Changes in TRPV1-Mediated Physiological Function in Rats Systemically Treated With Capsaicin on the Neonate


Abstract

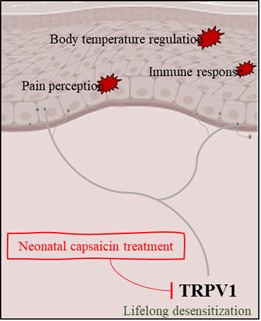
1. Introduction
2. Neonatal Capsaicin Treatment and TRPV1
3. Loss of Pain Perception
4. Changes in Body Temperature and Exposure to Infection
5. Chronic Itching
6. TRPV1 and Circadian Body Temperature Rhythm
7. Conclusions
Conflicts of Interest
Abbreviations
| Bmal1 | Brain and Muscle ARNT-Like protein 1 |
| Hsf1 | Heat shock factor 1 |
| Per1 | Period circadian regulator 1 |
| Per2 | Period circadian regulator 2 |
| TRPV1 | Transient receptor potential vanilloid 1 |
References
- Fattori, V.; Hohmann, M.S.; Rossaneis, A.C.; Ribeiro, F.P.; Verri, W.A. Capsaicin: Current Understanding of Its Mechanisms and Therapy of Pain and Other Pre-Clinical and Clinical Uses. Molecules 2016, 21, 844. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Bley, K. Topical capsaicin for pain management: Therapeutic potential and mechanisms of action of the new high-concentration capsaicin 8% patch. Br. J. Anaesth. 2011, 107, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Vlachova, V.; Lyfenko, A.; Orkand, R.K.; Vyklický, L. The effects of capsaicin and acidity on currents generated by noxious heat in cultured neonatal rat dorsal root ganglion neurones. J. Physiol. 2001, 533, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.; Sharper, V.; Hoberman, A.; Bley, K. Developmental Toxicity Study of Pure trans-Capsaicin in Rats and Rabbits. Int. J. Toxicol. 2006, 25, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Donnerer, J.; Liebmann, I.; Schicho, R. Differential Regulation of 3-Beta-Hydroxysteroid Dehydrogenase and Vanilloid Receptor TRPV1 mRNA in Sensory Neurons by Capsaicin and NGF. Pharmacology 2005, 73, 97–101. [Google Scholar] [CrossRef]
- Simone, N.A.; Collins, J.; Baumann, T.K.; LaMotte, R.H. Sensitization of cat dorsal horn neurons to innocuous mechanical stimulation after intradermal injection of capsaicin. Brain Res. 1989, 486, 185–189. [Google Scholar] [CrossRef]
- Srbely, J.; Dickey, J.P.; Bent, L.R.; Lee, D.; Lowerison, M. Capsaicin-Induced Central Sensitization Evokes Segmental Increases in Trigger Point Sensitivity in Humans. J. Pain 2010, 11, 636–643. [Google Scholar] [CrossRef]
- Holzer, P. The pharmacological challenge to tame the transient receptor potential vanilloid-1 (TRPV1) nocisensor. Br. J. Pharmacol. 2008, 155, 1145–1162. [Google Scholar] [CrossRef]
- Story, G.M. The emerging role of TRP channels in mechanisms of temperature and pain sensation. Curr. Neuropharmacol. 2006, 4, 183–196. [Google Scholar] [CrossRef]
- Jeong, K.-Y.; Seong, J. Neonatal capsaicin treatment in rats affects TRPV1-related noxious heat sensation and circadian body temperature rhythm. J. Neurol. Sci. 2014, 341, 58–63. [Google Scholar] [CrossRef]
- Newson, P.N.; Buuse, M.V.D.; Martin, S.; Lynch-Frame, A.; Chahl, L. Effects of neonatal treatment with the TRPV1 agonist, capsaicin, on adult rat brain and behaviour. Behav. Brain Res. 2014, 272, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Meller, S.T.; Gebhart, G.F.; Maves, T.J. Neonatal capsaicin treatment prevents the development of the thermal hyperalgesia produced in a model of neuropathic pain in the rat. Pain 1992, 51, 317–321. [Google Scholar] [CrossRef]
- Jeong, K.-Y.; Kim, H.M. Neonatal capsaicin treatment in rats induces chronic hyperthermia resulting in infectious disease. Exp. Ther. Med. 2015, 10, 2417–2423. [Google Scholar] [CrossRef] [PubMed]
