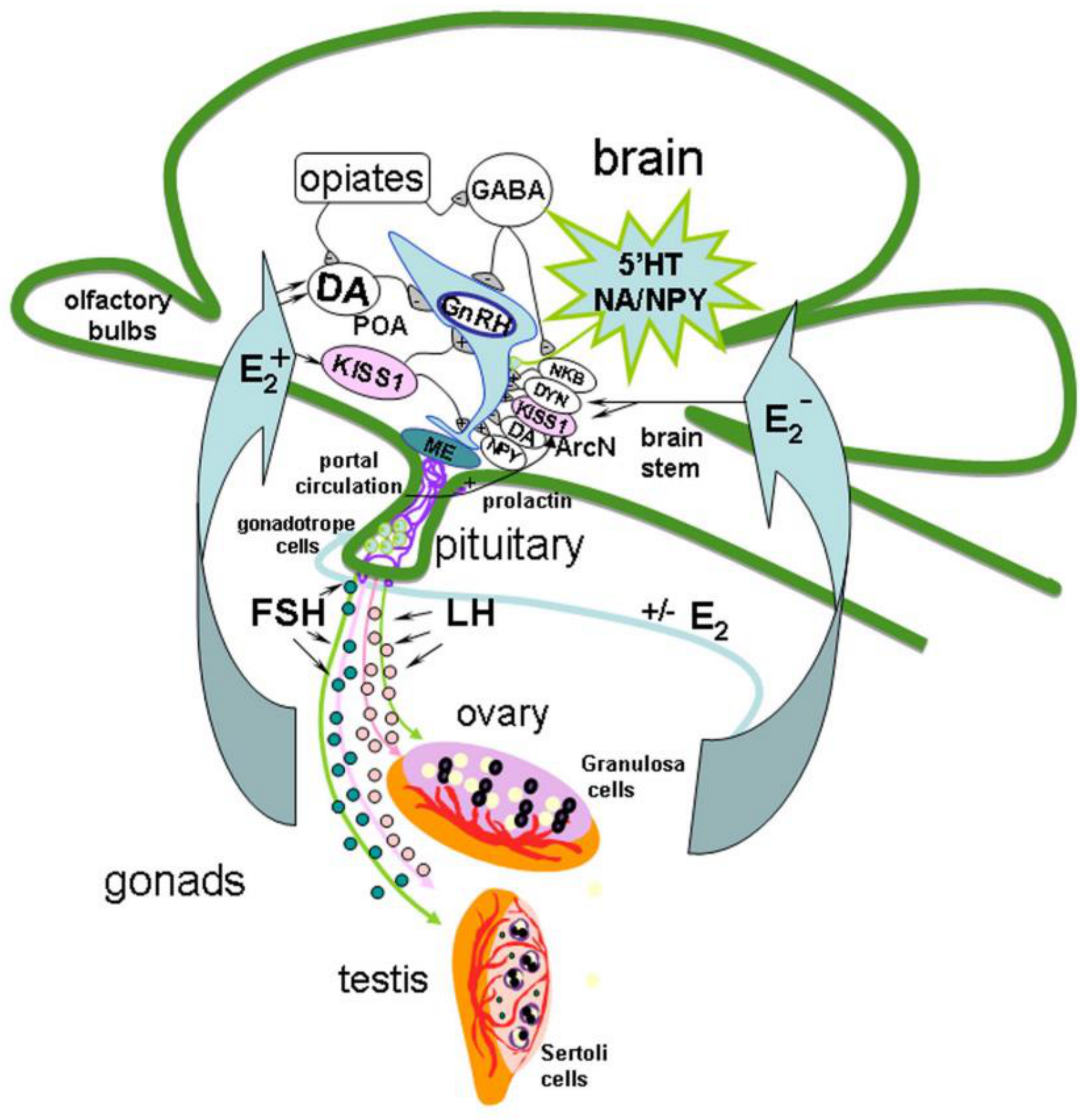
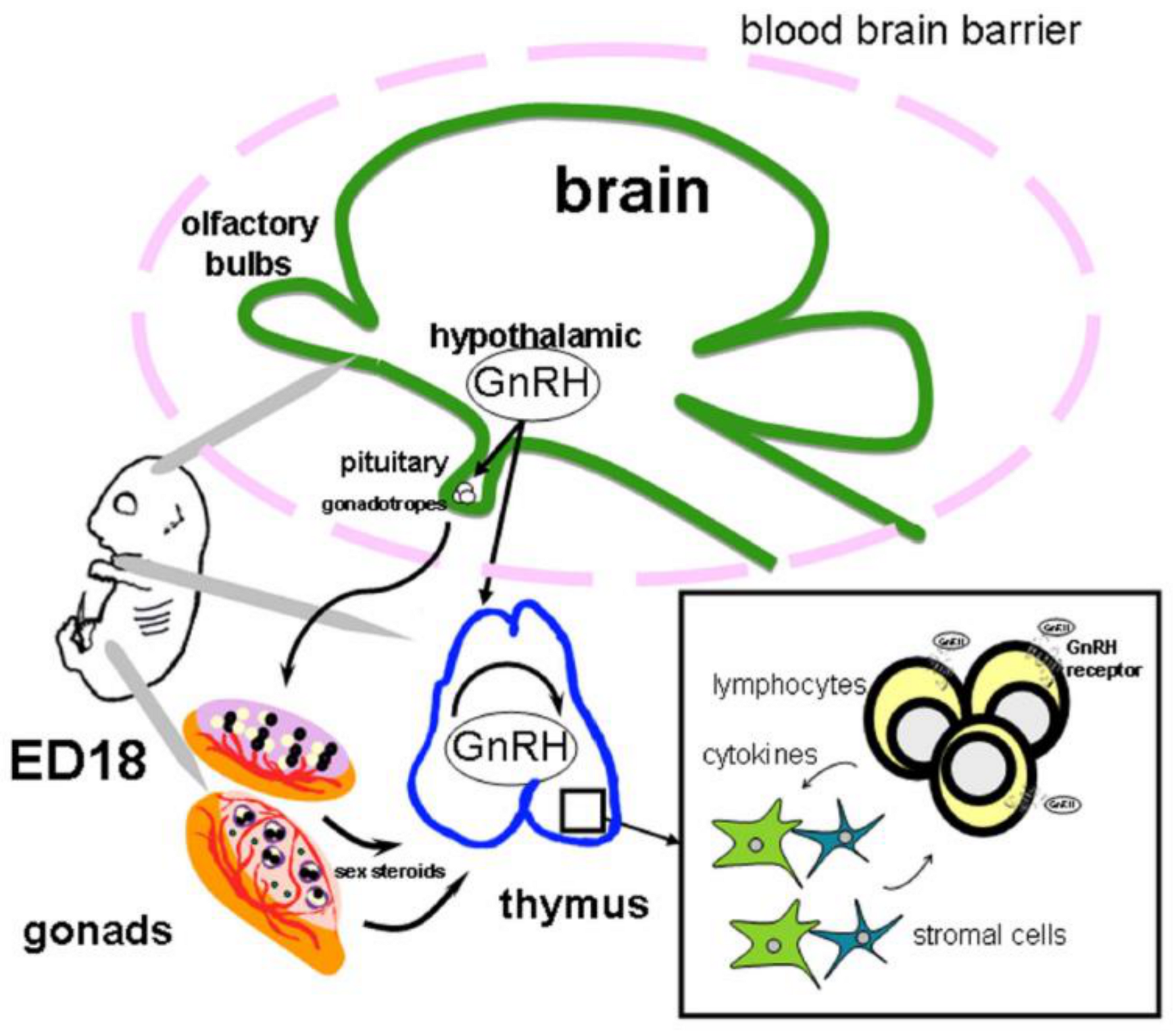
Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions


{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Zakharova, L.; Sharova, V.; Izvolskaia, M. Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions. Int. J. Mol. Sci. 2021, 22, 114. https://doi.org/10.3390/ijms22010114
Zakharova L, Sharova V, Izvolskaia M. Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions. International Journal of Molecular Sciences. 2021; 22(1):114. https://doi.org/10.3390/ijms22010114
Chicago/Turabian StyleZakharova, Liudmila, Viktoria Sharova, and Marina Izvolskaia. 2021. "Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions" International Journal of Molecular Sciences 22, no. 1: 114. https://doi.org/10.3390/ijms22010114
APA StyleZakharova, L., Sharova, V., & Izvolskaia, M. (2021). Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions. International Journal of Molecular Sciences, 22(1), 114. https://doi.org/10.3390/ijms22010114