The Influence of New Silicate Cement Mineral Trioxide Aggregate (MTA Repair HP) on Metalloproteinase MMP-2 and MMP-9 Expression in Cultured THP-1 Macrophages

 ,
,
Abstract
1. Introduction
2. Results
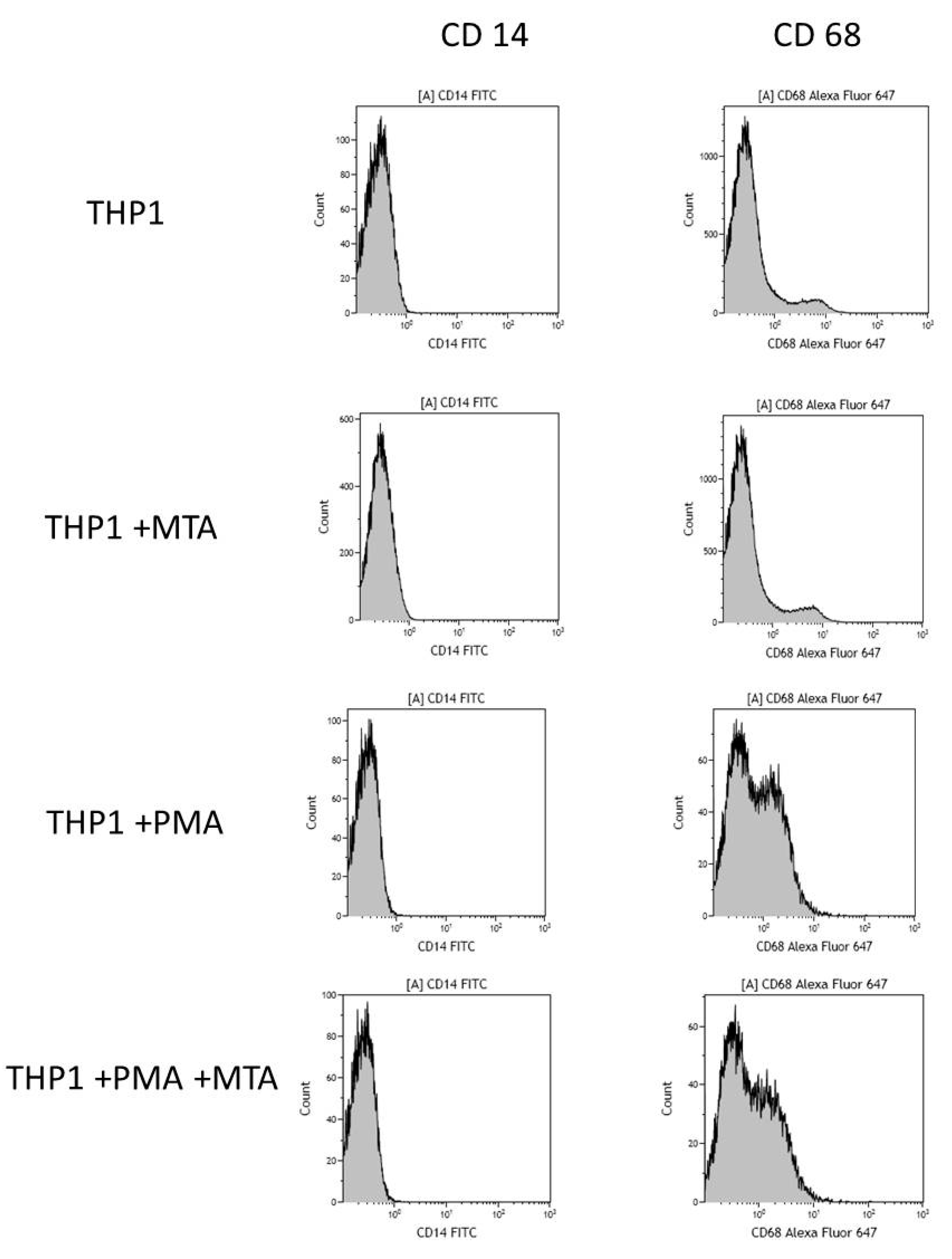
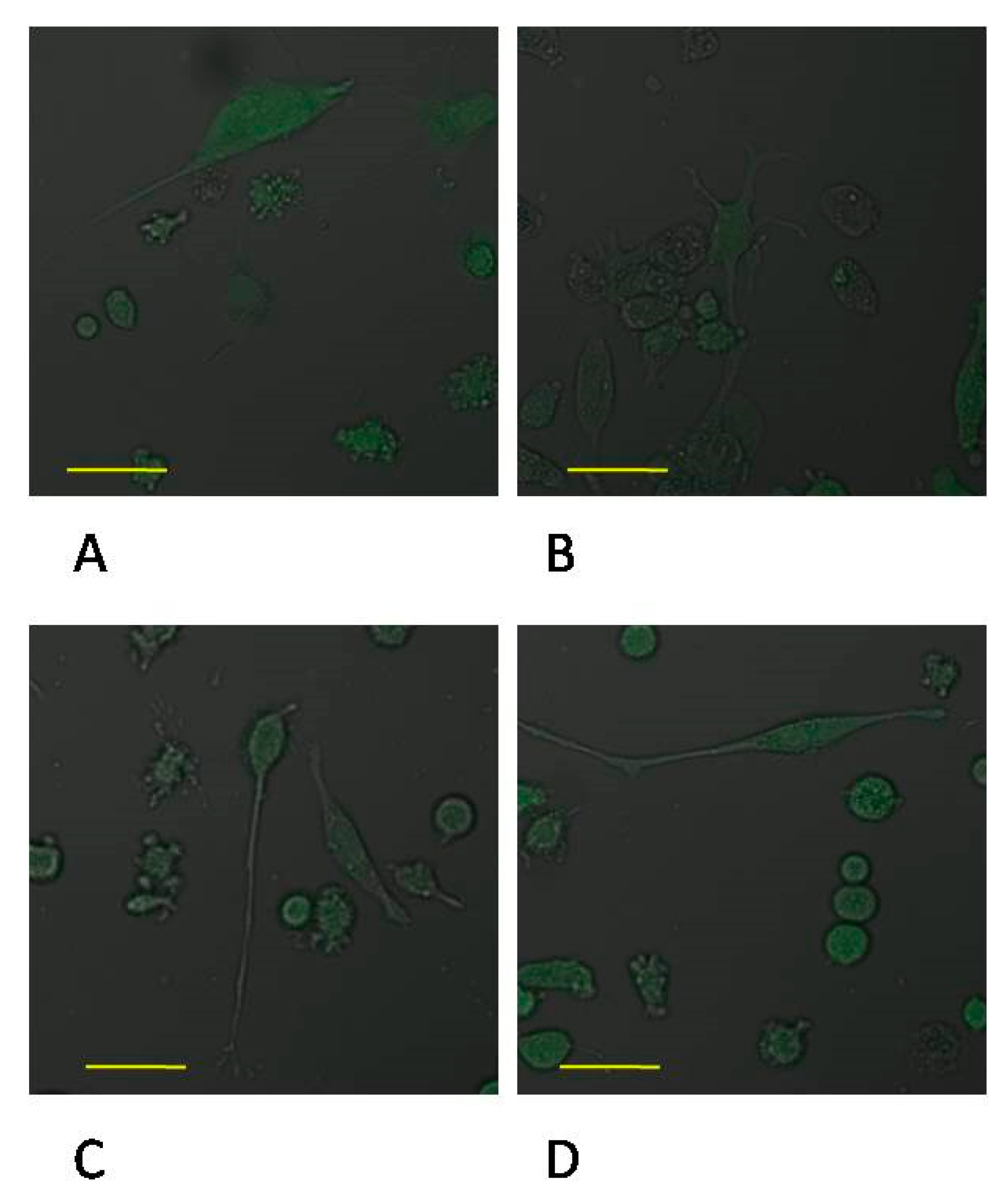
2.1. MTA Repair HP-Induced Activation of THP-1 Monocytes
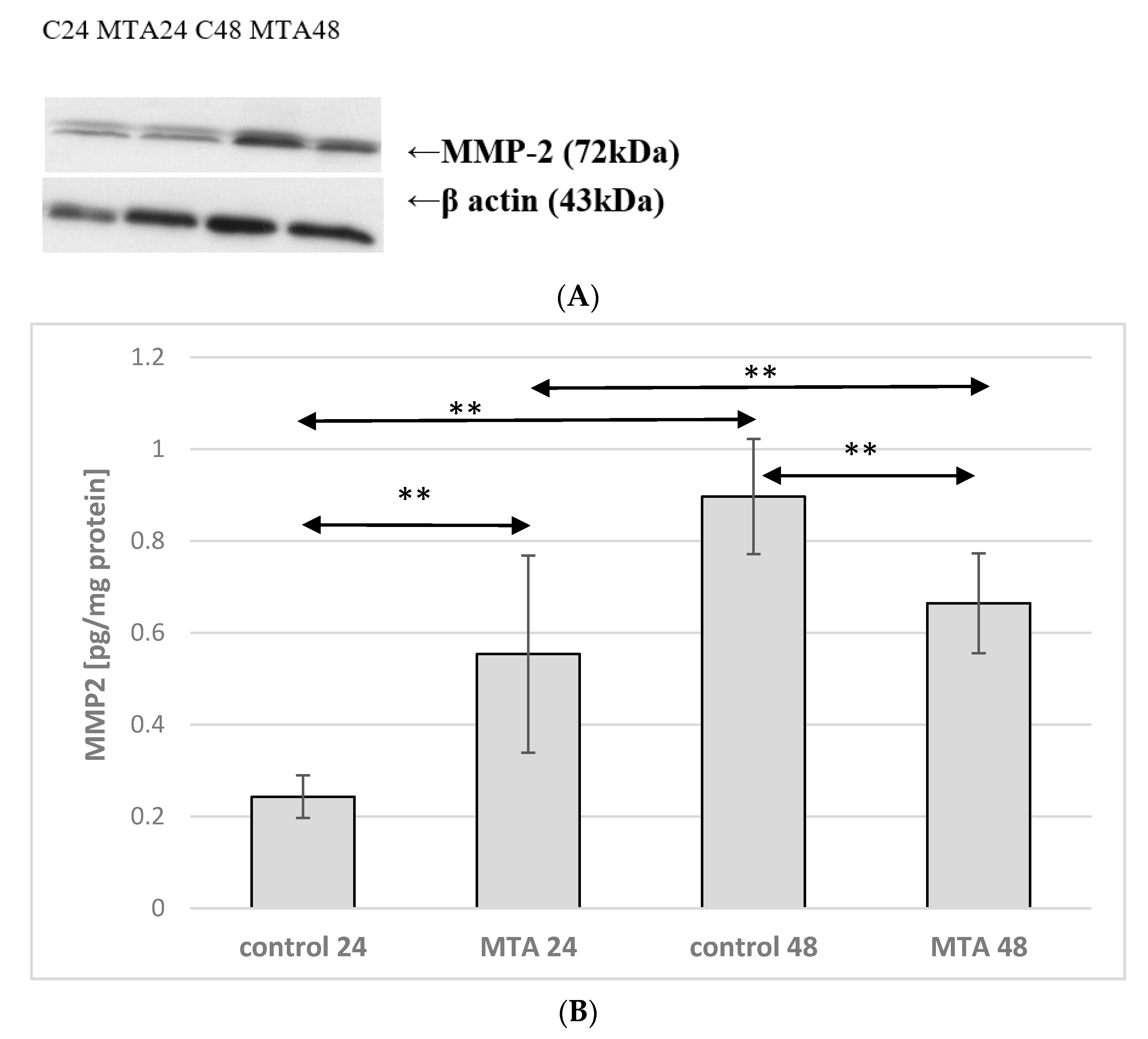
2.2. Expression of MMP-2 in Monocytes
2.3. Expression of MMP-2 in Macrophages
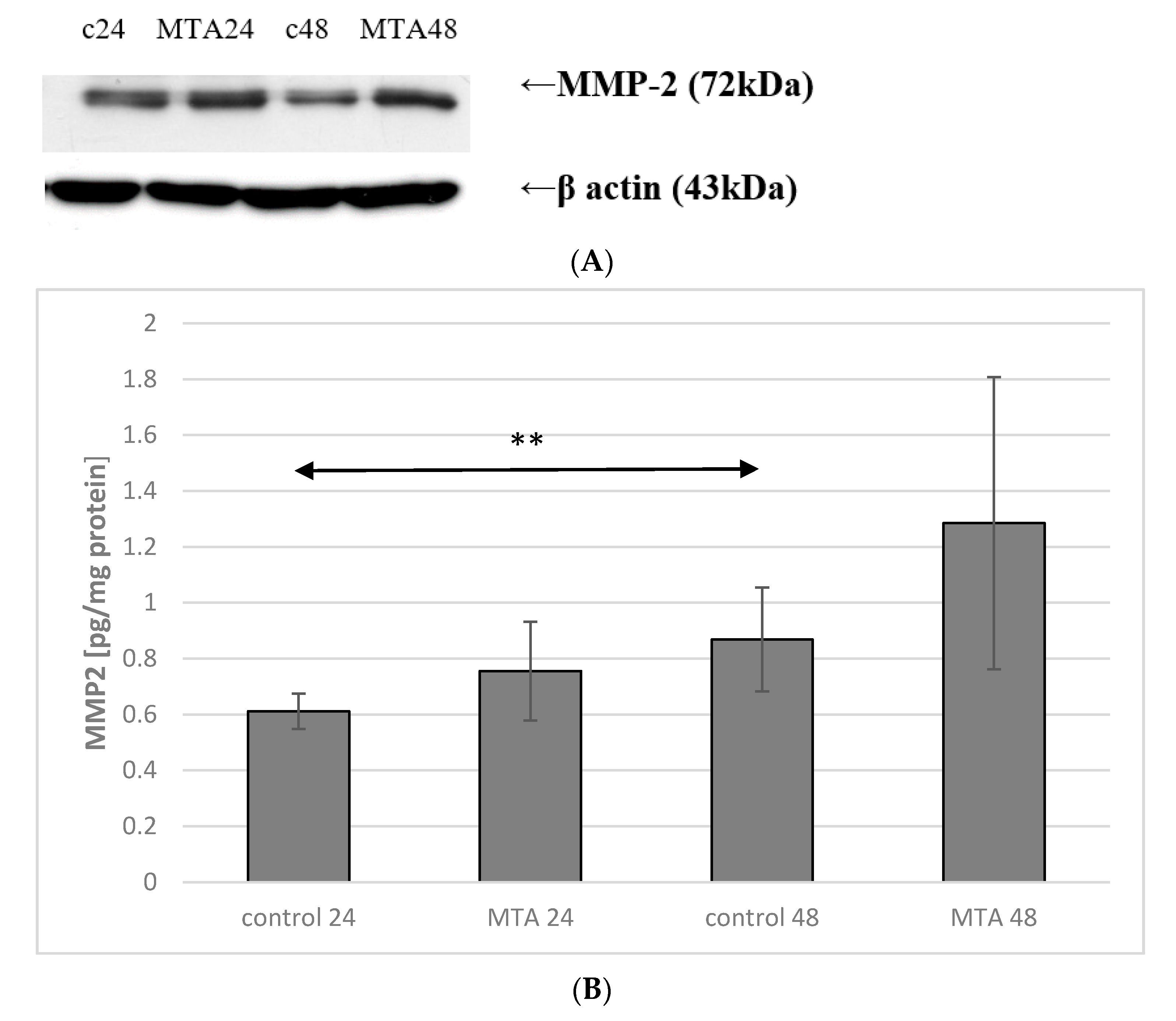
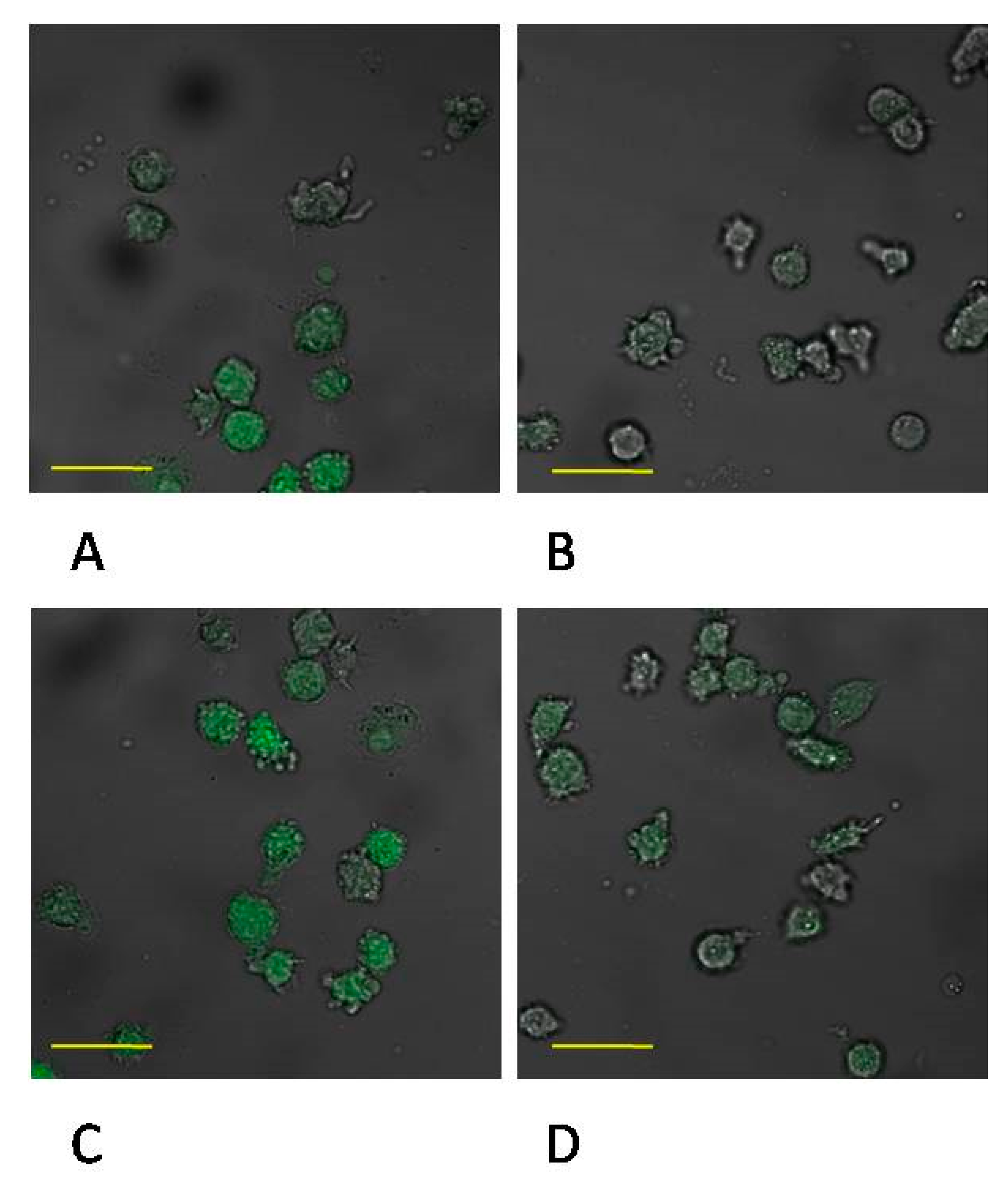
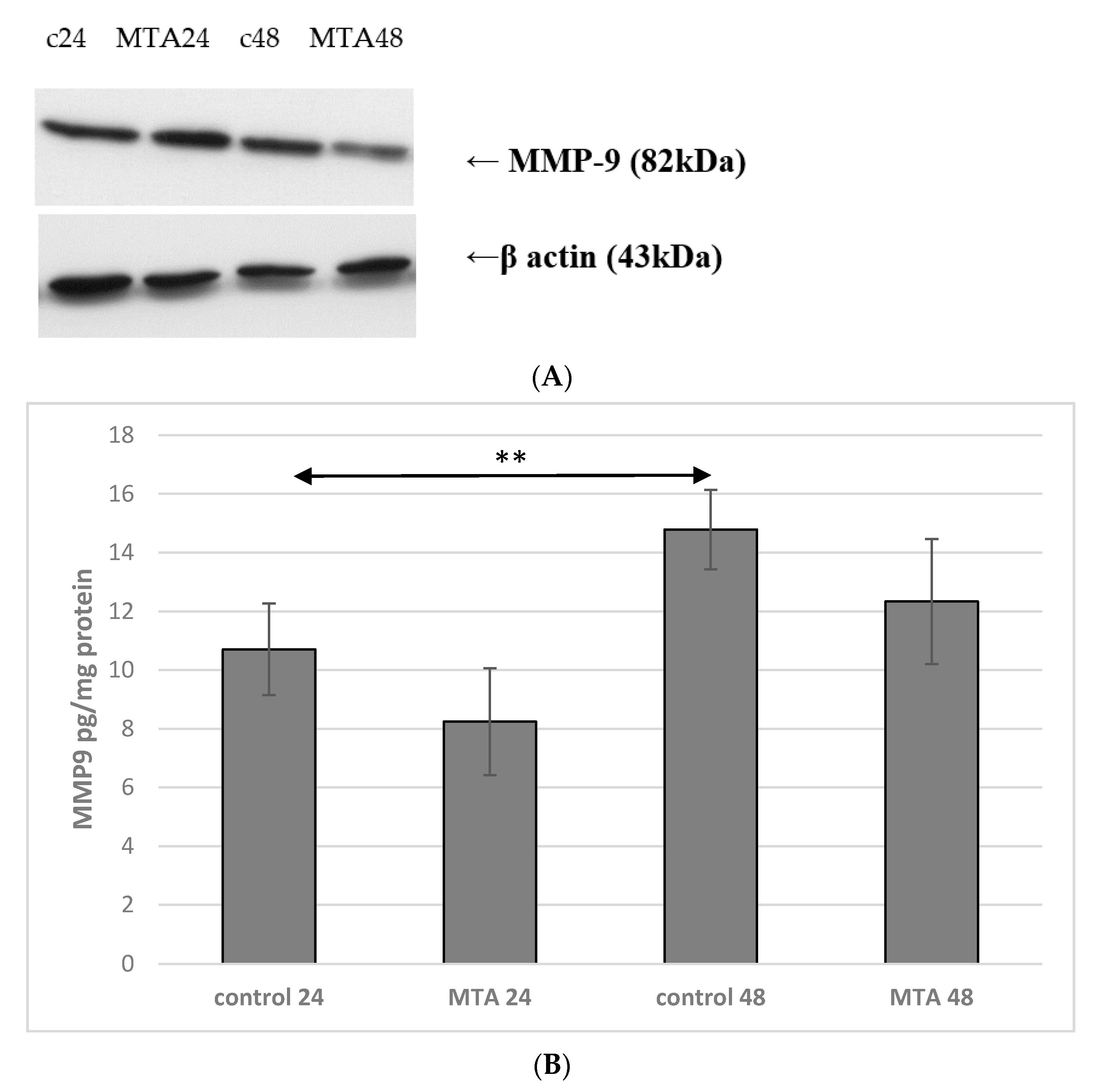
2.4. Expression of MMP-9 in Monocytes
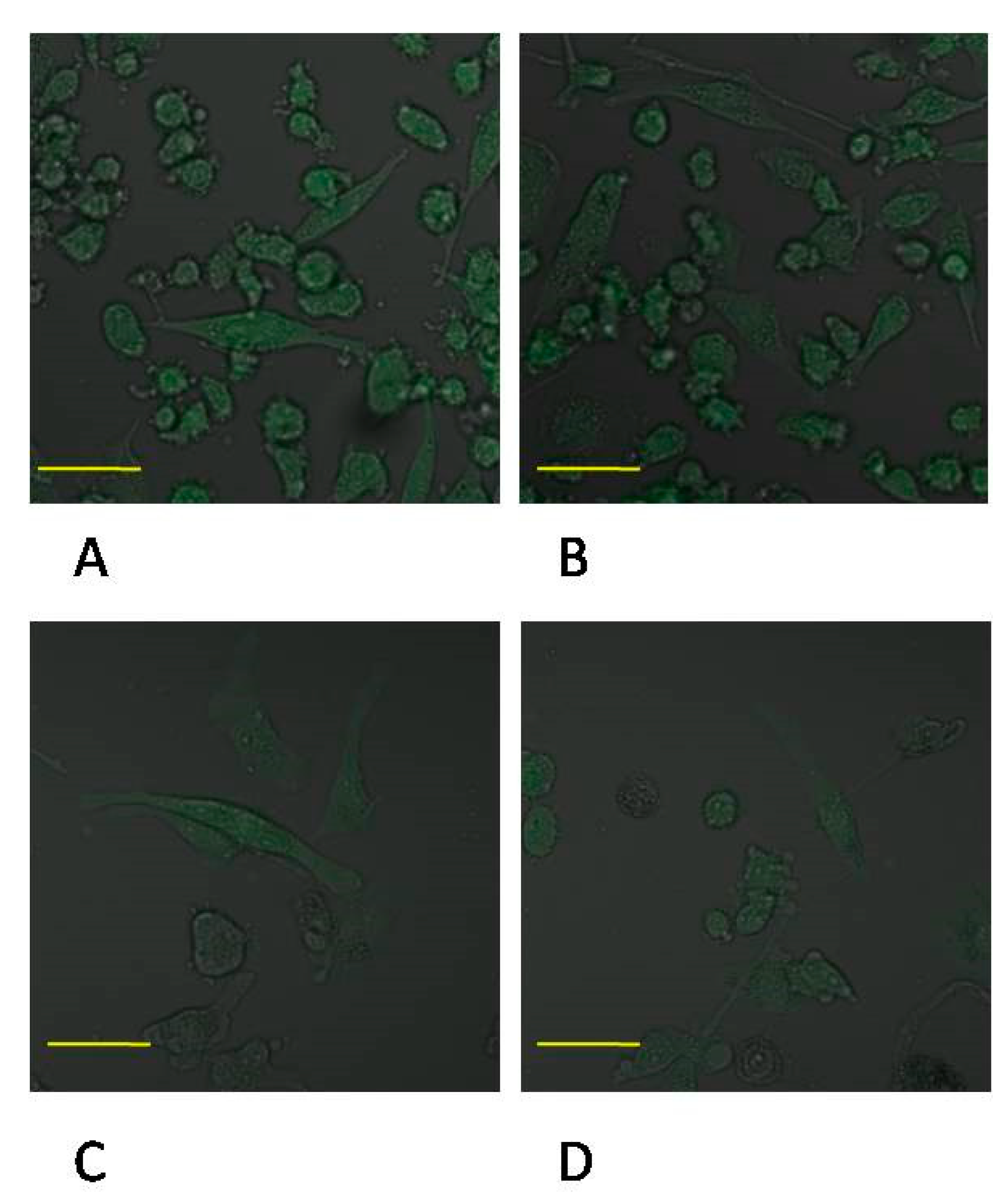
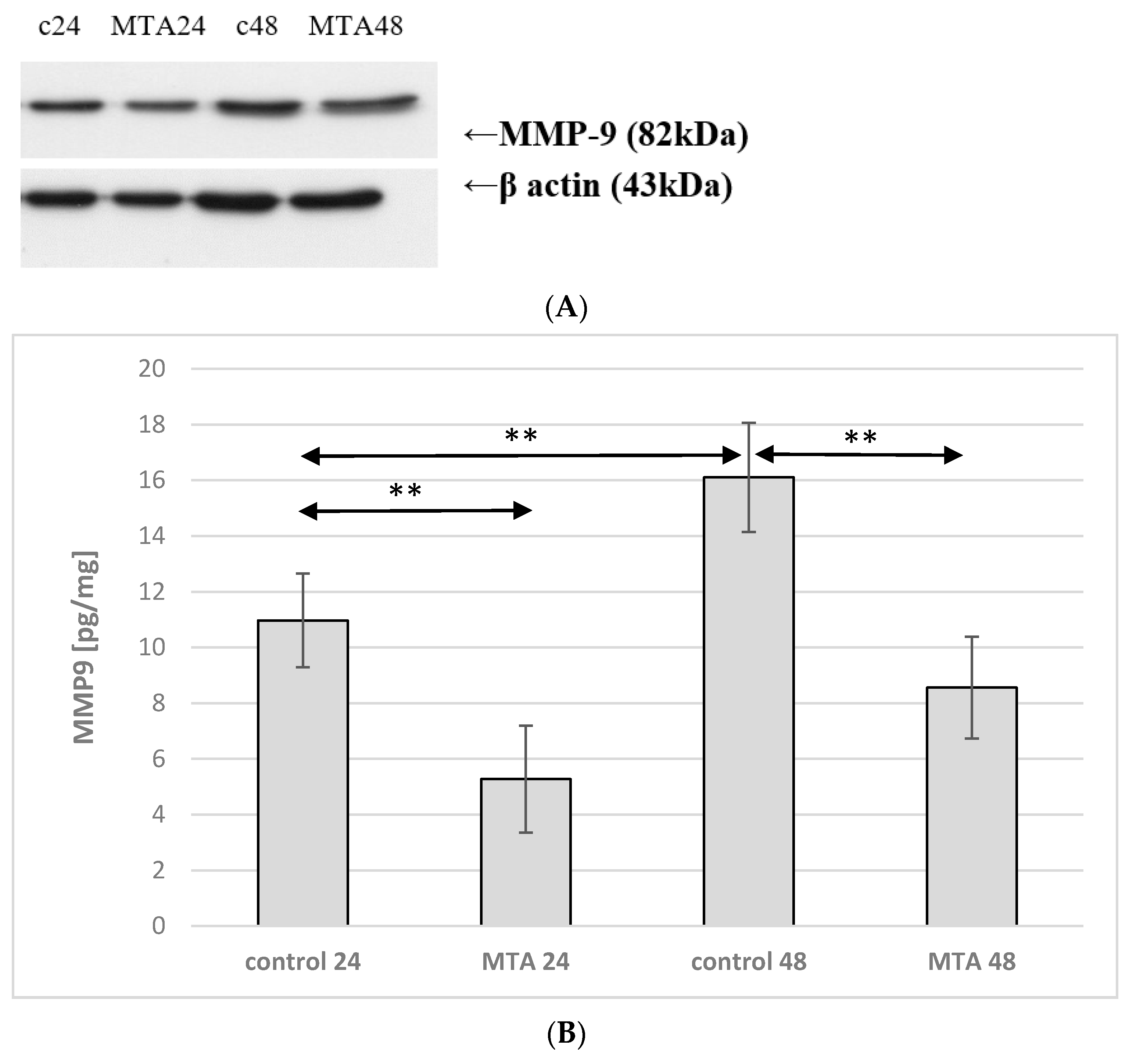
2.5. Expression of MMP-9 in Macrophages
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Chemical Properties of MTA Repair HP
4.3. Preparation of MTA Repair HP
4.4. Cell Culture and Treatment
4.5. MTA Repair HP-Induced Inflammatory Reaction in THP-1 Monocytes
4.6. Differentiation of THP-1 Cells into Macrophages. Flow Cytometry Measurement
4.7. MTA Repair HP-Induced Inflammatory Reaction in Macrophages
4.8. Determination of MMP-2 Expression by ELISA Method
4.9. The Determination of MMP-9 Expression by ELISA Method
4.10. Western Blot Analysis of MMP-2 and MMP-9 Expression
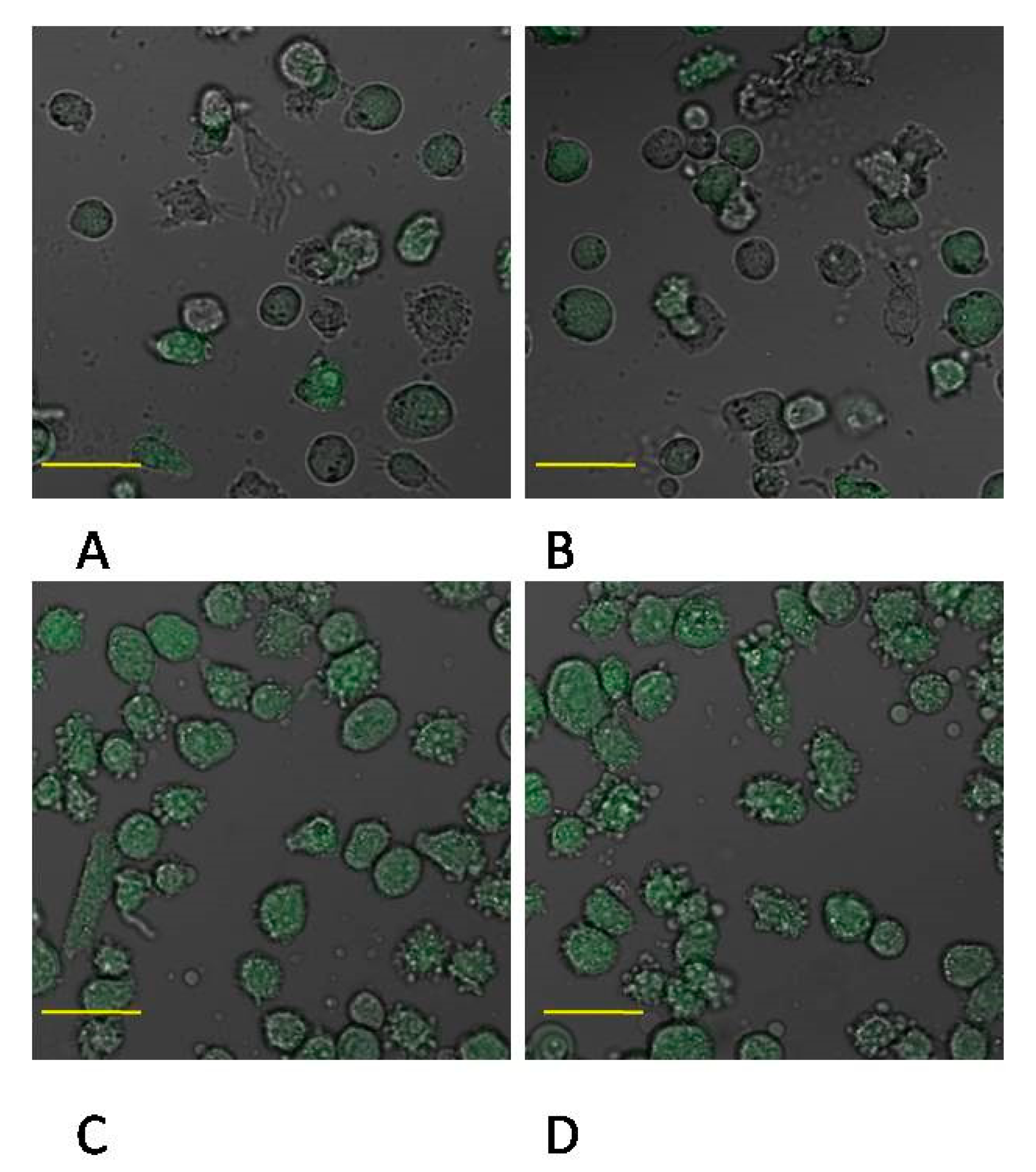
4.11. Confocal Microscopy Imaging of MMP-2 and MMP-9 Expression
4.12. Protein Concentration
