The Importance of Porins and ?-Lactamase in Outer Membrane Vesicles on the Hydrolysis of ?-Lactam Antibiotics


 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Characterization of Mutant Strains
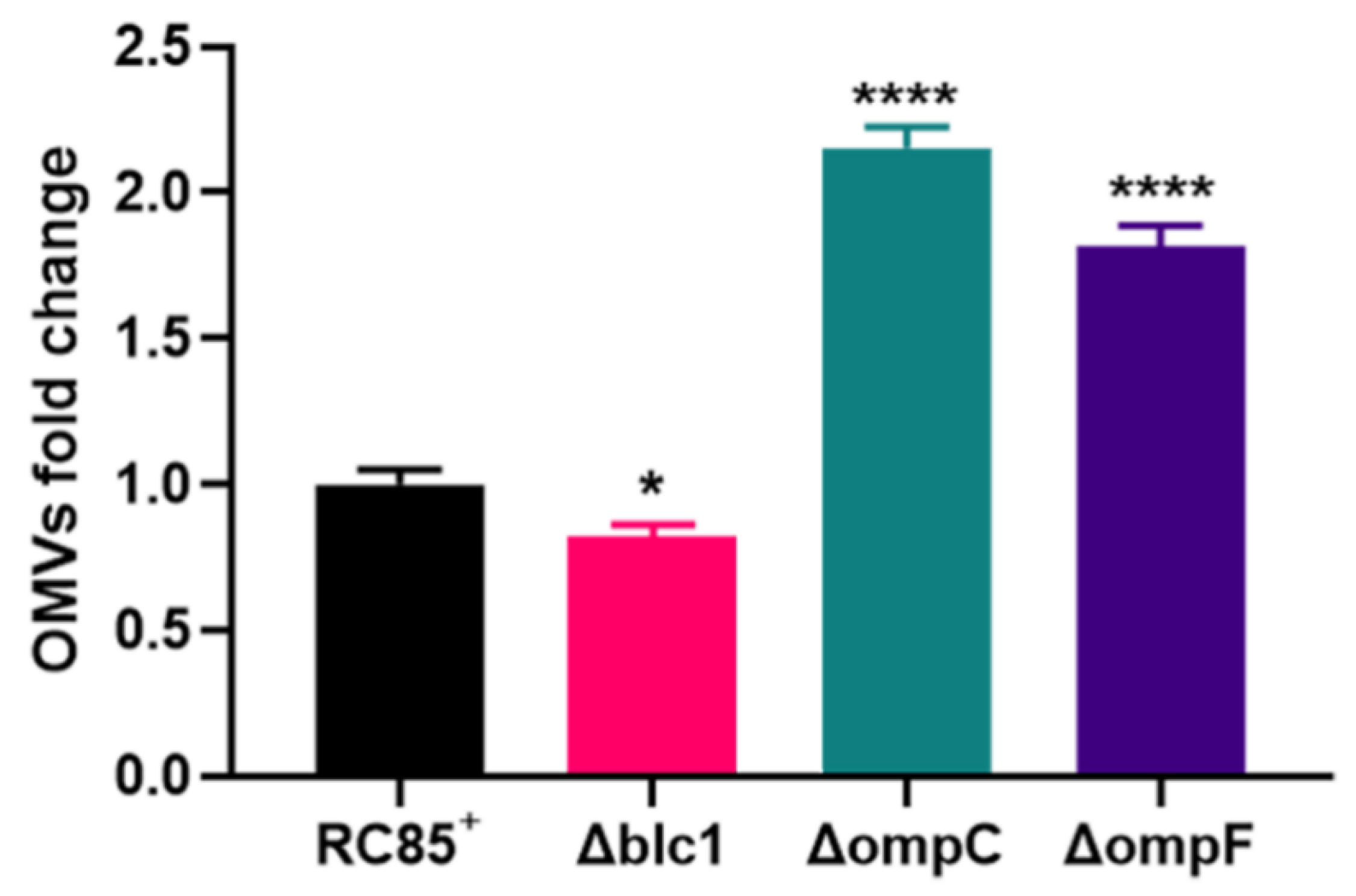
2.2. Quantification of the Produced OMVs
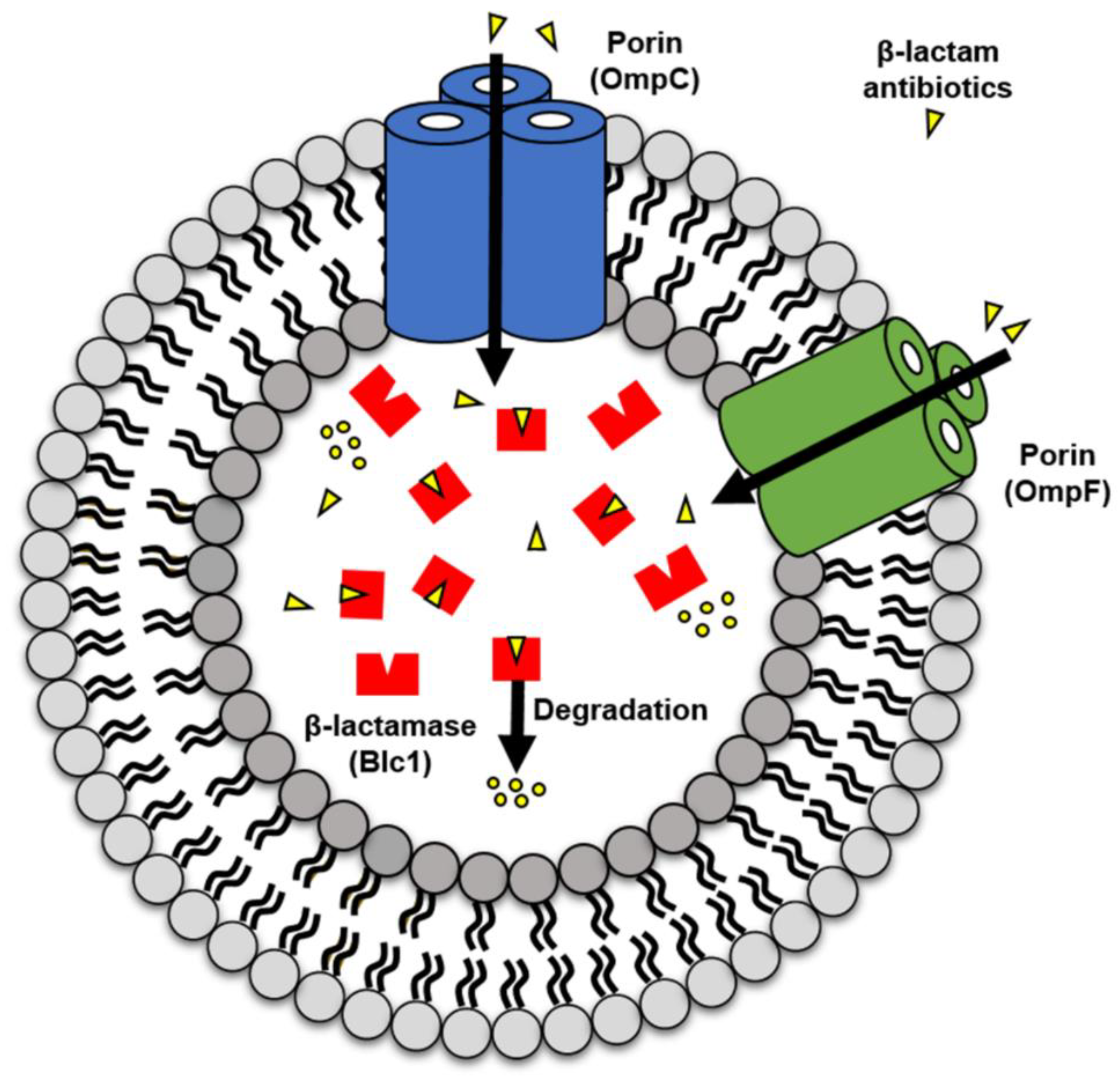
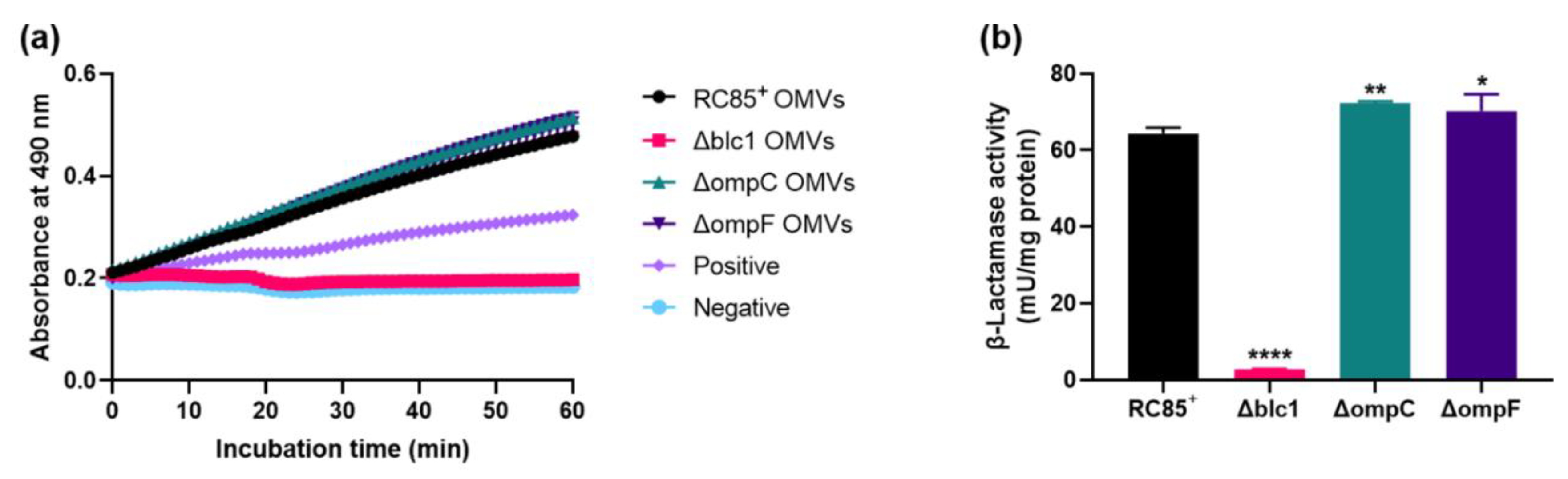
2.3. Comparison of β-Lactamase Activity
