Nasal Epithelial Cells Activated with Alternaria and House Dust Mite Induce Not Only Th2 but Also Th1 Immune Responses



Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics and Chemical Mediators in CRS with Nasal Polyps (CRSwNPs)
2.2. Production of Chemical Mediators from Nasal Epithelial Cells by Airborne Allergens
2.3. Expression of Transcription Factors from Nasal Epithelial Cells by Airborne Allergens
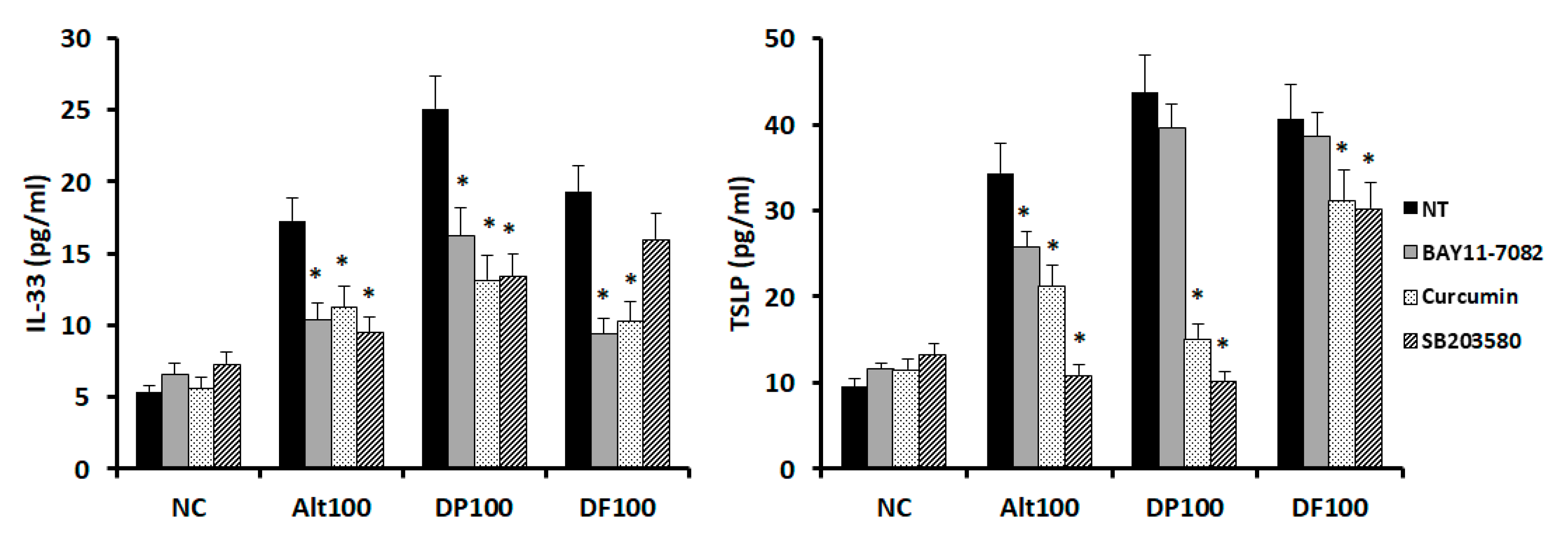
2.4. Effects of Transcription Factor Inhibitors on Chemical Mediator Production from Nasal Epithelial Cells
2.5. Effects of Nasal Epithelial Cell-Conditioned Media (NECM) on Peripheral Blood Mononuclear Cells (PBMCs) and Innate Lymphoid Type 2 Cell (ILC2) Expressions
3. Discussion
4. Materials and Methods
4.1. Chemical Mediators in CRSwNP
4.2. Stimulation of Primary Nasal Epithelial Cells with Airborne Allergens
4.3. Western Blot Analysis of Nasal Epithelial Cells for Transcription Factors
4.4. Effects of Transcription Factor Inhibitors on Cytokine Production
4.5. Effects of Nasal Epithelial Cell Conditioned Media (NECM) on Peripheral Blood Mononuclear Cells (PBMCs) and Innate Lymphoid Type 2 Cell (ILC2s) Expressions
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CRS | Chronic rhinosinusitis |
| IL | Interleukin |
| TSLP | Thymic stromal lymphopoietin |
| HDM | House dust mite |
| DP | Dermatophagoides pteronyssinus |
| DF | Dermatophagoides farina |
| NF-κB | Nuclear factor kappa B |
| AP-1 | Activator protein 1 |
| MAPK | Mitogen-activated protein kinase |
| NECM | Nasal epithelial cell-conditioned media |
| IFN-γ | Interferon gamma |
| PBMC | Peripheral blood mononuclear cell |
| ILC2 | Innate lymphoid type 2 cell |
References
- Roan, F.; Obata-Ninomiya, K.; Ziegler, S.F. Epithelial cell-derived cytokines: More than just signaling the alarm. J. Clin. Investig. 2019, 129, 1441–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sin Singer Brugiolo, A.; Carvalho Gouveia, A.C.; de Souza Alves, C.C.; de Castro E Silva, F.M.; Esteves de Oliveira, É.; Ferreira, A.P. Ferulic acid supresses Th2 immune response and prevents remodeling in ovalbumin-induced pulmonary allergy associated with inhibit ion of epithelial-derived cytokines. Pulm. Pharmacol. Ther. 2017, 45, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Saenz, S.A.; Taylor, B.C.; Artis, D. Welcome to the neighborhood: Epithelial cell-derived cytokines license innate and adaptive immune responses at mucosal sites. Immunol. Rev. 2008, 226, 172–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwakura, Y.; Ishigame, H.; Saijo, S.; Nakae, S. Functional