- Bowden, J.J.; Baluk, P.; Lefevre, P.M.; Schoeb, T.R.; Lindsey, J.R.; McDonald, D.M. Sensory denervation by neonatal capsaicin treatment exacerbates Mycoplasma pulmonis infection in rat airways. Am. J. Physiol. Cell. Mol. Physiol. 1996, 270, L393–L403. [Google Scholar] [CrossRef]
- Ljungdahl, A.; Kristensson, K.; Lundberg, J.; Lycke, E.; Svennerholm, B.; Ziegler, R. Herpes simplex virus infection in capsaicin-treated mice. J. Neurol. Sci. 1986, 72, 223–230. [Google Scholar] [CrossRef]
- Back, S.K.; Jeong, K.-Y.; Li, C.; Lee, J.; Lee, S.-B.; Na, H.S. Chronically relapsing pruritic dermatitis in the rats treated as neonate with capsaicin; a potential rat model of human atopic dermatitis. J. Dermatol. Sci. 2012, 67, 111–119. [Google Scholar] [CrossRef]
- Jeong, K.-Y.; Lee, J.; Li, C.; Han, T.; Lee, S.-B.; Lee, H.; Back, S.K.; Na, H.S. Juvenile Obesity Aggravates Disease Severity in a Rat Model of Atopic Dermatitis. Allergy, Asthma Immunol. Res. 2014, 7, 69–75. [Google Scholar] [CrossRef]
- Thomas, D.A.; Dubner, R.; Ruda, M. Neonatal capsaicin treatment in rats results in scratching behavior with skin damage: Potential model of non-painful dysesthesia. Neurosci. Lett. 1994, 171, 101–104. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Sikand, P. TRPV1: A Target for Next Generation Analgesics. Curr. Neuropharmacol. 2008, 6, 151–163. [Google Scholar] [CrossRef]
- Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Sensory TRP channels: The key transducers of nociception and pain. Prog. Mol. Biol. Transl. Sci. 2015, 131, 73–118. [Google Scholar]
- Hanson, S.M.; Newstead, S.; Swartz, K.J.; Sansom, M.S.P. Capsaicin interaction with TRPV1 channels in a lipid bilayer: Molecular dynamics simulation. Biophys. J. 2015, 108, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Vyklický, L.; Nováková-Tousová, K.; Benedikt, J.; Samad, A.; Touska, F.; Vlachová, V. Calcium-dependent desensitization of vanilloid receptor TRPV1: A mechanism possibly involved in analgesia induced by topical application of capsaicin. Physiol. Res. 2018, 57 (Suppl. 3), S59–S68. [Google Scholar]
- Touska, F.; Marsakova, L.; Teisinger, J.; Vlachová, V. A “cute” desensitization of TRPV1. Curr. Pharm. Biotechnol. 2011, 12, 122–129. [Google Scholar] [CrossRef]
- Holzer, P. Capsaicin as a Tool for Studying Sensory Neuron Functions. Results and Problems in Cell Differentiation 1991, 298, 3–16. [Google Scholar] [CrossRef]
- Baamonde, A.; Lastra, A.; Juárez, L.; Hidalgo, A.; Menendez, L. TRPV1 desensitisation and endogenous vanilloid involvement in the enhanced analgesia induced by capsaicin in inflamed tissues. Brain Res. Bull. 2005, 67, 476–481. [Google Scholar] [CrossRef]
- Dougherty, P.M.; Willis, W.; Lenz, F.; Dougherty, M.P.; Willis, D.W.; Lenz, A.F. Transient inhibition of responses to thermal stimuli of spinal sensory tract neurons in monkeys during sensitization by intradermal capsaicin. Pain 1998, 77, 129–136. [Google Scholar] [CrossRef]
- Nakao, A.; Takahashi, Y.; Nagase, M.; Ikeda, R.; Kato, F. Role of capsaicin-sensitive C-fiber afferents in neuropathic pain-induced synaptic potentiation in the nociceptive amygdala. Mol. Pain 2012, 8, 51. [Google Scholar] [CrossRef]
- Nagy, I.; Sántha, P.; Jancsó, G.; Urbán, L. The role of the vanilloid (capsaicin) receptor (TRPV1) in physiology and pathology. Eur. J. Pharmacol. 2004, 500, 351–369. [Google Scholar] [CrossRef]
- Hiura, A. Neuroanatomical Effects of Capsaicin on the Primary Afferent Neurons. Arch. Histol. Cytol. 2000, 63, 199–215. [Google Scholar] [CrossRef]