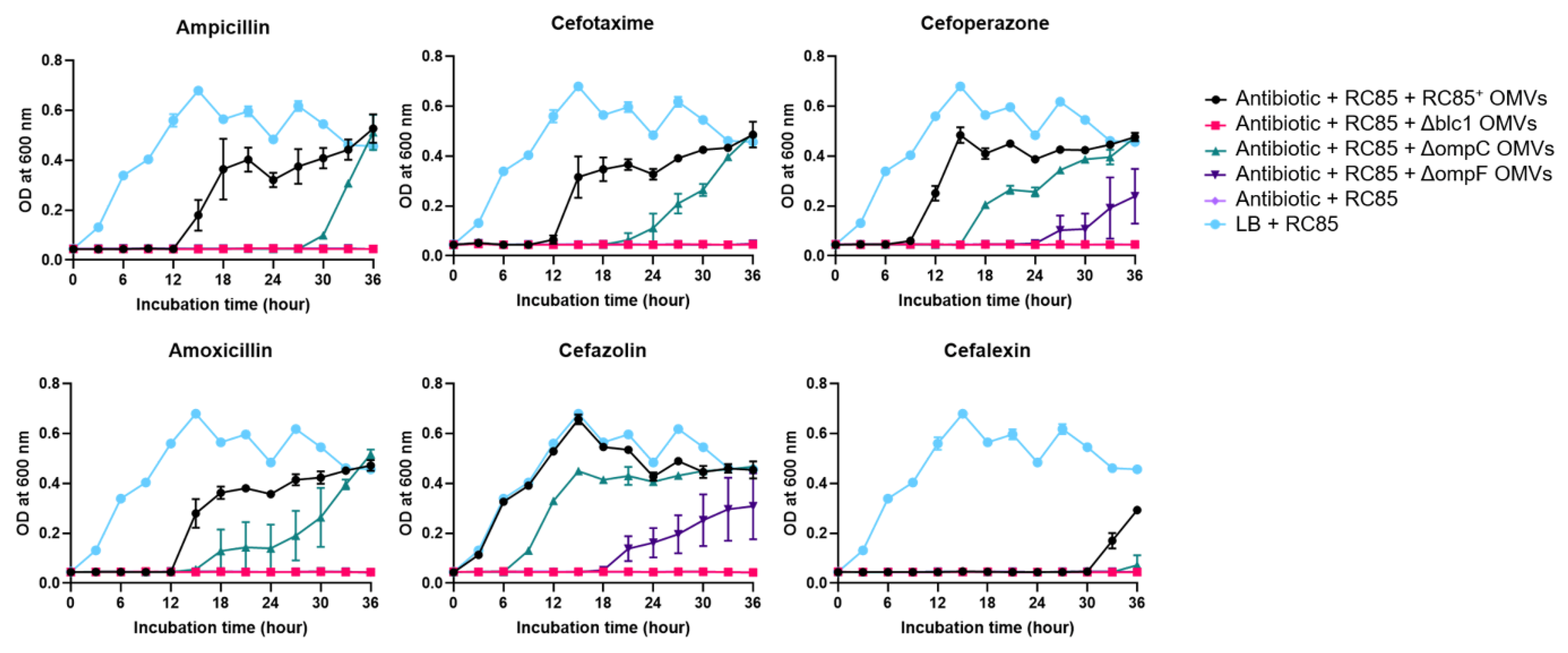
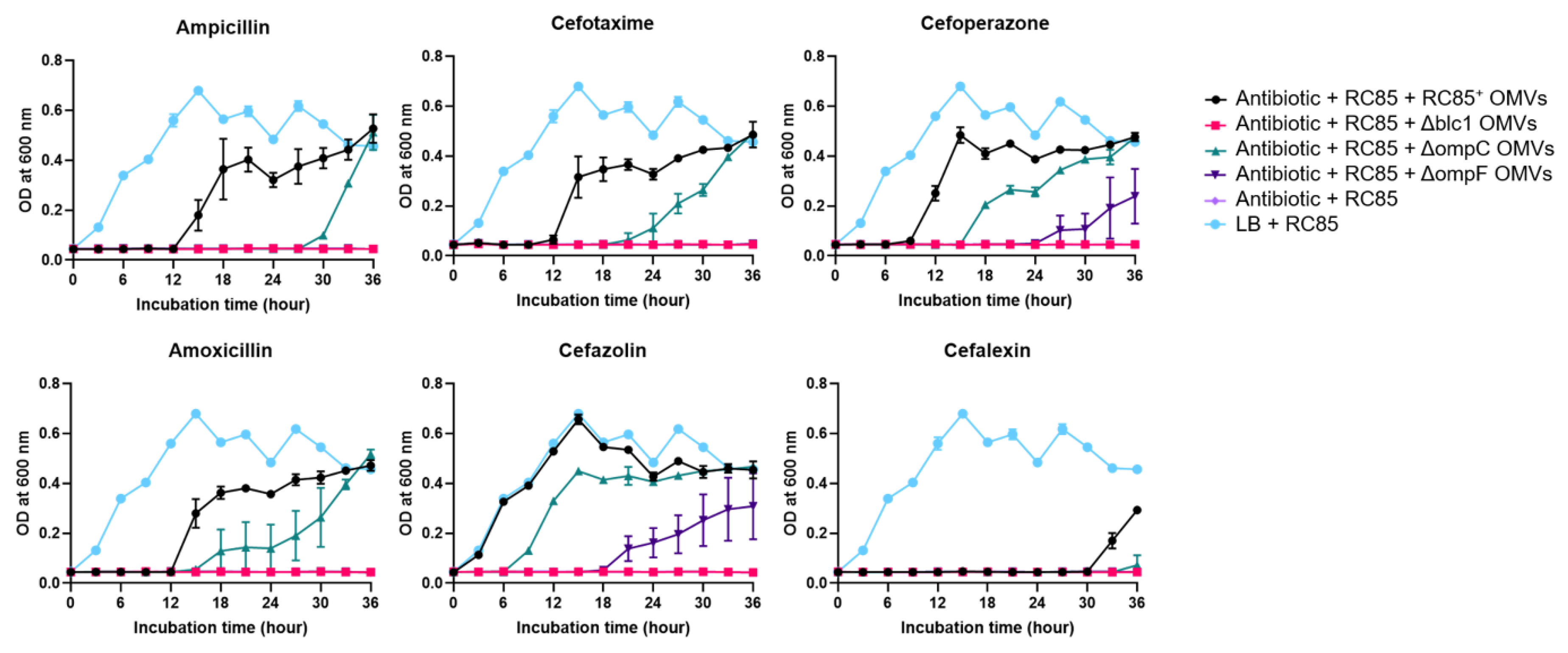
2.4. Evaluation of the Protective Role of OMVs against β-Lactam Antibiotics
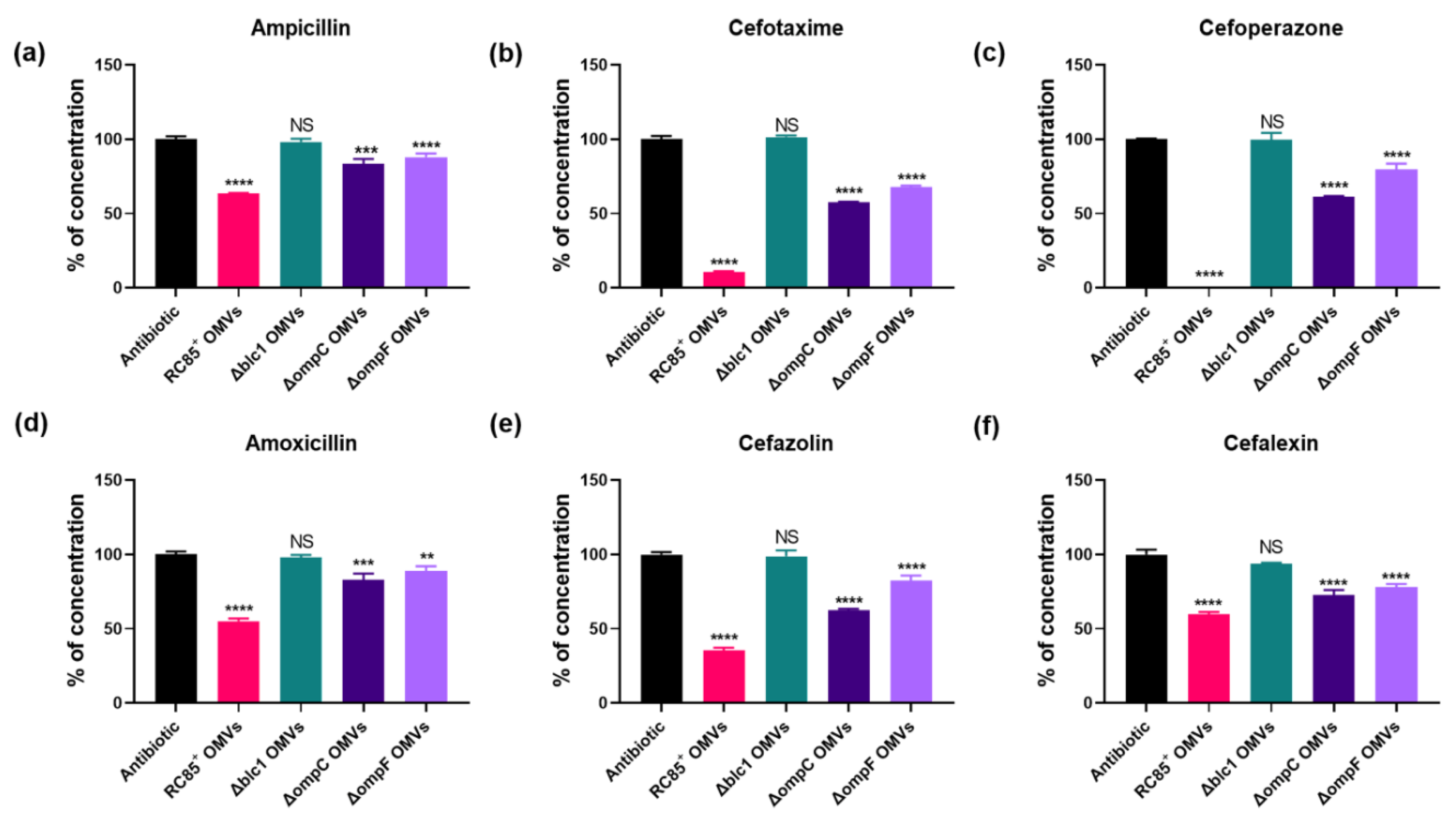
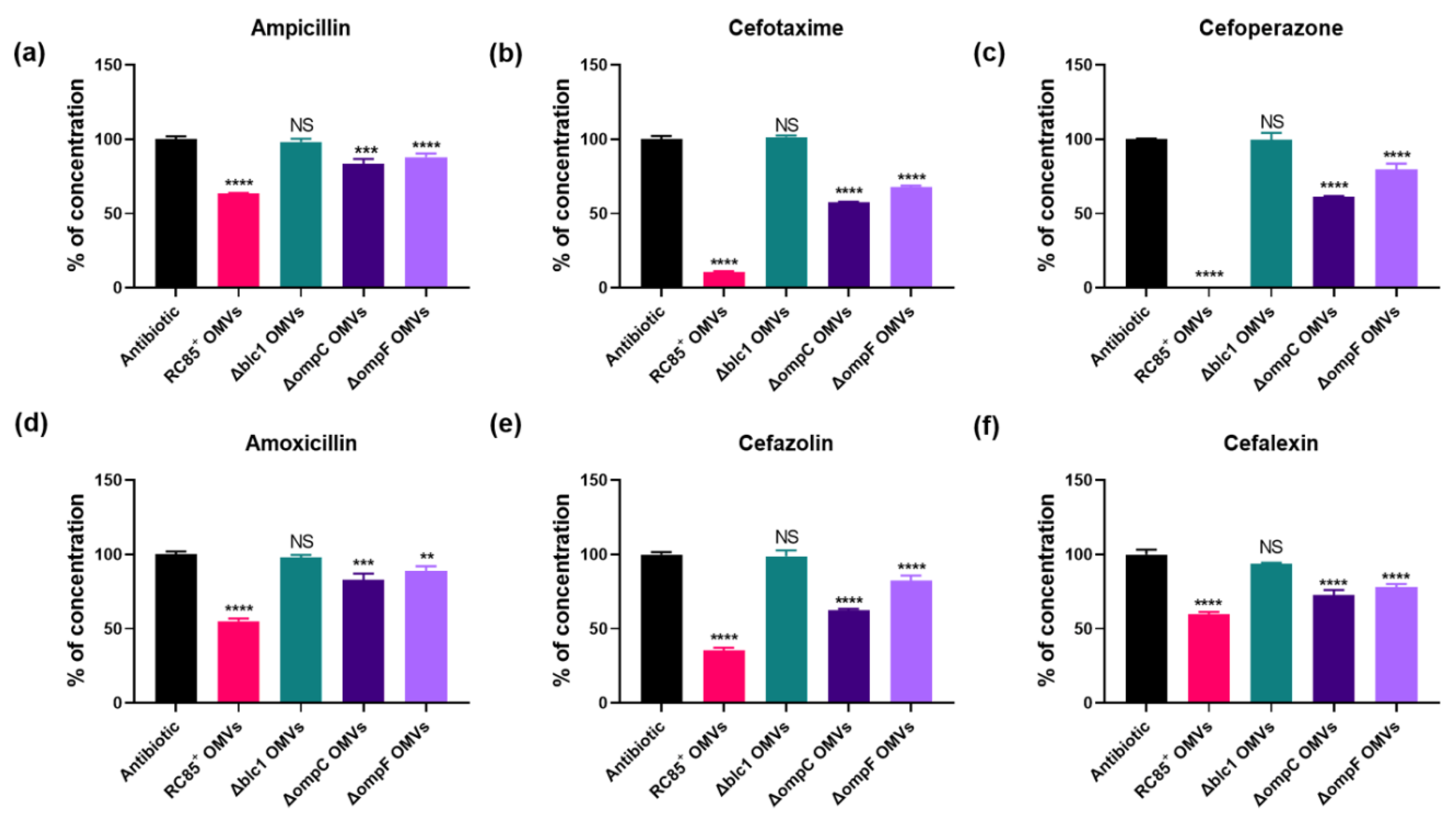
2.5. Hydrolysis of β-Lactam Antibiotics by OMVs
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Molecular Cloning and Mutant Construction
4.3. Analysis of Antibiotic Resistance
4.4. Isolation of Pure OMVs
4.5. Effect of OMVs on the Growth of Bacteria in the Presence of β-Lactam Antibiotics
4.6. Quantification of β-Lactamase Activity
4.7. Measurement of Antibiotic Concentrations
4.8. Statistical Analysis
4.9. Data Availability
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nazir, T.; Abraham, S.; Islam, A. Emergence of potential superbug Mycobacterium tuberculosis, lessons from New Delhi mutant-1 bacterial strains. Int. J. Health Sci. 2012, 6, 87. [Google Scholar] [CrossRef]
- Singh, S. Irrational use of antimicrobial agents. JDR Clin. Trans. Res. 2018, 2, 1–2. [Google Scholar] [CrossRef]
- Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. Review on Antimicrobial Resistance: Chaired by Jim O’Neill. 2014. Available online: http://amr-review.org/Publications (accessed on 1 September 2015).
- Centers for Disease Control and Prevention. Antibiotic resistance threats in the United States, 2013; Centres for Disease Control and Prevention: Atlanta, GA, USA, 2013.
- Antimicrobial Resistance: Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Chaired by Jim O’Neill. 2016. Available online: http://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf (accessed on 27 March 2017).