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Silva, E.J.; Carvalho, N.K.; Zanon, M.; Senna, P.M.; De-Deus, G.; Zuolo, M.L.; Zaia, A.A. Push-out bond strength of MTA HP, a new high-plasticity calcium silicate-based cement. Braz. Oral Res. 2016, 30. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.M.A.; Sassone, L.M.; Gonçalves, A.S.; de Carvalho, J.J.; Tomás-Catalá, C.J.; García-Bernal, D.; Oñate-Sánchez, R.E.; Rodríguez-Lozano, F.J.; Silva, E.J.N.L. Physicochemical, cytotoxicity and in vivo biocompatibility of a high-plasticity calcium-silicate based material. Sci. Rep. 2019, 9, 3933. [Google Scholar] [CrossRef]
- Tawil, P.Z.; Duggan, D.J.; Galicia, J.C. MTA: A clinical review. Compend. Contin. Educ. Dent. 2015, 36, 247–264. [Google Scholar] [PubMed]
- Kaur, M.; Singh, H.; Dhillon, J.S.; Batra, M.; Saini, M. MTA versus Biodentine: Review of literature with a comparative analysis. J. Clin. Diagn. Res. 2017, 11, ZG01–ZG05. [Google Scholar] [CrossRef] [PubMed]
- Dammaschke, T.; Nowicka, A.; Lipski, M.; Domenico, R. Histological evaluation of hard tissue formation after direct pulp capping with a fast-setting mineral trioxide aggregate (RetroMTA) in humans. Clin. Oral Investig. 2019, 23, 4289–4299. [Google Scholar] [CrossRef] [PubMed]
- Giraud, T.; Jeanneau, C.; Bergmann, M.; About, I. Tricalcium silicate capping materials modulate pulp healing and inflammatory activity in vitro. J. Endod. 2018, 44, 1686–1691. [Google Scholar] [CrossRef]