- Nagy, J.I.; Van Der Kooy, D. Effects of neonatal capsaicin treatment on nociceptive thresholds in the rat. J. Neurosci. 1983, 3, 1145–1150. [Google Scholar]
- Hua, X.-Y.; Calcutt, N.A.; Malmberg, A.B. Neonatal capsaicin treatment abolishes formalin-induced spinal PGE2 release. NeuroReport 1997, 8, 2325–2329. [Google Scholar] [CrossRef] [PubMed]
- Le Bars, D.; Gozariu, M.; Cadden, S.W. Animal models of nociception. Pharmacol. Rev. 2001, 53, 597–652. [Google Scholar] [PubMed]
- O’Neill, J.; Brock, C.; Olesen, A.E.; Andresen, T.; Nilsson, M.; Dickenson, A.H. Unravelling the mystery of capsaicin: A tool to understand and treat pain. Pharmacol. Rev. 2012, 64, 939–971. [Google Scholar] [CrossRef] [PubMed]
- Ilie, M.A.; Caruntu, C.; Tampa, M.; Georgescu, S.-R.; Matei, C.; Negrei, C.; Ion, R.-M.; Constantin, C.; Neagu, M.; Boda, D. Capsaicin: Physicochemical properties, cutaneous reactions and potential applications in painful and inflammatory conditions. Exp. Ther. Med. 2019, 18, 916–925. [Google Scholar] [CrossRef]
- Patapoutian, A.; Peier, A.M.; Story, G.M.; Viswanath, V. ThermoTRP channels and beyond: Mechanisms of temperature sensation. Nat. Rev. Neurosci. 2003, 4, 529–539. [Google Scholar] [CrossRef]
- Olszewska, J.; Tęgowska, E. Opposite effect of capsaicin and capsazepine on behavioral thermoregulation in insects. J. Comp. Physiol. A 2011, 197, 1021–1026. [Google Scholar] [CrossRef]
- Gavva, N.R.; Bannon, A.W.; Surapaneni, S.; Hovland, D.N.; Lehto, S.G.; Gore, A.; Juan, T.; Deng, H.; Han, B.; Klionsky, L.; et al. The Vanilloid Receptor TRPV1 Is Tonically Activated In Vivo and Involved in Body Temperature Regulation. J. Neurosci. 2007, 27, 3366–3374. [Google Scholar] [CrossRef]
- Jancsó-Gábor, A.; Szolcsányi, J.; Jancsó, N. Stimulation and desensitization of the hypothalamic heat-sensitive structures by capsaicin in rats. J. Physiol. 1970, 208, 449–459. [Google Scholar] [CrossRef]
- Cabanac, M.; Cormareche-Leydier, M.; Poirier, L.J. The effect of capsaïcin on temperature regulation of the rat. Pflugers Arch. 1976, 366, 2. [Google Scholar]
- Mori, N.; Urata, T.; Fukuwatari, T. Thermal conditions influence changes in body temperature induced by intragastric administration of capsaicin in mice. Biosci. Biotechnol. Biochem. 2016, 80, 1–8. [Google Scholar] [CrossRef]
- Inagaki, H.; Kurganov, E.; Park, Y.; Furube, E.; Miyata, S.; Pak, Y. Oral gavage of capsaicin causes TRPV1-dependent acute hypothermia and TRPV1-independent long-lasting increase of locomotor activity in the mouse. Physiol. Behav. 2019, 206, 213–224. [Google Scholar] [CrossRef]
- Bujak, J.K.; Kosmala, D.; Szopa, I.M.; Majchrzak, K.; Bednarczyk, P. Inflammation, Cancer and Immunity-Implication of TRPV1 Channel. Front. Oncol. 2019, 9, 1087. [Google Scholar] [CrossRef]
- Toraya-Brown, S.; Fiering, S. Local tumour hyperthermia as immunotherapy for metastatic cancer. Int. J. Hyperth. 2014, 30, 531–539. [Google Scholar] [CrossRef]
- Du, Q.; Liao, Q.; Chen, C.; Yang, X.; Xie, R.; Xu, J. The Role of Transient Receptor Potential Vanilloid 1 in Common Diseases of the Digestive Tract and the Cardiovascular and Respiratory System. Front. Physiol. 2019, 10, 1064. [Google Scholar] [CrossRef]
- Lee, L.-Y.; Gu, Q. Role of TRPV1 in inflammation-induced airway hypersensitivity. Curr. Opin. Pharmacol. 2009, 9, 243–249. [Google Scholar] [CrossRef]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Front. Endocrinol. 2016, 7, 1541. [Google Scholar] [CrossRef]
