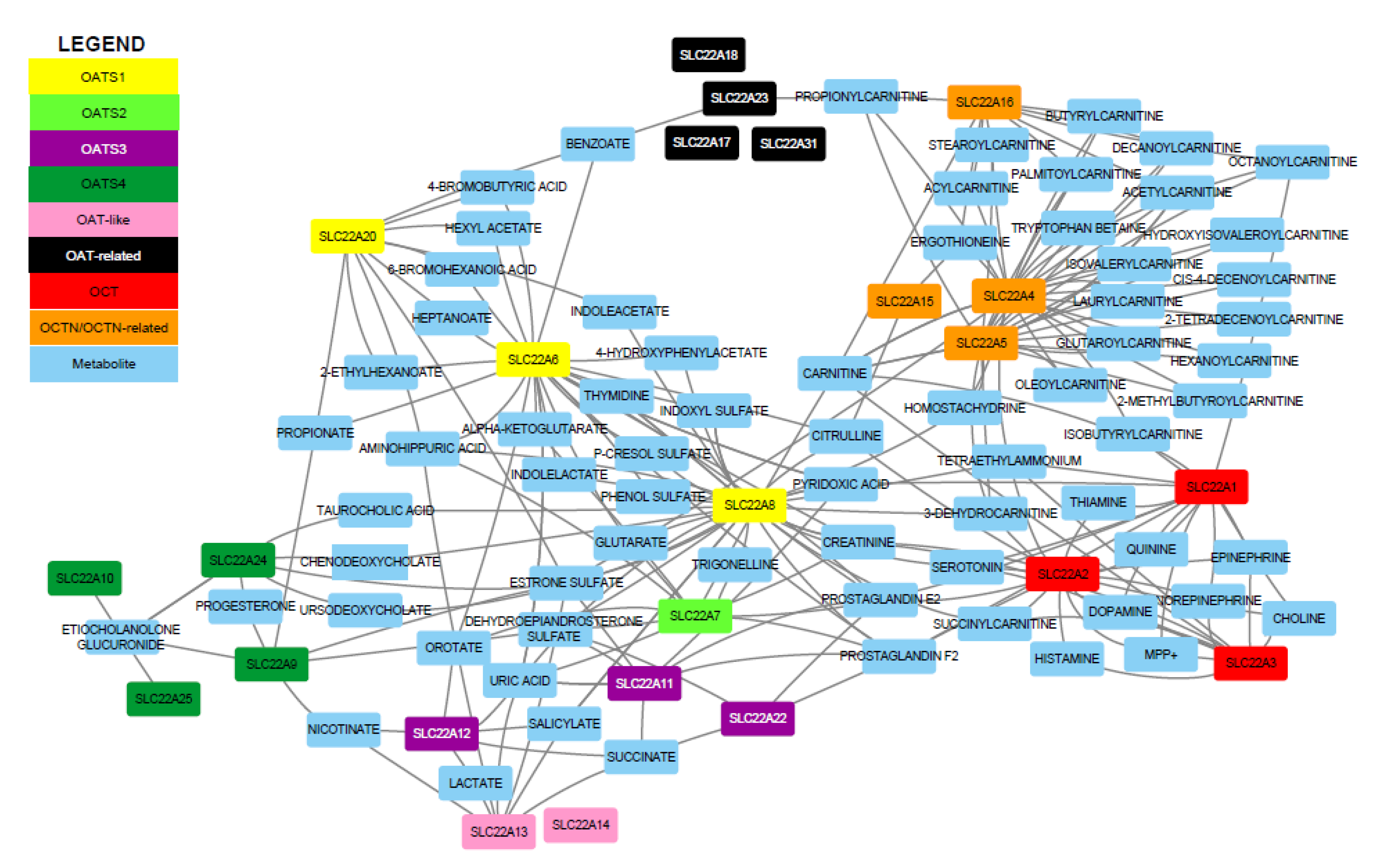
2.2. Construction of Functional Networks from Metabolite-Transporter Interaction Data Support the Eight Subgroups
2.3. OATS1 (SLC22A6, SLC22A8, and SLC22A20) Handles a Wide Variety of Metabolites, Signaling Molecules, Uremic Toxins, and Odorants
2.4. OATS2 (SLC22A7) is a Systemically-Expressed Transporter of Organic Anions and Cyclic Nucleotides
2.5. OATS3 (SLC22A11, SLC22A12, and Slc22a22) Functions to Balance Uric Acid and Prostaglandins
2.6. OATS4 (SLC22A9, SLC22A10, SLC2A24, and SLC22A25) Members are Specifically Associated with Conjugated Sex Hormones
2.7. OAT-Like (SLC22A13 and SLC22A14) has Potentially Physiologically Important Roles
2.8. OAT-Related (SLC22A17, SLC22A18, SLC22A23, and SLC22A31) is Anomalous Amongst SLC22 Members but has Interesting Functional Mechanisms and Disease Associations
2.9. OCT (SLC22A1, SLC22A2, and SLC22A3) Members Are Characteristic Organic Cation Transporters with High Affinities for Monoamine Neurotransmitters and Other Biologically Important Metabolites and Signaling Molecules
2.10. OCTN/OCTN-Related (SLC22A4, SLC22A5, SLC22A15, and SLC22A16) Subgroup Consists of Prototypical Carnitine and Ergothioneine Transporters
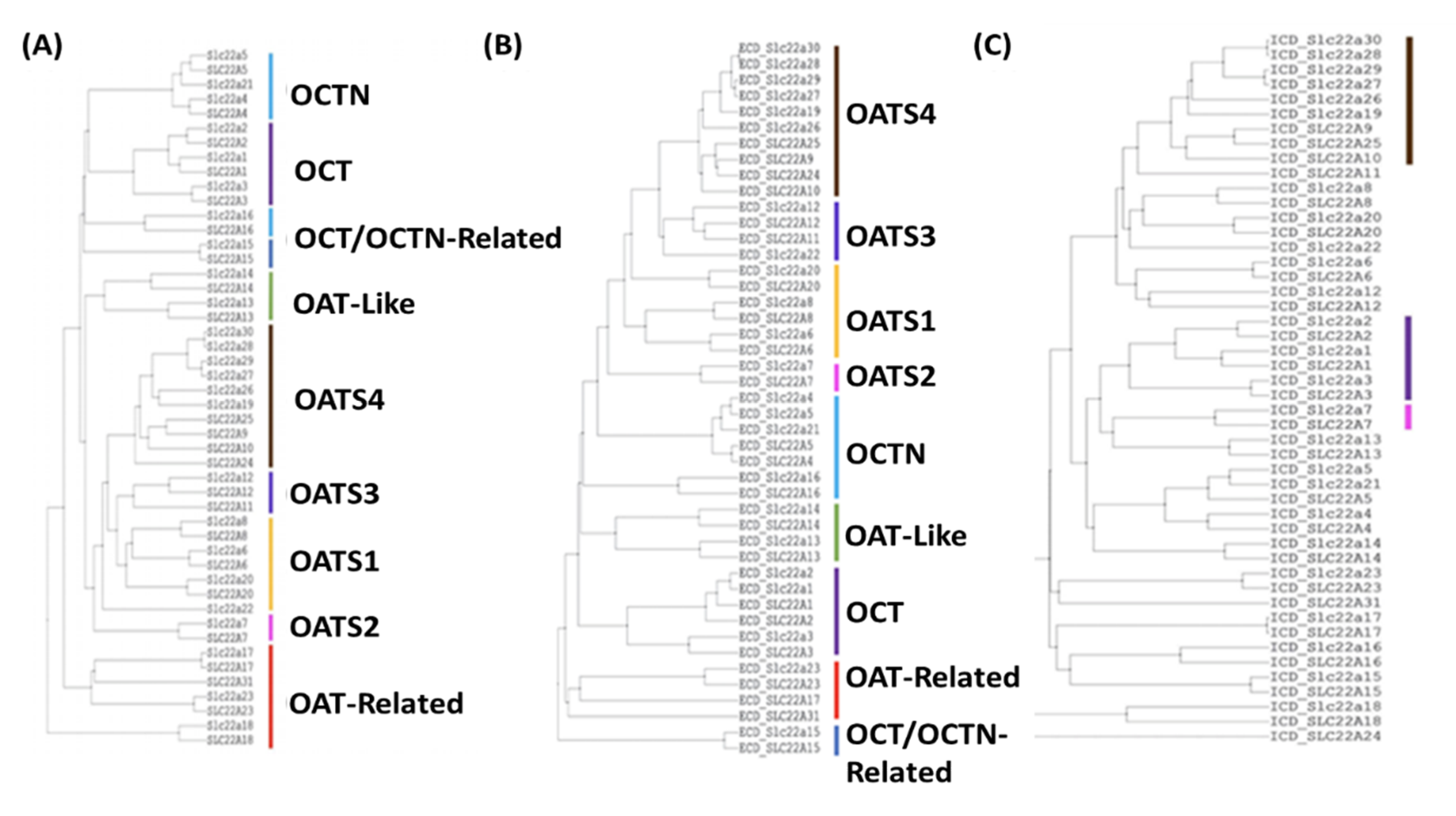
2.11. Multiple Sequence Alignment Further Supports the Classification of Subgroups
2.12. Analysis of Genomic Localization Highlights Evolutionary Relatedness of Subgroup Members and Suggests Basis of Coregulation
2.13. Analysis of OAT Subgroup Specific Motifs Highlight Patterns Potentially Involved in Specificity
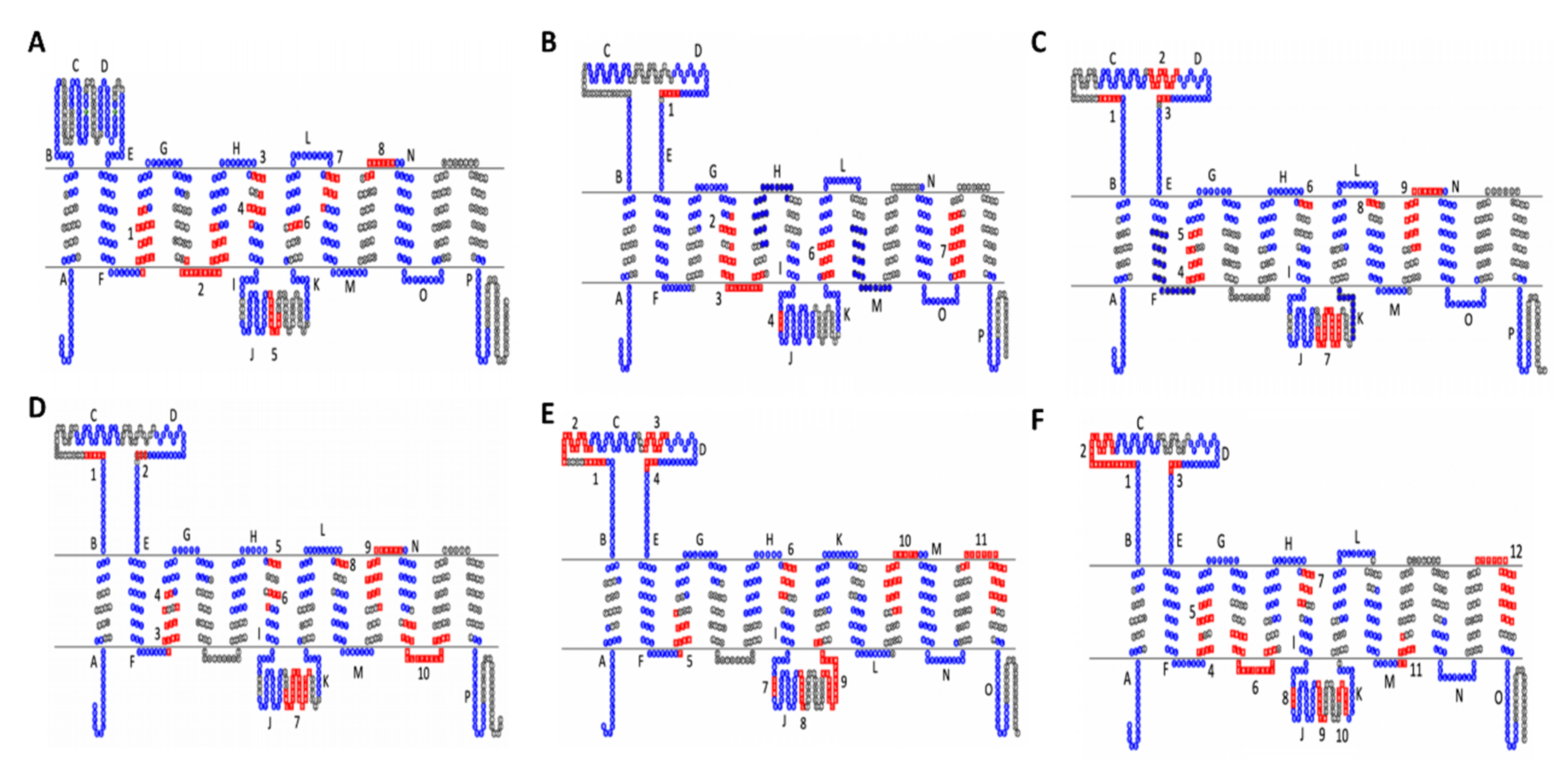
2.14. Sequence Similarity Study Suggests Novel Potential Functions to Explore and Possible Tertiary Structure of SLC22
3. Discussion
4. Materials and Methods
4.1. Data Collection
4.2. Sequence Alignment and Guide-Trees
4.3. Motif Analysis
4.4. SNP, Mutation, In-Vitro, Knockout, and Drug Transport Data
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SLC22 | Solute Carrier Family 22 |
| OAT | Organic Anion Transporter |
| OCT | Organic Cation Transporter |
| OCTN | Organic Zwitterion Transporter |
| GWAS | Genome Wide Association Study |
| SNP | Single Nucleotide Polymorphism |
| EGT | Ergothioneine |
| TMD | Transmembrane Domain |
| ECD | Extracellular Domain |
| ICD | Intracellular Domain |
| MFS | Major Facilitator Superfamily |
| ABCG2 | ATP-Binding Cassette Subfamily G, Member 2 |
| MSA | Multiple Sequence Alignment |
| MPP+ | 1-methyl-4-phenylpyridinium |
| TEA | tetraethylammonium |
| RSST | Remote Sensing and Signaling Theory |
References
- Hediger, M.A.; Clémençon, B.; Burrier, R.E.; Bruford, E.A. The ABCs of membrane transporters in health and disease (SLC series): Introduction. Mol. Asp. Med. 2013, 34, 95–107. [Google Scholar] [CrossRef]
- Lamhonwah, A.M.; Hawkins, C.E.; Tam, C.; Wong, J.; Mai, L.; Tein, I. Expression patterns of the organic cation/carnitine transporter family in adult murine brain. Brain Dev. 2008, 30, 31–42. [Google Scholar] [CrossRef]
- Sager, G.; Smaglyukova, N.; Fuskevaag, O.M. The role of OAT2 (SLC22A7) in the cyclic nucleotide biokinetics of human erythrocytes. J. Cell. Physiol. 2018, 233, 5972–5980. [Google Scholar] [CrossRef]
- Minuesa, G.; Purcet, S.; Erkizia, I.; Molina-Arcas, M.; Bofill, M.; Izquierdo-Useros, N.; Casado, J.F.; Clotet, B.; Pastor-Anglada, M.; Martinez-Picado, J. Expression and functionality of anti-human immunodeficiency virus and anticancer drug uptake transporters in immune cells. J. Pharmacol. Exp. Ther. 2008, 324, 558–567. [Google Scholar] [CrossRef]
- César-Razquin, A.; Snijder, B.; Frappier-Brinton, T.; Hediger, M.A.; Edwards, A.M.; Superti-Furga, G. A Call for Systematic Research on Solute Carriers. Cell 2015, 162, 478–487. [Google Scholar]