- Lammie, S.L.; Hughes, J.M. Antimicrobial resistance, food safety, and one health: The need for convergence. Annu. Rev. Food Sci. Technol. 2016, 7, 287–312. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Park, S.B.; Im, S.P.; Lee, J.S.; Jung, J.W.; Gong, T.W.; Lazarte, J.M.S.; Kim, J.; Seo, J.-S.; Kim, J.-H. Outer membrane vesicles from β-lactam-resistant Escherichia coli enable the survival of β-lactam-susceptible E. coli in the presence of β-lactam antibiotics. Sci. Rep. 2018, 8, 5402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beveridge, T.J. Structures of gram-negative cell walls and their derived membrane vesicles. J. Bacteriol. 1999, 181, 4725–4733. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, H.M.; Nagaraj, R.; Jagannadham, M.V. Protective role of E. coli outer membrane vesicles against antibiotics. Microbiol. Res. 2015, 181, 1–7. [Google Scholar] [CrossRef]
- Turner, K.L.; Cahill, B.K.; Dilello, S.K.; Gutel, D.; Brunson, D.N.; Albertí, S.; Ellis, T.N. Porin loss impacts the host inflammatory response to outer membrane vesicles of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2016, 60, 1360–1369. [Google Scholar] [CrossRef] [Green Version]
- Schooling, S.R.; Beveridge, T.J. Membrane vesicles: An overlooked component of the matrices of biofilms. J. Bacteriol. 2006, 188, 5945–5957. [Google Scholar] [CrossRef] [Green Version]
- Kulp, A.; Kuehn, M.J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef] [Green Version]
- Yaron, S.; Kolling, G.L.; Simon, L.; Matthews, K.R. Vesicle-mediated transfer of virulence genes from Escherichia coli O157: H7 to other enteric bacteria. Appl. Environ. Microbiol. 2000, 66, 4414–4420. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.-O.; Gho, Y.S.; Lee, J.C.; Kim, S.I. Proteome analysis of outer membrane vesicles from a clinical Acinetobacter baumannii isolate. FEMS Microbiol. Lett. 2009, 297, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Mashburn, L.M.; Whiteley, M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature 2005, 437, 422. [Google Scholar] [CrossRef] [PubMed]
- Schertzer, J.W.; Whiteley, M. Bacterial outer membrane vesicles in trafficking, communication and the host-pathogen interaction. J. Mol. Microbiol. Biotechnol. 2013, 23, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Deatherage, B.L.; Cookson, B.T. Membrane vesicle release in bacteria, eukaryotes, and archaea: A conserved yet underappreciated aspect of microbial life. Infect. Immun. 2012, 80, 1948–1957. [Google Scholar] [CrossRef] [Green Version]
- Rumbo, C.; Fernández-Moreira, E.; Merino, M.; Poza, M.; Mendez, J.A.; Soares, N.C.; Mosquera, A.; Chaves, F.; Bou, G. Horizontal transfer of the OXA-24 carbapenemase gene via outer membrane vesicles: A new mechanism of dissemination of carbapenem resistance genes in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 3084–3090. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, E.-Y.; Kim, S.-H.; Kim, D.-K.; Park, K.-S.; Kim, K.P.; Kim, Y.-K.; Roh, T.-Y.; Gho, Y.S. Staphylococcus aureus extracellular vesicles carry biologically active β-lactamase. Antimicrob. Agents Chemother. 2013, 57, 2589–2595. [Google Scholar] [CrossRef] [Green Version]
- Schaar, V.; Nordström, T.; Mörgelin, M.; Riesbeck, K. Moraxella catarrhalis outer membrane vesicles carry β-lactamase and promote survival of Streptococcus pneumoniae and Haemophilus influenzae by inactivating amoxicillin. Antimicrob. Agents Chemother. 2011, 55, 3845–3853. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, H.M.; Swamy, C.V.; Jagannadham, M.V. Molecular characterization and functional analysis of outer membrane vesicles from the antarctic bacterium Pseudomonas syringae suggest a possible response to environmental conditions. J. Proteome Res. 2014, 13, 1345–1358. [Google Scholar] [CrossRef]
- Doumith, M.; Ellington, M.J.; Livermore, D.M.; Woodford, N. Molecular mechanisms disrupting porin expression in ertapenem-resistant Klebsiella and Enterobacter spp. clinical isolates from the UK. J. Antimicrob. Chemother. 2009, 63, 659–667. [Google Scholar] [CrossRef]
- Bavoil, P.; Nikaido, H.; von Meyenburg, K. Pleiotropic transport mutants of Escherichia coli lack porin, a major outer membrane protein. Mol. Gen. Genet. 1977, 158, 23–33. [Google Scholar] [CrossRef]
- Nikaido, H.; Song, S.A.; Shaltiel, L.; Nurminen, M. Outer membrane of Salmonella XIV. Reduced transmembrane diffusion rates in porin-deficient mutants. Biochem. Biophys. Res. Commun. 1977, 76, 324–330. [Google Scholar] [CrossRef]
- Pagès, J.-M.; James, C.E.; Winterhalter, M. The porin and the permeating antibiotic: A selective diffusion barrier in Gram-negative bacteria. Nat. Rev. Microbiol. 2008, 6, 893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamers, R.P.; Nguyen, U.T.; Nguyen, Y.; Buensuceso, R.N.; Burrows, L.L. Loss of membrane-bound lytic transglycosylases increases outer membrane permeability and β-lactam sensitivity in Pseudomonas aeruginosa. Microbiologyopen 2015, 4, 879–895. [Google Scholar] [CrossRef] [PubMed]