- Ferrante, A. The immune cells in adipose tissue. Diabetes Obes. Metab. 2013, 15, 34–38. [Google Scholar] [CrossRef]
- Com, E.; Bourgeon, F.; Evrard, B.; Ganz, T.; Colleu, D.; Jégou, B.; Pineau, C. Expression of Antimicrobial Defensins in the Male Reproductive Tract of Rats, Mice, and Humans1. Biol. Reprod. 2003, 68, 95–104. [Google Scholar] [CrossRef]
- Dave, N.D.; Xiang, L.; Rehm, K.E.; Marshall, G.D. Stress and Allergic Diseases. Immunol. Allergy Clin. North Am. 2011, 31, 55–68. [Google Scholar] [CrossRef]
- Lynn, B. Capsaicin: Actions on nociceptive C-fibres and therapeutic potential. Pain 1990, 41, 61–69. [Google Scholar] [CrossRef]
- Franco-Cereceda, A.; Henke, H.; Lundberg, J.M.; Petermann, J.B.; Hökfelt, T.; Fischer, J.A. Calcitonin gene-related peptide (CGRP) in capsaicin-sensitive substance P-immunoreactive sensory neurons in animals and man: Distribution and release by capsaicin. Peptides 1987, 8, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Barr, T.; Gee, L.; Vickers, J.; Wymer, J.; Borsani, E.; Rodella, L.F.; Getsios, S.; Burdo, T.; Eisenberg, E.; et al. Keratinocyte expression of calcitonin gene-related peptide β: Implications for neuropathic and inflammatory pain mechanisms. Pain 2011, 152, 2036–2051. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, L.; Li, X.; Sahbaie, P.; Kingery, W.S.; Clark, J.D. Neuropeptides contribute to peripheral nociceptive sensitization by regulating interleukin-1beta production in keratinocytes. Anesth. Analg. 2011, 113, 175–183. [Google Scholar] [PubMed]
- Sandilands, A.; Sutherland, C.; Irvine, A.; McLean, W.H.I. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef]
- Liu, T.; Ji, R.-R. New insights into the mechanisms of itch: Are pain and itch controlled by distinct mechanisms? Pflüg. Arch. Eur. J. Phy. 2013, 465, 1671–1685. [Google Scholar] [CrossRef]
- Rinaldi, G. The Itch-Scratch Cycle: A Review of the Mechanisms. Dermatol. Pr. Concept. 2019, 9, 90–97. [Google Scholar] [CrossRef]
- Oaklander, A.L. Neuropathic itch. Semin. Cutan. Med. Surg. 2011, 30, 87–92. [Google Scholar]
- Satinoff, E.; Liran, J.; Clapman, R. Aberrations of circadian body temperature rhythms in rats with medial preoptic lesions. Am. J. Physiol. Integr. Comp. Physiol. 1982, 242, R352–R357. [Google Scholar] [CrossRef]
- Romanovsky, A.A.; Almeida, M.C.; Garami, A.; Steiner, A.A.; Norman, M.H.; Morrison, S.F.; Nakamura, K.; Burmeister, J.J.; Nucci, T.B. The transient receptor potential vanilloid-1 channel in thermoregulation: A thermosensor it is not. Pharmacol. Rev. 2009, 61, 228–261. [Google Scholar] [CrossRef]
- Kimura, Y.; Honda, M.; Sasaki, R.; Yumioka, T.; Iwamoto, H.; Tsounapi, P.; Morizane, S.; Hikita, K.; Osaki, M.; Okada, F.; et al. The circadian rhythm of bladder clock genes in the spontaneously hypersensitive rat. PLoS ONE 2019, 14, e0220381. [Google Scholar]
- Szelényi, Z.; Hummel, Z.; Szolcsányi, J.; Davis, J.B. Daily body temperature rhythm and heat tolerance in TRPV1 knockout and capsaicin pretreated mice. Eur. J. Neurosci. 2004, 19, 1421–1424. [Google Scholar] [CrossRef]
- Yang, S.-C.; Chen, C.-L.; Yi, C.-H.; Liu, T.-T.; Shieh, K.-R. Changes in Gene Expression Patterns of Circadian-Clock, Transient Receptor Potential Vanilloid-1 and Nerve Growth Factor in Inflamed Human Esophagus. Sci. Rep. 2015, 5, 13602. [Google Scholar] [CrossRef]
- Herrero, L.; Valcarcel, L.; Silva, C.; Albert, N.; Díez-Noguera, A.; Cambras, T.; Serra, D. Altered Circadian Rhythm and Metabolic Gene Profile in Rats Subjected to Advanced Light Phase Shifts. PLoS ONE 2015, 10, e0122570. [Google Scholar] [CrossRef]