specialization of interleukin-17 family members. Immunity 2011, 34, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Pei, C.; Zhao, C.; Wang, A.J.; Fan, A.X.; Grinchuk, V.; Smith, A.; Sun, R.; Xie, Y.; Lu, N.; Urban, J.F.; et al. Critical Role for Interleukin-25 in Host Protective Th2 Memory Response against Heligmosomoides polygyrus baker. Infect. Immun. 2016, 84, 3328–3337. [Google Scholar] [CrossRef] [Green Version]
- Mirchandani, A.S.; Salmond, R.J.; Liew, F.Y. Interleukin-33 and the function of innate lymphoid cells. Trends. Immunol. 2012, 33, 389–396. [Google Scholar] [CrossRef]
- Hardman, C.; Ogg, G. Interleukin-33, friend and foe in type-2 immune responses. Curr. Opin. Immunol. 2016, 42, 16–24. [Google Scholar] [CrossRef]
- Ziegler, S.F. The role of thymic stromal lymphopoietin (TSLP) in allergic disorders. Curr. Opin. Immunol. 2010, 22, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Kouzaki, H.; Tojima, I.; Kita, H.; Shimizu, T. Transcription of interleukin-25 and extracellular release of the protein is regulated by allergen protease in airway epithelial cells. Am. J. Resp. Cell. Mol. Biol. 2013, 49, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Uchida, M.; Anderson, E.L.; Squillace, D.L.; Patil, N.; Maniak, P.J.; Iijima, K.; Kita, H.; O’Grady, S.M. Oxidative stress serves as a key checkpoint for IL-33 release by airway epithelium. Allergy 2017, 72, 1521–1531. [Google Scholar] [CrossRef]
- Morita, H.; Arae, K.; Unno, H.; Miyauchi, K.; Toyama, S.; Nambu, A.; Oboki, K.; Ohno, T.; Motomura, K.; Matsuda, A.; et al. An interleukin-33-mast cell-interleukin-2 axis suppresses papain-induced allergic inflammation by promoting regulatory T cell numbers. Immunity 2015, 43, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouzaki, H.; Iijima, k.; Kobayashi, T.; O’Grady, S.M.; Kita, H. The danger signal, extracellular ATP, is a sensor for an airborne allergen and triggers IL-33 release and innate Th2-type responses. J. Immunol. 2011, 186, 4375–4387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Wei, Y.; Zhang, Y.-N.; Luo, X.; Yang, B.-Y.; Yu, S.-F.; Wu, X.-M.; Wu, C.-Y.; Li, H.-B. Increased IL-21 expression in chronic rhinosinusitis with nasalpolyps. Clin. Exp. Allergy 2015, 45, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Nagata, Y.; Maruoka, S.; Gon, Y.; Mizumura, K.; Kishi, H.; Nomura, Y.; Hikichi, M.; Hashimoto, S.; Oshima, T. Expression of IL-25, IL-33, and thymic stromal lymphopoietin in nasal polyp gland duct epithelium in patients with chronic rhinosinusitis. Am. J. Rhinol. Allergy 2019, 33, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Stevens, W.W.; Peters, A.T.; Tan, B.K.; Klingler, A.I.; Poposki, J.A.; Hulse, K.E.; Grammer, L.C.; Welch, K.C.; Smith, S.S.; Conley, D.B.; et al. Associations Between Inflammatory Endotypes and Clinical Presentations in Chronic Rhinosinusitis. J. Allergy Clin. Immunol. Pr. 2019, 7, 2812–2820. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; Bailey, M.; Zaunders, J.J.; Mrad, N.; Sacks, R.; Sewell, W.; Harvey, R.J. Group 2 innate lymphoid cells (ILC2s) are increased in chronic rhinosinusitis with nasal polyps or eosinophilia. Clin. Exp. Allergy 2015, 45, 394–403. [Google Scholar] [CrossRef]
- Morita, H.; Moro, K.; Koyasu, S. Innate lymphoid cells in allergic and nonallergic inflammation. J. Allergy Clin. Immunol. 2016, 138, 1253–1264. [Google Scholar] [CrossRef] [Green Version]
- Dogan, M.; Sahin, M.; Yenisey, C. Increased TSLP, IL-33, IL-25, IL-19, IL-21 and amphiregulin (AREG) levels in chronic rhinosinusitis with nasal polyps. Eur. Arch. Otorhinolaryngol. 2019, 276, 1685–1691. [Google Scholar] [CrossRef]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2020. Rhinol. Suppl. 2020, 58, 1–464. [Google Scholar] [CrossRef]
- Shin, H.W.; Kim, D.K.; Park, M.H.; Eun, K.M.; Lee, M.; So, D.; Kong, I.G.; Mo, J.H.; Yang, M.S.; Jin, H.R.; et al. IL-25 as a novel therapeutic target in nasal polyps of patients with chronic rhinosinusitis. J. Allergy Clin. Immunol. 2015, 135, 1476–1485. [Google Scholar] [CrossRef] [Green Version]
- Dolgachev, V.; Petersen, B.C.; Budelsky, A.L.; Berlin, A.A.; Lukacs, N.W. Pulmonary IL-17E (IL-25) production and IL-17RB1 myeloid cell-derived Th2 cytokine production are dependent upon stem cell factor-induced responses during chronic allergic pulmonary disease. J. Immunol. 2009, 183, 5705–5715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, Y.H.; Choi, J.K.; Choi, Y.A.; Ryoo, Z.Y.; Lee, H.S.; Park, P.H.; Kim, S.U.; Kwon, T.K.; Jang, M.H.; Im, S.H.; et al. House dust mite increases pro-Th2 cytokines IL-25 and IL-33 via the activation of TLR1/6 signaling. J. Invest. Dermatol. 2017, 139, 2354–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.H.; Yoo, J.Y.; Kim, M.J.; Hwang, S.G.; Ahn, K.C.; Ryu, J.C.; Choi, M.K.; Joo, J.H.; Kim, C.H.; Lee, S.N.; et al. Distinct TLR-mediated pathways regulate house dust mite-induced allergic disease in the upper and lower airways. J. Allergy Clin. Immunol. 2013, 131, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, A.R.; Shokri, H.; Hassan Al-Heidary, S.; Ghafarifar, F. Evaluation of murine lung epithelial cells (TC-1 JHU-1) line to develop Th2-promoting cytokines IL-25/IL-33/TSLP and genes Tlr2/Tlr4 in response to Aspergillus fumigatus. J. Mycol. Med. 2018, 28, 349–354. [Google Scholar] [CrossRef]