- Lelandais-Brière, C.; Jovanovic, M.; Torres, G.A.M.; Perrin, Y.; Lemoine, R.; Corre-Menguy, F.; Hartmann, C. Disruption of AtOCT1, an organic cation transporter gene, affects root development and carnitine-related responses in Arabidopsis. Plant J. 2007, 51, 154–164. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar]
- Lopez-Nieto, C.E.; You, G.; Bush, K.T.; Barros, E.J.G.; Beier, D.R.; Nigam, S.K. Molecular cloning and characterization of NKT, a gene product related to the organic cation transporter family that is almost exclusively expressed in the kidney. J. Biol. Chem. 1997, 272, 6471–6478. [Google Scholar] [CrossRef]
- Saier, M.H.; Reddy, V.S.; Tamang, D.S.; Västermark, Å. The transporter classification database. Nucleic Acids Res. 2014, 42, D251–D258. [Google Scholar] [CrossRef]
- Zhu, C.; Nigam, K.B.; Date, R.C.; Bush, K.T.; Springer, S.A.; Saier, M.H.; Wu, W.; Nigam, S.K. Evolutionary analysis and classification of OATs, OCTs, OCTNs, and other SLC22 transporters: Structure-function implications and analysis of sequence motifs. PLoS ONE 2015, 10, e0140569. [Google Scholar] [CrossRef]
- Nigam, S.K. The SLC22 Transporter Family: A Paradigm for the Impact of Drug Transporters on Metabolic Pathways, Signaling, and Disease. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 663–687. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Nigam, S.K. Toward a systems level understanding of organic anion and other multispecific drug transporters: A remote sensing and signaling hypothesis. Mol. Pharmacol. 2009, 76, 481–490. [Google Scholar] [CrossRef]
- Wu, W.; Dnyanmote, A.V.; Nigam, S.K. Remote communication through solute carriers and ATP binding cassette drug transporter pathways: An update on the Remote Sensing and Signaling Hypothesis. Mol. Pharmacol. 2011, 79, 795–805. [Google Scholar] [CrossRef]
- Nigam, S.K.; Bush, K.T.; Martovetsky, G.; Ahn, S.Y.; Liu, H.C.; Richard, E.; Bhatnagar, V.; Wu, W. The organic anion transporter (OAT) family: A systems biology perspective. Physiol. Rev. 2015, 95, 83–123. [Google Scholar] [CrossRef]
- Nigam, S.K. What do drug transporters really do? Nat. Rev. Drug Discov. 2014, 14, 29–44. [Google Scholar] [CrossRef]
- Rosenthal, S.B.; Bush, K.T.; Nigam, S.K. A Network of SLC and ABC Transporter and DME Genes Involved in Remote Sensing and Signaling in the Gut-Liver-Kidney Axis. Sci. Rep. 2019, 9, 11879. [Google Scholar] [CrossRef]
- Bhatnagar, V.; Richard, E.L.; Wu, W.; Nievergelt, C.M.; Lipkowitz, M.S.; Jeff, J.; Maihofer, A.X.; Nigam, S.K. Analysis of ABCG2 and other urate transporters in uric acid homeostasis in chronic kidney disease: Potential role of remote sensing and signaling. Clin. Kidney J. 2016, 9, 444–453. [Google Scholar] [CrossRef]
- Yano, H.; Tamura, Y.; Kobayashi, K.; Tanemoto, M.; Uchida, S. Uric acid transporter ABCG2 is increased in the intestine of the 5/6 nephrectomy rat model of chronic kidney disease. Clin. Exp. Nephrol. 2014, 18, 50–55. [Google Scholar] [CrossRef]
- Nigam, S.K.; Bhatnagar, V. The systems biology of uric acid transporters: The role of remote sensing and signaling. Curr. Opin. Nephrol. Hypertens. 2018, 27, 305–313. [Google Scholar] [CrossRef]
- Langelueddecke, C.; Roussa, E.; Fenton, R.; Wolff, N.; Lee, W.K.; Thévenod, F. Lipocalin-2 (24p3/neutrophil gelatinase-associated lipocalin (NGAL)) receptor is expressed in distal nephron and mediates protein endocytosis. J. Biol. Chem. 2012, 287, 159–169. [Google Scholar] [CrossRef]
- Kaler, G.; Truong, D.M.; Sweeney, D.E.; Logan, D.W.; Nagle, M.; Wu, W.; Eraly, S.A.; Nigam, S.K. Olfactory mucosa-expressed organic anion transporter, Oat6, manifests high affinity interactions with odorant organic anions. Biochem. Biophys. Res. Commun. 2006, 351, 872–876. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.L.; Ideker, T. Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef]