- Husse, J.; Eichele, G.; Oster, H. Synchronization of the mammalian circadian timing system: Light can control peripheral clocks independently of the SCN clock. BioEssays 2015, 37, 1119–1128. [Google Scholar] [CrossRef]
- Tamaru, T.; Hattori, M.; Honda, K.; Benjamin, I.; Ozawa, T.; Takamatsu, K. Synchronization of Circadian Per2 Rhythms and HSF1-BMAL1:CLOCK Interaction in Mouse Fibroblasts after Short-Term Heat Shock Pulse. PLoS ONE 2011, 6, e24521. [Google Scholar] [CrossRef]
- Park, K.; Kang, H.M. Circadian expression of clock genes in the rat eye and brain. Mol. Cells 2006, 22, 285–290. [Google Scholar]
- Sawka, M.N.; Leon, L.R.; Montain, S.J.; Sonna, L.A. Integrated Physiological Mechanisms of Exercise Performance, Adaptation, and Maladaptation to Heat Stress. Compr. Physiol. 2011, 1, 1883–1928. [Google Scholar] [CrossRef]

{kind=link}
| Study By | Study Design | Subject | Measurement | Results |
|---|---|---|---|---|
| Simone et al. [6]. | Intradermal injection of 100 µg of capsaicin | Cat | Innocuous mechanical stimulation | Sensitization of cat dorsal horn neurons |
| Donnerer et al. [5]. | Daily subcutaneous injection of 50 mg/kg of capsaicin | Rat | RT-PCR | Desensitization of TRPV1 in dorsal root ganglia |
| Vyklicky et al. [22]. | Treatment with 1 µM of capsaicin | HEK293T cells transfected with rat TRPV1 | Whole-cell current responses | Selectively excite and subsequently desensitize nociceptive neurons |
| Srbely et al. [7]. | 45 degrees heat for 10 min then received topical capsaicin cream (0.075%) | Human | Central sensitization at the C(5) segment and mechanical cutaneous sensitivity | An increase in central sensitization related to allodynia |
| Newson et al. [11]. | 0.1 M (5 mg/mL) neonatal capsaicin treatment | Rat | Superior staining for neuronal counting | Life-long loss of sensory neurons expressing TRPV1 channels. |
| Study By | Study Design | Subject | Measurement | Results |
|---|---|---|---|---|
| Meller et al. [12]. | Neonatal capsaicin treatment (50 mg/kg) | Rat | Thermal hyperalgesia in a sciatic nerve ligation model | Loss of thermal hyperalgesia |
| Dougherty et al. [26]. | Intradermal capsaicin administraion (0.1 mL, 1% solution) | Monkey | Cutaneous mechanical and thermal stimuli by an electrode | Loss of thermal and mechanical stimuli |
| Baamonde et al. [25]. | Intra-plantar administration of 10 μg of capsaicin | Mouse | Licking behavior following complete Freund’s adjuvant injection | Induction of long-lasting analgesia for at least 2 weeks |
| Jeong et al. [10]. | Neonatal capsaicin treatment (50 mg/kg) | Rat | Paw-withdrawal latency to radiant infrared heat stimulation, and intake volume of capsaicin water | Desensitization to noxious heat stimuli and impaired sensing of capsaicin |
| Nagy et al. [30]. | Neonatal capsaicin treatment (5 to 100 mg/kg) | Rat | Noxious thermal, mechanical, and chemical stimuli | Increased nociceptive threshold by higher doses of capsaicin, but the uncertain correlation with the extent of loss of primary afferent fiber |
| Study By | Study Design | Subject | Measurement | Results |
|---|---|---|---|---|
| Jancso-Gabor et al. [38]. | Subcutaneous injection of capsaicin (0.25 mg) | Rat | Rectal temperature | A decrease in the rectal temperature for 120 min |
| Cabanac et al. [39]. | Subcutaneous injection of capsaicin (6-66 mg, cumulative) | Rat | Rectal temperature | Induction of hyperthermia by a decreased salivary secretion |