- Paula, A.; Carrilho, E.; Laranjo, M.; Abrantes, A.M.; Casalta-Lopes, J.; Botelho, M.F.; Marto, C.M.; Ferreira, M.M. Direct pulp capping: Which is the most effective biomaterial? A retrospective clinical study. Materials (Basel) 2019, 12, 3382. [Google Scholar] [CrossRef]
- Nowicka, A.; Lipski, M.; Parafiniuk, M.; Sporniak-Tutak, K.; Lichota, D.; Kosierkiewicz, A.; Kaczmarek, W.; Buczkowska-Radlińska, J. Response of human dental pulp capped with Biodentine and Mineral Trioxide Aggregate. J. Endod. 2013, 39, 743–747. [Google Scholar] [CrossRef]
- Lipski, M.; Nowicka, A.; Kot, K.; Postek-Stefańska, L.; Wysoczańska-Jankowicz, I.; Borkowski, L.; Andersz, P.; Jarząbek, A.; Grocholewicz, K.; Sobolewska, E.; et al. Factors affecting the outcomes of direct pulp capping using Biodentine. Clin. Oral Investig. 2018, 22, 2021–2029. [Google Scholar] [CrossRef]
- Tomás-Catalá, C.J.; Collado-González, M.; García-Bernal, D.; Oñate-Sánchez, R.E.; Forner, L.; Llena, C.; Lozano, A.; Castelo-Baz, P.; Moraleda, J.M.; Rodríguez-Lozano, F.J. Comparative analysis of the biological effects of the endodontic bioactive cements MTA-Angelus, MTA Repair HP and NeoMTA Plus on human dental pulp stem cells. Int. Endod. J. 2017, 50 (Suppl. S2), e63–e72. [Google Scholar] [CrossRef]
- Da Fonseca, T.S.; Silva, G.F.; Guerreiro-Tanomaru, J.M.; Delfino, M.M.; Sasso-Cerri, E.; Tanomaru-Filho, M.; Cerri, P.S. Biodentine and MTA modulate immunoinflammatory response favoring bone formation in sealing of furcation perforations in rat molars. Clin. Oral Investig. 2018. [Google Scholar] [CrossRef] [PubMed]