- Bremer, J. Carnitine. Metabolism and functions. Physiol. Rev. 1983, 63, 1420–1480. [Google Scholar] [CrossRef]
- Blumberg, A.L.; Denny, S.E.; Marshall, G.E.; Needleman, P. Blood vessel hormone interactions: Angiotensin, bradykinin, and prostaglandins. Am. J. Physiol. 1977, 232, H305–H310. [Google Scholar] [CrossRef]
- Wikoff, W.R.; Nagle, M.A.; Kouznetsova, V.L.; Tsigelny, I.F.; Nigam, S.K. Untargeted metabolomics identifies enterobiome metabolites and putative uremic toxins as substrates of organic anion transporter 1 (Oat1). J. Proteome Res. 2011, 10, 2842–2851. [Google Scholar] [CrossRef]
- Vallon, V.; Eraly, S.A.; Rao, S.R.; Gerasimova, M.; Rose, M.; Nagle, M.; Anzai, N.; Smith, T.; Sharma, K.; Nigam, S.K.; et al. A role for the organic anion transporter OAT3 in renal creatinine secretion in mice. Am. J. Physiol. Ren. Physiol. 2012, 302, F1293–F1299. [Google Scholar] [CrossRef]
- Wu, W.; Bush, K.T.; Nigam, S.K. Key Role for the Organic Anion Transporters, OAT1 and OAT3, in the in vivo Handling of Uremic Toxins and Solutes. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Kimura, H.; Takeda, M.; Narikawa, S.; Enomoto, A.; Ichida, K.; Endou, H. Human organic anion transporters and human organic cation transporters mediate renal transport of prostaglandins. J. Pharmacol. Exp. Ther. 2002, 301, 293–298. [Google Scholar] [CrossRef]
- Bush, K.T.; Wu, W.; Lun, C.; Nigam, S.K. The drug transporter OAT3 (SLC22A8) and endogenous metabolite communication via the gut–liver– kidney axis. J. Biol. Chem. 2017, 292, 15789–15803. [Google Scholar] [CrossRef]
- Wu, W.; Bush, K.T.; Liu, H.C.; Zhu, C.; Abagyan, R.; Nigam, S.K. Shared ligands between organic anion transporters (OAT1 and OAT6) and odorant receptors. Drug Metab. Dispos. 2015, 43, 1855–1863. [Google Scholar] [CrossRef]
- Monte, J.C.; Nagle, M.A.; Eraly, S.A.; Nigam, S.K. Identification of a novel murine organic anion transporter family member, OAT6, expressed in olfactory mucosa. Biochem. Biophys. Res. Commun. 2004, 323, 429–436. [Google Scholar] [CrossRef]
- Bien, S.A.; Su, Y.R.; Conti, D.V.; Harrison, T.A.; Qu, C.; Guo, X.; Lu, Y.; Albanes, D.; Auer, P.L.; Banbury, B.L.; et al. Genetic variant predictors of gene expression provide new insight into risk of colorectal cancer. Hum. Genet. 2019, 138, 307–326. [Google Scholar] [CrossRef]
- Wu, X.; Kekuda, R.; Huang, W.; Fei, Y.J.; Leibach, F.H.; Chen, J.; Conway, S.J.; Ganapathy, V. Identity of the organic cation transporter OCT3 as the extraneuronal monoamine transporter (uptake2) and evidence for the expression of the transporter in the brain. J. Biol. Chem. 1998, 273, 32776–32786. [Google Scholar] [CrossRef]
- Shen, H.; Liu, T.; Morse, B.L.; Zhao, Y.; Zhang, Y.; Qiu, X.; Chen, C.; Lewin, A.C.; Wang, X.T.; Liu, G.; et al. Characterization of Organic Anion Transporter 2 (SLC22A7): A Highly Efficient Transporter for Creatinine and Species-Dependent Renal Tubular Expression. Drug Metab. Dispos. 2015, 43, 984–993. [Google Scholar] [CrossRef]
- Köttgen, A.; Albrecht, E.; Teumer, A.; Vitart, V.; Krumsiek, J.; Hundertmark, C.; Pistis, G.; Ruggiero, D.; O’Seaghdha, C.M.; Haller, T.; et al. Genome-wide association analyses identify 18 new loci associated with serum urate concentrations. Nat. Genet. 2013, 45, 145–154. [Google Scholar] [CrossRef]
- Shin, S.Y.; Fauman, E.B.; Petersen, A.K.; Krumsiek, J.; Santos, R.; Huang, J.; Arnold, M.; Erte, I.; Forgetta, V.; Yang, T.P. An atlas of genetic influences on human blood metabolites. Nat. Genet. 2014, 46, 543–550. [Google Scholar] [CrossRef]