| Jeong et al. [13]. | Neonatal capsaicin treatment (50 mg/kg) | Rat | Core body temperature | Induction of chronic hyperthermia |
| Mori et al. [40]. | Intragastric administration of capsaicin (10 and 15 mg/kg) | Mouse | Colonic temperature | Decreased colonic temperature by 15 mg/kg capsaicin, and increased colonic temperature by but 10 mg/kg capsaicin |
| Inagaki et al. [41]. | Oral gavage of capsaicin (10-20 mg/kg) | Mouse | Core body and tail surface temperature | A decrease in core body temperature and an increase in tail surface temperature |
| Study By | Study Design | Subject | Methods | Results |
|---|---|---|---|---|
| Ljungdahl et al. [15]. | Neonatal capsaicin treatment (50 mg/kg) | Mouse | Herpes simplex virus infection | The extensive infection of glial cells proximal by an amplified degenerative effect on central branches of the substanceP |
| Bowden et al. [14]. | Neonatal capsaicin treatment (50 mg/kg) | Rat | Mycoplasma pulmonis infection | Severe infection in the airway mucosa by reduction of substance P immunoreactive nerve fiber |
| Jeong et al. [13]. | Neonatal capsaicin treatment (50 mg/kg) | Rat | Staphylococcus aureus infection | An increase in the Staphylococcus aureus infection by disruption of the immune defense system |
| Study By | Study Design | Subject | Measurement | Results |
|---|---|---|---|---|
| Back et al. [16]. | Neonatal capsaicin treatment (50 mg/kg) | Rat | Scratching behavior and dermatitis score | Severe scratching behavior and cutaneous lesions by loss of immunoreactions in sensory nerve |
| Thomas et al. [18]. | Neonatal capsaicin treatment (50 mg/kg) | Rat | Damaged skin and behaviors of scratching | Induction of scratching behavior resulting in the total area of skin damage by loss of c-fiber |
| Jeong et al. [17]. | Neonatal capsaicin treatment (50 mg/kg) + juvenile obesity | Rat | Dermatitis score | Robust development of dermatitis by juvenile obesity |
| Study By | Study Design | Subject | Results |
|---|---|---|---|
| Szelényi et al. [61]. | Subcutaneous injection of capsaicin (15 and 30 mg/kg) on two consecutive days | Mouse | Higher daily body temperature amplitude in the capsaicin pretreated mice by vanilloid receptor-mediated altering action on temperature regulation |
| Jeong et al. [10]. | Neonatal capsaicin treatment (50 mg/kg) | Rat | The reverse amplitude of the circadian body temperature rhythm in neonatal capsaicin treatment rats caused by altering the expression of clock genes in the hypothalamus and liver |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, K.-Y. Changes in TRPV1-Mediated Physiological Function in Rats Systemically Treated With Capsaicin on the Neonate. Int. J. Mol. Sci. 2020, 21, 3143. https://doi.org/10.3390/ijms21093143
Jeong K-Y. Changes in TRPV1-Mediated Physiological Function in Rats Systemically Treated With Capsaicin on the Neonate. International Journal of Molecular Sciences. 2020; 21(9):3143. https://doi.org/10.3390/ijms21093143
Chicago/Turabian StyleJeong, Keun-Yeong. 2020. "Changes in TRPV1-Mediated Physiological Function in Rats Systemically Treated With Capsaicin on the Neonate" International Journal of Molecular Sciences 21, no. 9: 3143. https://doi.org/10.3390/ijms21093143
APA StyleJeong, K.-Y. (2020). Changes in TRPV1-Mediated Physiological Function in Rats Systemically Treated With Capsaicin on the Neonate. International Journal of Molecular Sciences, 21(9), 3143. https://doi.org/10.3390/ijms21093143