- Futami, T.; Fujii, N.; Ohnihi, H.; Taguchi, N.; Kusakari, H.; Ohshima, H.; Maeda, T. Tissue response to titanium implants in the rat maxilla: Ultrastructural and histochemical observations of the bone-titanium interface. J. Periodontol. 2000, 71, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.; Vreeburg, R.A.M.; Savelkoul, H.F.J.; Wichers, H.J. Transcription profiles of LPS-stimulated THP-1 monocytes and macrophages: A tool to study inflammation modulating effects of food-derived compounds. Food Funct. 2010, 1, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro model for immunomodulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- Brown, B.N.; Ratner, B.D.; Goodman, S.B.; Amar, S.; Badylak, S.F. Macrophage polarization: An opportunity for improved outcomes in biomaterials and regenerative medicine. Biomaterials 2012, 33, 792–802. [Google Scholar] [CrossRef]
- Shirakura, M.; Fujii, N.; Ohnishi, H.; Taguchi, Y.; Ohshima, H.; Nomura, S.; Maeda, T. Tissue response to titanium implantation in the rat maxilla, with special reference to the effects of surface conditions on bone formation. Clin. Oral Implants Res. 2003, 14, 687–696. [Google Scholar] [CrossRef]
- Refai, A.K.; Textor, M.; Brunette, D.M.; Waterfield, J.D. Effect of titanium surface topography on macrophage activation and secretion of proinflammatory cytokines and chemokines. J. Biomed Mater. Res. A 2004, 70, 194–205. [Google Scholar] [CrossRef]