- Sun, W.; Wu, R.R.; Van Poelje, P.D.; Erion, M.D. Isolation of a family of organic anion transporters from human liver and kidney. Biochem. Biophys. Res. Commun. 2001, 283, 417–422. [Google Scholar] [CrossRef]
- Eraly, S.A.; Vallon, V.; Rieg, T.; Gangoiti, J.A.; Wikoff, W.R.; Siuzdak, G.; Barshop, B.A.; Nigam, S.K. Multiple organic anion transporters contribute to net renal excretion of uric acid. Physiol. Genom. 2008, 33, 180–192. [Google Scholar] [CrossRef]
- Hagos, Y.; Stein, D.; Ugele, B.; Burckhardt, G.; Bahn, A. Human renal organic anion transporter 4 operates as an asymmetric urate transporter. J. Am. Soc. Nephrol. 2007, 18, 430–439. [Google Scholar] [CrossRef]
- Maxwell, S.R.; Thomason, H.; Sandler, D.; Leguen, C.; Baxter, M.A.; Thorpe, G.H.; Jones, A.F.; Barnett, A.H. Antioxidant status in patients with uncomplicated insulin-dependent and non-insulin-dependent diabetes mellitus. Eur. J. Clin. Investig. 1997, 27, 484–490. [Google Scholar] [CrossRef]
- Sakiyama, M.; Matsuo, H.; Shimizu, S.; Nakashima, H.; Nakayama, A.; Chiba, T.; Naito, M.; Takada, T.; Suzuki, H.; Hamajima, N.; et al. A common variant of organic anion transporter 4 (OAT4/SLC22A11) gene is associated with renal underexcretion type gout. Drug Metab. Pharmacokinet. 2014, 29, 208–210. [Google Scholar] [CrossRef]
- Cha, S.H.; Sekine, T.; Kusuhara, H.; Yu, E.; Kim, J.Y.; Kim, D.K.; Sugiyama, Y.; Kanai, Y.; Endou, H. Molecular cloning and characterization of multispecific organic anion transporter 4 expressed in the placenta. J. Biol. Chem. 2000, 275, 4507–4512. [Google Scholar] [CrossRef]
- Tomi, M.; Eguchi, H.; Ozaki, M.; Tawara, T.; Nishimura, S.; Higuchi, K.; Maruyama, T.; Nishimura, T.; Nakashima, E. Role of OAT4 in uptake of estriol precursor 16α-hydroxydehydroepiandrosterone sulfate into human placental syncytiotrophoblasts from fetus. Endocrinology 2015, 156, 2704–2712. [Google Scholar] [CrossRef]
- Skwara, P.; Schömig, E.; Gründemann, D. A novel mode of operation of SLC22A11: Membrane insertion of estrone sulfate versus translocation of uric acid and glutamate. Biochem. Pharmacol. 2017, 128, 74–82. [Google Scholar] [CrossRef]
- Shiraya, K.; Hirata, T.; Hatano, R.; Nagamori, S.; Wiriyasermkul, P.; Jutabha, P.; Matsubara, M.; Muto, S.; Tanaka, H.; Asano, S.; et al. A novel transporter of SLC22 family specifically transports prostaglandins and co-localizes with 15-hydroxyprostaglandin dehydrogenase in renal proximal tubules. J. Biol. Chem. 2010, 285, 22141–22151. [Google Scholar] [CrossRef]
- Long, T.; Hicks, M.; Yu, H.C.; Biggs, W.H.; Kirkness, E.F.; Menni, C.; Zierer, J.; Small, K.S.; Mangino, M.; Messier, H.; et al. Whole-genome sequencing identifies common-to-rare variants associated with human blood metabolites. Nat. Genet. 2017, 49, 568–578. [Google Scholar] [CrossRef]
- Yee, S.W.; Stecula, A.; Chien, H.C.; Zou, L.; Feofanova, E.V.; Van Borselen, M.; Cheung, K.W.K.; Yousri, N.A.; Suhre, K.; Kinchen, J.M.; et al. Unraveling the functional role of the orphan solute carrier, SLC22A24 in the transport of 2 steroid conjugates through metabolomic and genome-wide association studies. PLoS Genet. 2019, 15, e1008208. [Google Scholar] [CrossRef]
- Shin, H.J.; Anzai, N.; Enomoto, A.; He, X.; Kim, D.K.; Endou, H.; Kanai, Y. Novel liver-specific organic anion transporter OAT7 that operates the exchange of sulfate conjugates for short chain fatty acid butyrate. Hepatology 2007, 45, 1046–1055. [Google Scholar] [CrossRef]
- Sardiello, M.; Annunziata, I.; Roma, G.; Ballabio, A. Sulfatases and sulfatase modifying factors: An exclusive and promiscuous relationship. Hum. Mol. Genet. 2005, 14, 3203–3217. [Google Scholar] [CrossRef]