- Shin, S.H.; Lee, Y.H. Airborne fungi induce nasal polyp epithelial cell activation and toll-like receptor expression. Int. Arch. Allergy Immunol. 2010, 153, 46–52. [Google Scholar] [CrossRef]
- Zhong, H.; Fan, X.L.; Yu, Q.N.; Chen, D.; Xu, R.; Chen, D.H.; Lin, Z.B.; Wen, W.; Fu, Q.L. Increased innate type 2 immune response in house dust mite-allergic patients with allergic rhinitis. Clin. Immunol. 2017, 183, 293–299. [Google Scholar] [CrossRef]
- Satitsuksanoa, P.; Kennedy, M.; Gilis, D.; Le Mignon, M.; Suratannor, N.; Soh, W.T.; Wonqpiyabovom, J.; Chatchatee, P.; Vanqveravong, M.; Rerkpattanapipat, T.; et al. The minor house dust mite allergen Der p 13 is a fatty acid-binding protein and an activator of a TLR2-mediated innate immune response. Allergy 2016, 71, 1425–1434. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ENP (n = 14) | NENP (n = 16) | p Value | |
|---|---|---|---|
| Atopy | 8 (57.1%) | 6 (37.5%) | 0.021 |
| PB eosinophilia (>5%) | 7 (50.0%) | 2 (12.5%) | <0.001 |
| OFT | <0.001 | ||
| Normosmia | 2 | 5 | |
| Hyposmia | 3 | 7 | |
| Anosmia | 9 | 4 | |
| Bilaterality | 13 (92.9%) | 11 (68.8%) | 0.026 |
| LM score | 7.9 ± 2.3 | 7.2 ± 2.4 | 0.137 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.-H.; Ye, M.-K.; Lee, D.-W.; Chae, M.-H.; Han, B.-D. Nasal Epithelial Cells Activated with Alternaria and House Dust Mite Induce Not Only Th2 but Also Th1 Immune Responses. Int. J. Mol. Sci. 2020, 21, 2693. https://doi.org/10.3390/ijms21082693
Shin S-H, Ye M-K, Lee D-W, Chae M-H, Han B-D. Nasal Epithelial Cells Activated with Alternaria and House Dust Mite Induce Not Only Th2 but Also Th1 Immune Responses. International Journal of Molecular Sciences. 2020; 21(8):2693. https://doi.org/10.3390/ijms21082693
Chicago/Turabian StyleShin, Seung-Heon, Mi-Kyung Ye, Dong-Won Lee, Mi-Hyun Chae, and Ba-Da Han. 2020. "Nasal Epithelial Cells Activated with Alternaria and House Dust Mite Induce Not Only Th2 but Also Th1 Immune Responses" International Journal of Molecular Sciences 21, no. 8: 2693. https://doi.org/10.3390/ijms21082693
APA StyleShin, S.-H., Ye, M.-K., Lee, D.-W., Chae, M.-H., & Han, B.-D. (2020). Nasal Epithelial Cells Activated with Alternaria and House Dust Mite Induce Not Only Th2 but Also Th1 Immune Responses. International Journal of Molecular Sciences, 21(8), 2693. https://doi.org/10.3390/ijms21082693