- Auger, M.J.; Ross, J.A. The biology of the macrophage. In The Macrophage; IRL Press: New York, NY, USA, 1992; pp. 3–74. [Google Scholar]
- Van Furth, R. Human monocytes and cytokines. Res. Immunol. 1998, 149, 719–720. [Google Scholar] [CrossRef]
- Gordon, S. The macrophage. Bioessays 1995, 17, 977–986. [Google Scholar] [CrossRef]
- Cawston, T.E.; Wilson, A.J. Understanding the role of tissue degrading enzymes and their inhibitors in development and diease. Best Pract. Res. Clin. Rheumatol. 2006, 20, 983–1002. [Google Scholar] [CrossRef]
- Lamort, A.S.; Gravier, R.; Laffitte, A.; Juliano, L.; Zani, M.L.; Moreau, T. New insights into the substrate specificity of macrophage elastase MMP-12. Biol. Chem. 2006, 397, 469–484. [Google Scholar] [CrossRef] [PubMed][Green Version]
- McCawley, L.J.; Matrisian, L.M. Matrix metalloproteinases: They’re not just for matrix anymore! Curr. Opin. Cell Biol. Engl. 2001, 13, 534–540. [Google Scholar] [CrossRef]
- Parks, W.C.; Wilson, C.L.; Lopez-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. Engl. 2004, 4, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Macwan, C.; Anshula Deshpande, A. Mineral trioxide aggregate (MTA) in dentistry: A review of literature. J. Oral Res. Rev. 2014, 6, 71–74. [Google Scholar] [CrossRef]