- Papatheodorou, I.; Fonseca, N.A.; Keays, M.; Tang, Y.A.; Barrera, E.; Bazant, W.; Burke, M.; Füllgrabe, A.; Fuentes, A.M.P.; George, N.; et al. Expression Atlas: Gene and protein expression across multiple studies and organisms. Nucleic Acids Res. 2018, 46, D246–D251. [Google Scholar] [CrossRef]
- Bahn, A.; Hagos, Y.; Reuter, S.; Balen, D.; Brzica, H.; Krick, W.; Burckhardt, B.C.; Sabolić, I.; Burckhardt, G. Identification of a new urate and high affinity nicotinate transporter - human organic anion transporter 10 (hOAT10, SLC22A13. J. Biol. Chem. 2008, 283, 16332–16341. [Google Scholar] [CrossRef]
- Uhl, G.R.; Liu, Q.R.; Drgon, T.; Johnson, C.; Walther, D.; Rose, J.E.; David, S.P.; Niaura, R.; Lerman, C. Molecular genetics of successful smoking cessation: Convergent genome-wide association study results. Arch. Gen. Psychiatry 2008, 65, 683–693. [Google Scholar] [CrossRef]
- Uhl, G.R.; Drgon, T.; Li, C.Y.; Johnson, C.; Liu, Q.R. Smoking and smoking cessation in disadvantaged women: Assessing genetic contributions. Drug Alcohol Depend. 2009, 104, S58–S63. [Google Scholar] [CrossRef]
- Maruyama, S.Y.; Ito, M.; Ikami, Y.; Okitsu, Y.; Ito, C.; Toshimori, K.; Fujii, W.; Yogo, K. A critical role of solute carrier 22a14 in sperm motility and male fertility in mice. Sci. Rep. 2016, 6, 36468. [Google Scholar] [CrossRef]
- Bennett, K.M.; Liu, J.; Hoelting, C.; Stoll, J. Expression and analysis of two novel rat organic cation transporter homologs, SLC22A17 and SLC22A23. Mol. Cell. Biochem. 2011, 352, 143–154. [Google Scholar] [CrossRef]
- Wan-Jie, C.; Wei-Yi, O.; Gavin, D. Lipocalin 2 and lipocalin 2 receptor in neuron-glia interactions following injury. Front. Cell. Neurosci. 2016, 10. [Google Scholar] [CrossRef]
- Zhao, Z.Z.; Croft, L.; Nyholt, D.R.; Chapman, B.; Treloar, S.A.; Hull, M.L.; Montgomery, G.W. Evaluation of polymorphisms in predicted target sites for micro RNAs differentially expressed in endometriosis. Mol. Hum. Reprod. 2011, 17, 92–103. [Google Scholar] [CrossRef]
- Serrano León, A.; Amir Shaghaghi, M.; Yurkova, N.; Bernstein, C.N.; El-Gabalawy, H.; Eck, P. Single-nucleotide polymorphisms in SLC22A23 are associated with ulcerative colitis in a Canadian white cohort. Am. J. Clin. Nutr. 2014, 100, 289–294. [Google Scholar] [CrossRef]
- Aberg, K.; Adkins, D.E.; Liu, Y.; McClay, J.L.; Bukszár, J.; Jia, P.; Zhao, Z.; Perkins, D.; Stroup, T.S.; Lieberman, J.A.; et al. Genome-wide association study of antipsychotic-induced QTc interval prolongation. Pharmacogenomics J. 2012, 12, 165–172. [Google Scholar] [CrossRef]
- Nigam, A.K.; Li, J.G.; Lall, K.; Shi, D.; Bush, K.T.; Bhatnagar, V.; Abagyan, R.; Nigam, S.K. Unique metabolite preferences of the drug transporters OAT1 and OAT3 analyzed by machine learning. J. Biol. Chem. 2020, RA119, 010729. [Google Scholar] [CrossRef]
- Busch, A.E.; Quester, S.; Ulzheimer, J.C.; Gorboulev, V.; Akhoundova, A.; Waldegger, S.; Lang, F.; Koepsell, H. Monoamine neurotransmitter transport mediated by the polyspecific cation transport rOCT1. FEBS Lett. 1996, 395, 153–156. [Google Scholar] [CrossRef]
- Gründemann, D.; Liebich, G.; Kiefer, N.; Köster, S.; Schömig, E. Selective substrates for non-neuronal monoamine transporters. Mol. Pharmacol. 1999, 56, 1–10. [Google Scholar] [CrossRef]
- Urakami, Y.; Kimura, N.; Okuda, M.; Inui, K.I. Creatinine transport by basolateral organic cation transporter hOCT2 in the human kidney. Pharm. Res. 2004, 21, 976–981. [Google Scholar] [CrossRef]
- Enomoto, A.; Wempe, M.F.; Tsuchida, H.; Shin, H.J.; Cha, S.H.; Anzai, N.; Goto, A.; Sakamoto, A.; Niwa, T.; Kanai, Y.; et al. Molecular identification of a novel carnitine transporter specific to human testis: Insights into the mechanism of carnitine recognition. J. Biol. Chem. 2002, 277, 36262–36271. [Google Scholar] [CrossRef]
