Cloning, Molecular Characterization and Expression Patterns of DMRTC2 Implicated in Germ Cell Development of Male Tibetan Sheep

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Cloning and Sequence Analysis of Tibetan Sheep DMRTC2 CDS
2.2. Molecular Characteristics of Tibetan Sheep DMRTC2
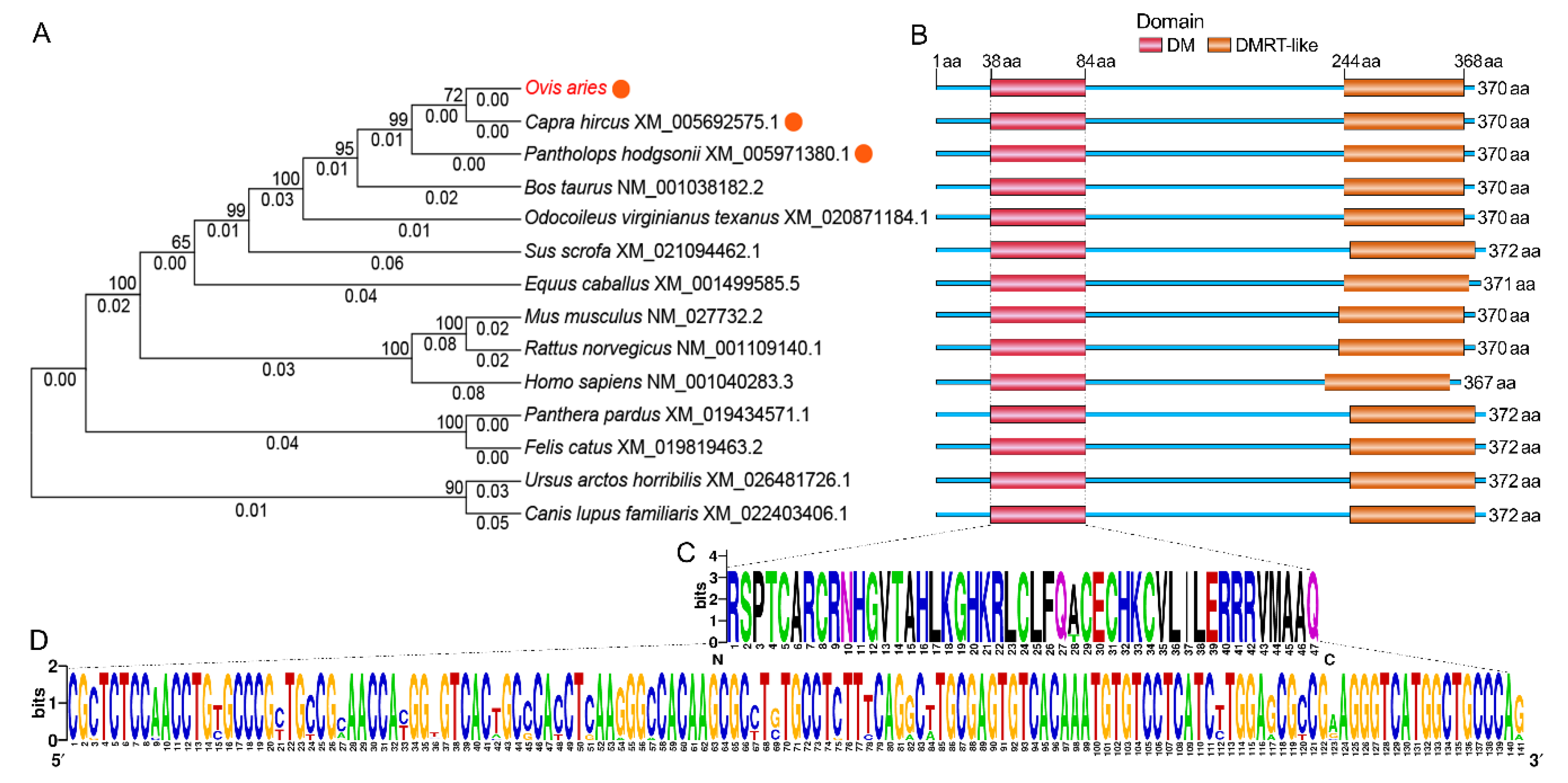
2.3. The Spatial Structures of Tibetan Sheep DMRTC2 and Evolutionary Relationships Between Sheep and Other Known Mammals
2.4. Expression Patterns of DMRTC2 at the Transcript and Protein Levels
2.5. Localization of DMRTC2 Protein in Developmental Tibetan Sheep Testes
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Collection
4.2. RNA Isolation and cDNA Synthesis
4.3. Full-length cDNA Cloning of DMRTC2 Gene
4.4. Bioinformatics Analysis
4.5. Quantitative Real-time PCR (qPCR)
4.6. Western Blot
4.7. Histologic and Immunofluorescence Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DMRTC2 | Double sex and mab-3-related transcription factors like family C2 |
| ORF | Open reading frame |
| qPCR | Quantitative real-time PCR |
| RIPA | Radio immunoprecipitation assay |
| BCA | Bicinchoninic acid |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
References
- Hao, S.; Ni, F.; Yang, W. The dynamics and regulation of chromatin remodeling during spermiogenesis. Gene 2019, 706, 201–210. [Google Scholar] [CrossRef]
- Lee, J. Roles of cohesin and condensin in chromosome dynamics during mammalian meiosis. J. Reprod. Dev. 2013, 59, 431–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Xu, Z.; Khawar, M.; Liu, C.; Li, W. The histone codes for meiosis. Reproduction 2017, 154, R65–R79. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.; Cai, T.; Lin, X.; Wu, Y.; Wang, X.; Yang, F.; Han, C. Integrative proteomic and transcriptomic analyses reveal multiple post-transcriptional regulatory mechanisms of mouse spermatogenesis. Mol. Cell. Proteomics 2013, 12, 1144–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalmel, F.; Rolland, A. Linking transcriptomics and proteomics in spermatogenesis. Reproduction 2015, 150, R149–R157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, S. Retinoic Acid Receptor Alpha in Germ Cells is Important for Mitosis of Spermatogonia, Spermatogonial Differentiation and Meiosis. Ph.D. Thesis, Washington State University, Pullman, WA, USA, 2013. [Google Scholar]
- Kawamata, M.; Nishimori, K. Mice deficient in Dmrt7 show infertility with spermatogenic arrest at pachytene stage. FEBS Lett. 2006, 580, 6442–6446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, P.; Xiang, L.; Guo, X.; Bao, P.; Jin, S.; Wu, X. The low expression of Dmrt7 is associated with spermatogenic arrest in cattle-yak. Mol. Biol. Rep. 2014, 41, 7255–7263. [Google Scholar] [CrossRef]