- Abou ElReash, A.; Hamama, H.; Abdo, W.; Wu, Q.; Zaen El-Din, A.; Xiaoli, X. Biocompatibility of new bioactive resin composite versus calcium silicate cements: An animal study. BMC Oral Health 2019, 19, 194. [Google Scholar] [CrossRef]
- Guimarães, M.B.; Prati, C.; Duarte, M.A.H.; Bramante, C.M.; Gandolfi, M.G. Physicochemical properties of calcium silicate-based formulations MTA Repair HP and MTA Vitalcem. J. Appl. Oral Sci. 2018, 26, e2017115. [Google Scholar] [CrossRef]
- Zhu, C.; Ju, B.; Ni, R. Clinical outcome of direct pulp capping with MTA or calcium hydroxide: A systematic review and meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 17055–17060. [Google Scholar]
- Shi, S.; Bao, Z.F.; Chen, X.; Zhang, D.D. Cytotoxicity of a novel endodontic treatment material iRoot BP Plus to human gingival fibroblasts. Shanghai Kou Qiang Yi Xue 2014, 23, 681–684. [Google Scholar]
- Zafar, K.; Jamal, S.; Ghafoor, R. Bio-active cements-Mineral Trioxide Aggregate based calcium silicate materials: A narrative review. J. Pak. Med. Assoc. 2020, 70, 497–504. [Google Scholar] [CrossRef]
- Adıgüzel, M.; Ahmetoğlu, F.; Eldeniz, A.Ü.; Tekin, M.G.; Göğebakan, B. Comparison of cytotoxic effects of calcium silicate-based materials on human pulp fibroblasts. Mehmet. J Dent. Res. Dent. Clin. Dent. Prospects. 2019, 13, 241–246. [Google Scholar] [CrossRef]
- Lim, E.S.; Park, Y.B.; Kwon, Y.S.; Shon, W.J.; Lee, K.W.; Min, K.S. Physical properties and biocompatibility of an injectable calcium-silicate-based root canal sealer: In vitro and in vivo study. BMC Oral Health 2015, 15, 129. [Google Scholar] [CrossRef] [PubMed]