- Ohashi, R.; Tamai, I.; Yabuuchi, H.; Nezu, J.I.; Oku, A.; Sai, Y.; Shimane, M.; Tsuji, A. Na+-dependent carnitine transport by organic cation transporter (OCTN2): Its pharmacological and toxicological relevance. J. Pharmacol. Exp. Ther. 1999, 291, 778–784. [Google Scholar]
- Gründemann, D.; Harlfinger, S.; Golz, S.; Geerts, A.; Lazar, A.; Berkels, R.; Jung, N.; Rubbert, A.; Schömig, E. Discovery of the ergothioneine transporter. Proc. Natl. Acad. Sci. USA 2005, 102, 5256–5261. [Google Scholar] [CrossRef]
- Cheah, I.K.; Halliwell, B. Ergothioneine; antioxidant potential, physiological function and role in disease. Biochim. et Biophys. Acta Mol. Basis Dis. 2012, 1822, 784–793. [Google Scholar] [CrossRef]
- Ogden, T.H.; Rosenberg, M.S. Multiple sequence alignment accuracy and phylogenetic inference. Syst. Biol. 2006, 55, 314–328. [Google Scholar] [CrossRef]
- Wu, W.; Baker, M.E.; Eraly, S.A.; Bush, K.T.; Nigam, S.K. Analysis of a large cluster of SLC22 transporter genes, including novel USTs, reveals species-specific amplification of subsets of family members. Physiol. Genom. 2009, 38, 116–124. [Google Scholar] [CrossRef][Green Version]
- Yokoyama, H.; Anzai, N.; Ljubojevic, M.; Ohtsu, N.; Sakata, T.; Miyazaki, H.; Nonoguchi, H.; Islam, R.; Onozato, M.; Tojo, A.; et al. Functional and immunochemical characterization of a novel organic anion transporter Oat8 (Slc22a9) in rat renal collecting duct. Cell. Physiol. Biochem. 2008, 21, 269–278. [Google Scholar] [CrossRef]
- Eraly, S.A.; Monte, J.C.; Nigam, S.K. Novel slc22 transporter homologs in fly, worm, and human clarify the phylogeny of organic anion and cation transporters. Physiol. Genom. 2004, 18, 12–24. [Google Scholar] [CrossRef]
- Eraly, S.A.; Nigam, S.K. Novel human cDNAs homologous to Drosophila Orct and mammalian carnitine transporters. Biochem. Biophys. Res. Commun. 2002, 297, 1159–1166. [Google Scholar] [CrossRef]
- Quistgaard, E.M.; Löw, C.; Guettou, F.; Nordlund, P. Understanding transport by the major facilitator superfamily (MFS): Structures pave the way. Nat. Rev. Mol. Cell Biol. 2016, 17, 123–132. [Google Scholar] [CrossRef]
- Inana, G.; Piatigorsky, J.; Norman, B.; Slingsby, C.; Blundell, T. Gene and protein structure of a β-crystallin polypeptide in murine lens: Relationship of exons and structural motifs. Nature 1983, 302, 310–315. [Google Scholar] [CrossRef]
- Casper, J.; Zweig, A.S.; Villarreal, C.; Tyner, C.; Speir, M.L.; Rosenbloom, K.R.; Raney, B.J.; Lee, C.M.; Lee, B.T.; Karolchik, D.; et al. The UCSC Genome Browser database: 2018 update. Nucleic Acids Res. 2018, 46, D762–D769. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.; Potter, S.C.; Finn, R.D. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Abagyan, R.; Totrov, M.; Kuznetsov, D. ICM—A new method for protein modeling and design: Applications to docking and structure prediction from the distorted native conformation. J. Comput. Chem. 1994, 15, 488–506. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Reddy, V.S.; Saier, M.H. BioV Suite—A collection of programs for the study of transport protein evolution. FEBS J. 2012, 279, 2036–2046. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.F.; Waterman, M.S. Identification of common molecular subsequences. J. Mol. Biol. 1981, 147, 195–197. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar]
- Dobson, L.; Reményi, I.; Tusnády, G.E. CCTOP: A Consensus Constrained TOPology prediction web server. Nucleic Acids Res. 2015, 43, W408–W412. [Google Scholar] [CrossRef]
- Johns, S.J. TOPO2, Transmembrane Protein Display Software. Available online: http://www.sacs.ucsf.edu/TOPO2/ (accessed on 3 March 2020).