- Date, S.; Nozawa, O.; Inoue, H.; Hidema, S.; Nishimori, K. Impairment of pachytene spermatogenesis in Dmrt7 deficient mice, possibly causing meiotic arrest. Biosci. Biotechnol. Biochem. 2012, 76, 1621–1626. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Zhou, X.; Liu, J.; Yuan, J.; Cheng, H.; Zhou, R. Nuclear factor-Y (NF-Y) regulates transcription of mouse Dmrt7 gene by binding to tandem CCAAT boxes in its proximal promoter. Int. J. Biol. Sci. 2010, 6, 655–664. [Google Scholar] [CrossRef]
- Kim, S.; Namekawa, S.; Niswander, L.; Ward, J.; Lee, J.; Bardwell, V.; Zarkower, D. A mammal-specific Doublesex homolog associates with male sex chromatin and is required for male meiosis. PLoS Genet. 2007, 3, e62. [Google Scholar] [CrossRef]
- Kawamata, M.; Inoue, H.; Nishimori, K. Male-specific function of Dmrt7 by sexually dimorphic translation in mouse testis. Sex. Dev. 2007, 1, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Poulain, M.; Frydman, N.; Tourpin, S.; Muczynski, V.; Mucsynski, V.; Souquet, B.; Benachi, A.; Habert, R.; Rouiller-Fabre, V.; Livera, G. Involvement of doublesex and mab-3-related transcription factors in human female germ cell development demonstrated by xenograft and interference RNA strategies. Mol. Human Reprod. 2014, 20, 960–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gegenschatz-Schmid, K.; Verkauskas, G.; Demougin, P.; Bilius, V.; Dasevicius, D.; Stadler, M.; Hadziselimovic, F. DMRTC2, PAX7, BRACHYURY/T and TERT are implicated in male germ cell development following curative hormone treatment for cryptorchidism-induced infertility. Genes 2017, 8, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wang, W.; Jing, X.; Degen, A.; Guo, Y.; Kang, J.; Shang, Z.; Yu, Z.; Qiu, Q.; Guo, X.; et al. Effect of dietary energy on digestibilities, rumen fermentation, urinary purine derivatives and serum metabolites in Tibetan and small-tailed Han sheep. J. Anim. Physiol. Anim. Nutr. 2019, 103, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Guo, X.; Bao, P.; Liang, C.; Wu, X.; Liu, J.; Yan, P. Sequence analysis and study on the expression level of Dmrt7 gene in yak and cattle-yak testis. Sci. Agric. Sin. 2013, 46, 1036–1043. [Google Scholar] [CrossRef]
- Zhang, T.; Zarkower, D. DMRT proteins and coordination of mammalian spermatogenesis. Stem Cell Res. 2017, 24, 195–202. [Google Scholar] [CrossRef]
- Matson, C.; Zarkower, D. Sex and the singular DM domain: Insights into sexual regulation, evolution and plasticity. Nat. Rev. Genet. 2012, 13, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.; Zarkower, D.; Bardwell, V. Vertebrate DM domain proteins bind similar DNA sequences and can heterodimerize on DNA. BMC Mol. Biol. 2007, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Zarkower, D. DMRT genes in vertebrate gametogenesis. Curr. Top. Dev. Biol. 2013, 102, 327–356. [Google Scholar] [CrossRef]