- Schaar, V.; Paulsson, M.; Mörgelin, M.; Riesbeck, K. Outer membrane vesicles shield Moraxella catarrhalis β-lactamase from neutralization by serum IgG. J. Antimicrob. Chemother. 2012, 68, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Fulsundar, S.; Harms, K.; Flaten, G.E.; Johnsen, P.J.; Chopade, B.A.; Nielsen, K.M. Gene transfer potential of outer membrane vesicles of Acinetobacter baylyi and effects of stress on vesiculation. Appl. Environ. Microbiol. 2014, 80, 3469–3483. [Google Scholar] [CrossRef] [Green Version]
- Renelli, M.; Matias, V.; Lo, R.Y.; Beveridge, T.J. DNA-containing membrane vesicles of Pseudomonas aeruginosa PAO1 and their genetic transformation potential. Microbiology 2004, 150, 2161–2169. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605. [Google Scholar] [CrossRef] [Green Version]
- Manning, A.J.; Kuehn, M.J. Contribution of bacterial outer membrane vesicles to innate bacterial defense. BMC Microbiol. 2011, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Andreoni, F.; Toyofuku, M.; Menzi, C.; Kalawong, R.; Shambat, S.M.; François, P.; Zinkernagel, A.S.; Eberl, L. Antibiotics stimulate formation of vesicles in Staphylococcus aureus in both phage-dependent and-independent fashions and via different routes. Antimicrob. Agents Chemother. 2019, 63, e01439-18. [Google Scholar] [CrossRef] [Green Version]
- Quan, K.; Zhu, Z.; Cao, S.; Zhang, F.; Miao, C.; Wen, X.; Huang, X.; Wen, Y.; Wu, R.; Yan, Q. Escherichia coli-derived outer membrane vesicles deliver galactose-1-phosphate uridyltransferase and yield partial protection against Actinobacillus pleuropneumoniae in mice. J. Microbiol. Biotechnol. 2018, 28, 2095–2105. [Google Scholar] [CrossRef] [Green Version]
- McBroom, A.J.; Johnson, A.P.; Vemulapalli, S.; Kuehn, M.J. Outer membrane vesicle production by Escherichia coli is independent of membrane instability. J. Bacteriol. 2006, 188, 5385–5392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwechheimer, C.; Kulp, A.; Kuehn, M.J. Modulation of bacterial outer membrane vesicle production by envelope structure and content. BMC Microbiol. 2014, 14, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valeru, S.P.; Shanan, S.; Alossimi, H.; Saeed, A.; Sandström, G.; Abd, H. Lack of outer membrane protein A enhances the release of outer membrane vesicles and survival of Vibrio cholerae and suppresses viability of Acanthamoeba castellanii. Int. J. Microbiol. 2014, 2014, 610190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogami, T.; Mizushima, S. Outer membrane porins are important in maintenance of the surface structure of Escherichia coli cells. J. Bacteriol. 1983, 156, 402–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holten, K.B. Appropriate prescribing of oral beta-lactam antibiotics. Am. Fam. Physician 2000, 62, 611–620. [Google Scholar] [PubMed]
- Jaffe, A.; Chabbert, Y.A.; Semonin, O. Role of porin proteins OmpF and OmpC in the permeation of beta-lactams. Antimicrob. Agents Chemother. 1982, 22, 942–948. [Google Scholar] [CrossRef] [Green Version]
- Drawz, S.M.; Bonomo, R.A. Three decades of β-lactamase inhibitors. Clin. Microbiol. Rev. 2010, 23, 160–201. [Google Scholar] [CrossRef] [Green Version]
- Davies, J. Inactivation of antibiotics and the dissemination of resistance genes. Science 1994, 264, 375–382. [Google Scholar] [CrossRef]
- Nikaido, H.; Nakae, T. The outer membrane of Gram-negative bacteria. In Advances in Microbial Physiology; Elsevier: Amsterdam, The Netherlands, 1980; Volume 20, pp. 163–250. [Google Scholar]
- Nakae, T. Outer membrane of Salmonella. Isolation of protein complex that produces transmembrane channels. J. Biol. Chem. 1976, 251, 2176–2178. [Google Scholar]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Minasov, G.; Shoichet, B.K. Evolution of an antibiotic resistance enzyme constrained by stability and activity trade-offs. J. Mol. Biol. 2002, 320, 85–95. [Google Scholar] [CrossRef]
- Hwang, J.; Cho, K.-H.; Song, H.; Yi, H.; Kim, H.S. Deletion mutations conferring substrate spectrum extension in the class A β-lactamase. Antimicrob. Agents Chemother. 2014, 58, 6265–6269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, U.; Lee, C.-R. Distinct roles of outer membrane porins in antibiotic resistance and membrane integrity in Escherichia coli. Front. Microbiol. 2019, 10, 953. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Rosenberg, E.; Foulds, J. Porin channels in Escherichia coli: Studies with beta-lactams in intact cells. J. Bacteriol. 1983, 153, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Lou, H.; Chen, M.; Black, S.S.; Bushell, S.R.; Ceccarelli, M.; Mach, T.; Beis, K.; Low, A.S.; Bamford, V.A.; Booth, I.R. Altered antibiotic transport in OmpC mutants isolated from a series of clinical strains of multi-drug resistant E. coli. PLoS ONE 2011, 6, e25825. [Google Scholar] [CrossRef] [Green Version]
- Kojima, S.; Nikaido, H. High salt concentrations increase permeability through OmpC channels of Escherichia coli. J. Biol. Chem. 2014, 289, 26464–26473. [Google Scholar] [CrossRef] [Green Version]