- Suhre, K.; Shin, S.Y.; Petersen, A.K.; Mohney, R.P.; Meredith, D.; Wägele, B.; Altmaier, E.; Deloukas, P.; Erdmann, J.; Grundberg, E.; et al. Human metabolic individuality in biomedical and pharmaceutical research. Nature 2011, 477, 54–62. [Google Scholar] [CrossRef]
- Kastenmüller, G.; Raffler, J.; Gieger, C.; Suhre, K. Genetics of human metabolism: An update. Human Mol. Genet. 2015, 24, R93–R101. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Former Groupings | Updated Groupings | ||
|---|---|---|---|
| Subclade | Members | Subgroup | Members |
| OAT | A6, A7, A8, A9, A10, A11, A12, A19, A20, A22, A24, A25, A26, A27, A28, A29, A30 | OATS1 | A6, A8, A20 |
| OATS2 | A7 | ||
| OATS3 | A11, A12, a22 | ||
| OATS4 1 | A9, A10, A24, A25 | ||
| OAT-like | A13, A14 | OAT-like | A13, A14 |
| OAT-related | A17, A18. A23, A31 | OAT-related | A17, A18. A23, A31 |
| OCTN-related | A15, A16 | OCTN/OCTN related | A4, A5, A15, A16, a21 |
| OCTN | A4, A5 | ||
| OCT | A1, A2, A3 | OCT | A1, A2, A3 |
| SLC22 Transporter | Common Name | # of Unique Drugs | # of Metabolites | Sum | Specificity | Metabolic Pathways |
|---|---|---|---|---|---|---|
| A1 | OCT1 | 70 | 15 | 85 | multi | Monoamines, carnitines, PG 1 |
| A2 | OCT2 | 84 | 24 | 108 | multi | Monoamines, carnitines, PG, creatinine |
| A3 | OCT3 | 40 | 12 | 52 | oligo | Monoamines, carnitines, creatinine |
| A4 | OCTN1 | 33 | 25 | 58 | oligo | Carnitines, ergothioneine |
| A5 | OCTN2 | 55 | 20 | 75 | oligo | Carnitines |
| A6 | OAT1 | 99 | 52 | 151 | multi | Uric acid, PG, gut microbiome derived products, TCA 2 |
| A7 | OAT2 | 35 | 16 | 51 | oligo | Cyclic nucleotides, PG, carnitine, creatinine, TCA |
| A8 | OAT3 | 126 | 88 | 214 | multi | Uric acid, PG, creatinine, gut microbiome derived products, TCA, bile acids |
| A9 | OAT7 | 0 | 9 | 9 | oligo | Conjugated sex steroids, SCFA 3 |
| A10 | OAT5 | 3 | 2 | 5 | mono | Conjugated sex steroids |
| A11 | OAT4 | 42 | 9 | 51 | oligo | Uric acid, PG, conjugated sex steroids |
| A12 | URAT1 | 4 | 7 | 11 | mono | Uric acid, TCA |
| A13 | OAT10, ORCTL3 | n/a | 13 | 13 | mono | Uric acid, TCA |
| A14 | ORCTL4 | n/a | n/a | n/a | n/a | Understudied |
| A15 | FLIPT1 | n/a | 7 | 7 | mono | EGT, complex lipids |
| A16 | FLIPT2, CT2 | 2 | 16 | 18 | oligo | Carnitines, EGT |
| A17 | BOCT1, NGAL, Lcn2-R | n/a | 2 | 2 | mono | Lipocalin |
| A18 | SLC22A1L, TSSC5, | n/a | 2 | 2 | n/a | Understudied |
| A20 | OAT6 | n/a | 13 | 13 | oligo | Odorants, SCFA |
| a21 | Octn3, Slc22a9 | n/a | 1 | 1 | mono | Carnitine |
| a22 | OAT-PG | n/a | 12 | 12 | mono | PG, conjugated sex steroids |
| A23 | BOCT2 | n/a | 12 | 12 | oligo | Fatty acids |
| A24 | n/a | n/a | 10 | 10 | oligo | Conjugated sex steroids, bile acids |
| A25 | UST6 | n/a | 1 | 1 | mono | Conjugated sex steroids |
| A31 | n/a | n/a | n/a | n/a | n/a | Understudied |
| Genomic Loci | Tissue Expression | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Subgroup | SLC22 Transporter | Common Name | Human Chr. | Mouse Chr. | Liver | Kidney | Brain | Gut | Heart | Lung | Testis | Immune Cell | Bone Marrow | Placenta |
| OATS1 | SLC22A6 | OAT1 | 11 | 19 | ✓ | ✓ | ||||||||
| SLC22A8 | OAT3 | 11 | 19 | ✓ | ✓ | |||||||||
| SLC22A20 | OAT6 | 11 | 19 | ✓ | ✓(m) | ✓ | ||||||||
| OATS2 | SLC22A7 | OAT2 | 6 | 17 | ✓ | ✓ | ✓ | |||||||
| OATS3 | SLC22A11 | OAT4 | 11 | n/a | ✓ | ✓ | ||||||||
| SLC22A12 | URAT1 | 11 | 19 | ✓ | ||||||||||
| Slc22a22 | OAT-PG | - | 15 | ✓(m) | ||||||||||
| OATS4 | SLC22A9 | OAT7 | 11 | - | ✓ | ✓ | ✓ | |||||||
| SLC22A10 | OAT5 | 11 | - | ✓ | ✓ | |||||||||
| SLC22A24 | n/a | 11 | - | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| SLC22A25 | UST6 | 11 | - | ✓ | ||||||||||
| OAT-like | SLC22A13 | OAT10, ORCTL3 | 3 | 9 | ✓ | ✓ | ✓ | ✓ | ||||||
| SLC22A14 | ORCTL4 | 3 | 9 | ✓ | ✓ | ✓ | ||||||||
| OAT-related | SLC22A17 | BOCT1, NGAL, Lcn2-R | 14 | 14 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||
| SLC22A18 | SLC22A1L, TSSC5 | 11 | 7 | ✓ | ✓ | ✓ | ✓ | |||||||
| SLC22A23 | BOCT2 | 6 | 13 | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| SLC22A31 | n/a | 16 | - | ✓ | ✓ | |||||||||
| OCTN/OCTN-related | SLC22A4 | OCTN1 | 5 | 11 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||
| SLC22A5 | OCTN2 | 5 | 11 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||
| SLC22A15 | FLIPT1 | 6 | 10 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||
| SLC22A16 | FLIPT2, CT2 | 1 | 3 | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| Slc22a21 | Octn3, Slc22a9 | - | 11 | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| OCT | SLC22A1 | OCT1 | 6 | 17 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||