- Wu, S.; Ji, L.; Zhang, Y.; Du, P.; Kang, D.; Ha, F.; Miao, Y. Cloning, molecular characterization and tissue expression profile of Binglangjiang buffalo gene Dmrt7. J. Yunnan Agric. Univ. (Nat. Sci.) 2017, 32, 612–619. [Google Scholar] [CrossRef]
- Tsend-Ayush, E.; Lim, S.; Pask, A.; Hamdan, D.; Renfree, M.; Grützner, F. Characterisation of ATRX, DMRT1, DMRT7 and WT1 in the platypus (Ornithorhynchus anatinus). Reprod. Fertil. Dev. 2009, 21, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.; Vormer, T.; Jongejan, A.; Röling, M.; Silber, S.; de Rooij, D.; Hamer, G.; Repping, S.; van Pelt, A. Unraveling transcriptome dynamics in human spermatogenesis. Development 2017, 144, 3659–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.; Geer, R.; Gonzales, N.; Gwadz, M.; Hurwitz, D.; Marchler, G.; Song, J.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, T.; Lu, Z.; Luo, R.; Gao, J.; Zhao, X.; Ma, Y. Expression and cellular localization of double sex and mab-3 related transcription factor 1 in testes of postnatal Small-Tail Han sheep at different developmental stages. Gene 2017, 642, 467–473. [Google Scholar] [CrossRef]
- Li, T.; Wang, X.; Zhang, H.; Chen, H.; Liu, N.; Xue, R.; Zhao, X.; Ma, Y. Gene expression patterns and protein cellular localization suggest a novel role for DAZL in developing Tibetan sheep testes. Gene 2020, 731, 144335. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession no. | Primer Sequence (5′–3′) | Size (bp) | Purpose |
|---|---|---|---|---|
| DMRTC2 | XM_027978391.1 | F: ATGGACCCCAATGAAATGCC | 1178 | cDNA cloning |
| R: CTAGCTTAGGAGGGAAATGG | ||||
| DMRTC2 | XM_027978391.1 | F: CTGAGGCTCTTGTGGGACTG | 96 | qPCR |
| R:CACAAGGATGGAGCAGGGAG | ||||
| β-actin | NM_001009784.2 | F: CTTCCAGCCTTCCTTCCTGG | 180 | qPCR |
| R: GCCAGGGCAGTGATCTCTTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Zhang, H.; Wang, X.; Yin, D.; Chen, N.; Kang, L.; Zhao, X.; Ma, Y. Cloning, Molecular Characterization and Expression Patterns of DMRTC2 Implicated in Germ Cell Development of Male Tibetan Sheep. Int. J. Mol. Sci. 2020, 21, 2448. https://doi.org/10.3390/ijms21072448
Li T, Zhang H, Wang X, Yin D, Chen N, Kang L, Zhao X, Ma Y. Cloning, Molecular Characterization and Expression Patterns of DMRTC2 Implicated in Germ Cell Development of Male Tibetan Sheep. International Journal of Molecular Sciences. 2020; 21(7):2448. https://doi.org/10.3390/ijms21072448
Chicago/Turabian StyleLi, Taotao, Hongyu Zhang, Xia Wang, De′en Yin, Nana Chen, Lingyun Kang, Xingxu Zhao, and Youji Ma. 2020. "Cloning, Molecular Characterization and Expression Patterns of DMRTC2 Implicated in Germ Cell Development of Male Tibetan Sheep" International Journal of Molecular Sciences 21, no. 7: 2448. https://doi.org/10.3390/ijms21072448