- Moya-Torres, A.; Mulvey, M.R.; Kumar, A.; Oresnik, I.J.; Brassinga, A.K.C. The lack of OmpF, but not OmpC, contributes to increased antibiotic resistance in Serratia marcescens. Microbiology 2014, 160, 1882–1892. [Google Scholar] [CrossRef]
- Dutzler, R.; Rummel, G.; Albertí, S.; Hernández-Allés, S.; Phale, P.; Rosenbusch, J.; Benedi, V.; Schirmer, T. Crystal structure and functional characterization of OmpK36, the osmoporin of Klebsiella pneumoniae. Structure 1999, 7, 425–434. [Google Scholar] [CrossRef]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim Biophys Acta Proteins Proteom 2009, 1794, 808–816. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.Y.; Bang, J.Y.; Park, G.W.; Choi, D.S.; Kang, J.S.; Kim, H.J.; Park, K.S.; Lee, J.O.; Kim, Y.K.; Kwon, K.H. Global proteomic profiling of native outer membrane vesicles derived from Escherichia coli. Proteomics 2007, 7, 3143–3153. [Google Scholar] [CrossRef]
- Yoshimura, F.; Nikaido, H. Diffusion of beta-lactam antibiotics through the porin channels of Escherichia coli K-12. Antimicrob. Agents Chemother. 1985, 27, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, T.J.; Snow, G.A. Biochemistry and Molecular Biology of Antimicrobial Drug Action; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Kim, J.; Im, S.P.; Lee, J.S.; Lazarte, J.M.S.; Kim, S.W.; Jung, J.W.; Kim, J.Y.; Kim, Y.R.; Lee, S.; Kim, G.J. Globular-shaped variable lymphocyte receptors B antibody multimerized by a hydrophobic clustering in hagfish. Sci. Rep. 2018, 8, 10801. [Google Scholar] [CrossRef] [PubMed]
- Heermann, R.; Zeppenfeld, T.; Jung, K. Simple generation of site-directed point mutations in the Escherichia coli chromosome using Red®/ET® Recombination. Microb. Cell Fact. 2008, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Jang, H.B.; Lee, J.S.; Im, S.P.; Lazarte, J.M.S.; Seo, J.P.; Lee, W.J.; Kim, J.S.; Jung, T.S. Comparison of proteome typing and serotyping of Streptococcus parauberis isolates from olive flounder (Paralichthys olivaceus). J. Microbiol. Methods 2015, 118, 168–172. [Google Scholar] [CrossRef]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48 (Suppl. 1), 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokri, A.; Larsson, G. Characterisation of the Escherichia coli membrane structure and function during fedbatch cultivation. Microb. Cell Fact. 2004, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Bush, K.; Singer, S.B. Effective cooling allows sonication to be used for liberation of β-lactamases from Gramnegative bacteria. J. Antimicrob. Chemother. 1989, 24, 82–84. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Antibiotics | MIC (μg/mL)a | |||
|---|---|---|---|---|---|
| RC85+ | Δblc1 | ΔompC | ΔompF | ||
| β-lactam antibiotics | Ampicillin | >1024 | 4 | >1024 | >1024 |
| Cefotaxime | >1024 | <1/2 | >1024 | >1024 | |
| Cefoperazone | 1024 | <1/2 | 1024 | >1024 | |
| Methicillin | >1024 | 256 | >1024 | >1024 | |
| Amoxicillin | >1024 | 2 | >1024 | >1024 | |
| Cefazolin | 1024 | 1 | >1024 | >1024 | |
| Cefalexin | 512 | 8 | 512 | 1024 | |
| Cloxacillin | >1024 | 128 | >1024 | >1024 | |
| Other class antibiotics | Streptomycin | >1024 | >1024 | >1024 | >1024 |
| Kanamycin | >1024 | >1024 | >1024 | >1024 | |
| Colistin | 4 | 4 | 4 | 4 | |
| Amikacin | 8 | 8 | 8 | 8 | |
| Nalidixic acid | >1024 | >1024 | >1024 | >1024 | |
| Primer | Oligonucleotide Sequence (5′ to 3′) | Target Gene | Fragment Size (bp) |
|---|---|---|---|
| blc1-F | CTGGGTGTGGCATTGATTAAC | blc1 | 374 |
| blc1-R | TAACGTCGGCTCGGTACG | ||
| ompC-F | ATGAAAGTTAAAGTACTGTCCCTC | ompC | 1103 |
| ompC-R | TTAGAACTGGTAAACCAGACCC | ||
| ompF-F | CTGACCGGTTATGGTCAGTG | ompF | 599 |
| ompF-R | CGTTTTGTTGGCGAAGCC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.W.; Lee, J.S.; Park, S.B.; Lee, A.R.; Jung, J.W.; Chun, J.H.; Lazarte, J.M.S.; Kim, J.; Seo, J.-S.; Kim, J.-H.; et al. The Importance of Porins and ?-Lactamase in Outer Membrane Vesicles on the Hydrolysis of ?-Lactam Antibiotics. Int. J. Mol. Sci. 2020, 21, 2822. https://doi.org/10.3390/ijms21082822
Kim SW, Lee JS, Park SB, Lee AR, Jung JW, Chun JH, Lazarte JMS, Kim J, Seo J-S, Kim J-H, et al. The Importance of Porins and ?-Lactamase in Outer Membrane Vesicles on the Hydrolysis of ?-Lactam Antibiotics. International Journal of Molecular Sciences. 2020; 21(8):2822. https://doi.org/10.3390/ijms21082822
Chicago/Turabian StyleKim, Si Won, Jung Seok Lee, Seong Bin Park, Ae Rin Lee, Jae Wook Jung, Jin Hong Chun, Jassy Mary S. Lazarte, Jaesung Kim, Jong-Su Seo, Jong-Hwan Kim, and et al. 2020. "The Importance of Porins and ?-Lactamase in Outer Membrane Vesicles on the Hydrolysis of ?-Lactam Antibiotics" International Journal of Molecular Sciences 21, no. 8: 2822. https://doi.org/10.3390/ijms21082822
APA StyleKim, S. W., Lee, J. S., Park, S. B., Lee, A. R., Jung, J. W., Chun, J. H., Lazarte, J. M. S., Kim, J., Seo, J.-S., Kim, J.-H., Song, J.-W., Ha, M. W., Thompson, K. D., Lee, C.-R., Jung, M., & Jung, T. S. (2020). The Importance of Porins and ?-Lactamase in Outer Membrane Vesicles on the Hydrolysis of ?-Lactam Antibiotics. International Journal of Molecular Sciences, 21(8), 2822. https://doi.org/10.3390/ijms21082822