| SLC22A2 | OCT2 | 6 | 17 | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| SLC22A3 | OCT3 | 6 | 17 | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||
| Gene | Metabolite | Source | Metrics | Citation |
|---|---|---|---|---|
| SLC22A9 | butyrate | in vitro, Xenopus oocytes | trans-stimulates transport p < 0.001 | [37] |
| SLC22A9 | dehydroepiandrosterone sulfate | in vitro, Xenopus oocytes | Km: 2.2 uM | [37] |
| SLC22A9 | estrone sulfate | in vitro, Xenopus oocytes | Km: 8.7 uM | [37] |
| SLC22A9 | etiocholanolone glucuronide | GWAS, rs113747568 | p = 5.27 × 10−28 | [47] |
| SLC22A9 | nicotinate | in vitro, Xenopus oocytes | trans-stimulates transport p < 0.01 | [37] |
| SLC22A9 | progesterone | GWAS, rs112295236 | p = 8.00 × 10−12 | [47] |
| SLC22A9 | propionate | in vitro, Xenopus oocytes | trans-stimulates transport p < 0.01 | [37] |
| SLC22A9 | tyramine o-sulfate | GWAS, rs397740636 | p = 2.06 × 10−6 | [47] |
| SLC22A9 | valerate | in vitro, Xenopus oocytes | trans-stimulates transport p < 0.001 | [37] |
| SLC22A10 | epiandrosterone sulfate | GWAS, rs1939769 | p = 2.06 × 10−7 | [37] |
| SLC22A10 | etiocholanolone glucuronide | GWAS, rs112753913 | p = 1.88 × 10−27 | [47] |
| SLC22A24 | androstanediol glucuronide | in vitro, HEK293 Flp-In | IC50: 21 ± 11 uM | [48] |
| SLC22A24 | chenodeoxycholate | in vitro, HEK293 Flp-In | IC50: 2.6 ± 1.0 uM | [48] |
| SLC22A24 | estradiol glucuronide | in vitro, HEK293 Flp-In | 3-5 fold over vector control | [48] |
| SLC22A24 | estrone sulfate | in vitro, HEK293 Flp-In | 5-10 fold over vector control | [48] |
| SLC22A24 | etiocholanolone glucuronide | in vitro, HEK293 Flp-In | IC50: 29 ± 4.7 uM | [48] |
| SLC22A24 | etiocholanolone glucuronide | GWAS, rs113532193 | p = 5.90 × 10−37 | [47] |
| SLC22A24 | pregnanediol-3-glucuronide | in vitro, HEK293 Flp-In | IC50: >200 uM | [48] |
| SLC22A24 | pregnanediol-3-glucuronide | GWAS, rs202187460 | p = 5.91 × 10−7 | [47] |
| SLC22A24 | pregnenolone sulfate | in vitro, HEK293 Flp-In | IC50: 1.4 ± 0.1 uM | [48] |
| SLC22A24 | progesterone | in vitro, HEK293 Flp-In | IC50: 7.4 ± 3.0 uM | [48] |
| SLC22A24 | taurocholic acid | in vitro, HEK293 Flp-In | 10–20 fold over vector control | [48] |
| SLC22A24 | ursodeoxycholate | in vitro, HEK293 Flp-In | IC50: 7.6 ± 1.2 uM | [48] |
| SLC22A25 | etiocholanolone glucuronide | GWAS, rs113950742 | p = 4.12 × 10−27 | [47] |
| Subclade | SLC22 Family Member | Common Name | Non-SLC22 Protein | Identity Shared (%) | pP Value |
|---|---|---|---|---|---|
| OCT | hSLC22A1 ECD | OCT1 | hSCO-spondin | 28.97 | 5.47 |
| bSCO-spondin | 30.84 | 5.35 | |||
| mSCO-spondin | 24.3 | 6.24 | |||
| rSCO-spondin | 24.3 | 6.16 | |||
| hSLC22A2 ECD | OCT2 | bSCO-spondin | 30.84 | 5.29 | |
| mSCO-spondin | 25.23 | 5.92 | |||
| rSCO-spondin | 24.3 | 5.61 | |||
| hSLC22A3 ECD | OCT3 | hSCO-spondin | 27.43 | 5.33 | |
| bSCO-spondin | 22.12 | 5.49 | |||
| mSCO-spondin | 27.43 | 5.89 | |||
| rSCO-spondin | 25.66 | 5.81 | |||
| OAT-related | hSLC22A31 ECD | n/a | hRBM42 | 30.95 | 5.95 |
| bRBM42 | 32.14 | 6.05 | |||
| mRBM42 | 30.95 | 5.95 | |||
| rRBM42 | 30.95 | 5.95 | |||
| hBAHD1 | 36.9 | 5.39 | |||
| OCTN | mSlc22a16 ECD | FLIPT2, CT2 | gCRBB3 | 26 | 5.4 |
| hSLC22A16 | hTAS2R41 | 20 | 5.3 | ||
| hSLC22A5 | OCTN2 | GPR160 | 21 | 6.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Engelhart, D.C.; Granados, J.C.; Shi, D.; Saier Jr., M.H., Jr.; Baker, M.E.; Abagyan, R.; Nigam, S.K. Systems Biology Analysis Reveals Eight SLC22 Transporter Subgroups, Including OATs, OCTs, and OCTNs. Int. J. Mol. Sci. 2020, 21, 1791. https://doi.org/10.3390/ijms21051791
Engelhart DC, Granados JC, Shi D, Saier Jr. MH Jr., Baker ME, Abagyan R, Nigam SK. Systems Biology Analysis Reveals Eight SLC22 Transporter Subgroups, Including OATs, OCTs, and OCTNs. International Journal of Molecular Sciences. 2020; 21(5):1791. https://doi.org/10.3390/ijms21051791
Chicago/Turabian StyleEngelhart, Darcy C., Jeffry C. Granados, Da Shi, Milton H. Saier Jr., Jr., Michael E. Baker, Ruben Abagyan, and Sanjay K. Nigam. 2020. "Systems Biology Analysis Reveals Eight SLC22 Transporter Subgroups, Including OATs, OCTs, and OCTNs" International Journal of Molecular Sciences 21, no. 5: 1791. https://doi.org/10.3390/ijms21051791
APA StyleEngelhart, D. C., Granados, J. C., Shi, D., Saier Jr., M. H., Jr., Baker, M. E., Abagyan, R., & Nigam, S. K. (2020). Systems Biology Analysis Reveals Eight SLC22 Transporter Subgroups, Including OATs, OCTs, and OCTNs. International Journal of Molecular Sciences, 21(5), 1791. https://doi.org/10.3390/ijms21051791