- Cosme-Silva, L.; Dal-Fabbro, R.; Gonçalves, L.O.; Prado, A.S.D.; Plazza, F.A.; Viola, N.V.; Cintra, L.T.A.; Gomes Filho, J.E. Hypertension affects the biocompatibility and biomineralization of MTA, high-plasticity MTA, and Biodentine®. Braz. Oral Res. 2019, 33, e060. [Google Scholar] [CrossRef] [PubMed]
- Cintra, L.T.A.; Benetti, F.; de AzevedoQueiroz, Í.O.; de Araújo Lopes, J.M.; de Oliveira, S.H.P.; SivieriAraújo, G.; Gomes-Filho, J.E. Cytotoxicity, biocompatibility, and biomineralization of the new high-plasticity MTA Material. J. Endod. 2017, 43, 774–778. [Google Scholar] [CrossRef] [PubMed]
- Galarça, A.D.; Da Rosa, W.L.O.; Da Silva, T.M.; da Silveira Lima, G.; Carreño, N.L.V.; Pereira, T.M.; Aguirre Guedes, O.; Borges, A.H.; da Silva, A.F.; Piva, E. Physical and biological properties of a high-plasticity tricalcium silicate cement. Biomed. Res. Int. 2018, 2018, 8063262. [Google Scholar] [CrossRef]
- Mahmood Talabani, R.; Taha Garib, B.; Masaeli, R. The Response of the Pulp-Dentine Complex, PDL, and Bone to Three Calcium Silicate-Based Cements: A histological study in an animal rat model. Bioinorg. Chem. Appl. 2020, 9582165. [Google Scholar] [CrossRef]
- Barczak, K.; Palczewska-Komsa, M.; Nowicka, A.; Chlubek, D.; Buczkowska-Radlińska, J. Analysis of the activity and expression of cyclooxygenases COX1 and COX2 in THP-1 monocytes and macrophages cultured with BiodentineTM silicate cement. Int. J. Mol. Sci. 2020, 21, 2237. [Google Scholar] [CrossRef]
- Qin, Z. The use of THP-1 cells as a model for mimicking the function and regulation of monocytes and macrophages in the vasculature. Atherosclerosis 2012, 221, 2–11. [Google Scholar] [CrossRef]
- Auwerx, J. The human leukemia cell line, THP1: A multifaceted model for study of monocyte-macrophage differentiation. Experientia 1991, 47, 22–31. [Google Scholar] [CrossRef]
- Goppelt-Struebe, M.; Schaefer, D.; Habenicht, A.J.R. Differential regulation of cyclo-oxygenase-2 and 5-lipoxygenase-activating protein (FLAP) expression by glucocorticoids in monocytic cells. Br. J. Pharmacol. 1997, 122, 619–624. [Google Scholar] [CrossRef]
- Olszowski, T.; Baranowska-Bosiacka, I.; Gutowska, I.; Piotrowska, K.; Mierzejewska, K.; Korbecki, J.; Kurzawski, M.; Tarnowski, M.; Chlubek, D. The effects of cadmium at low environmental concentrations on THP-1 macrophage apoptosis. Int. J. Mol. Sci. 2015, 16, 21410–21427. [Google Scholar] [CrossRef]
- Metryka, E.; Kupnicka, P.; Kapczuk, P.; Simińska, D.; Tarnowski, M.; Goschorska, M.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Lead (Pb) as a Factor Initiating and Potentiating Inflammation in Human THP-1 Macrophages. Int. J. Mol. Sci. 2020, 21, 2254. [Google Scholar] [CrossRef] [PubMed]
- Goschorska, M.; Gutowska, I.; Baranowska-Bosiacka, I.; Piotrowska, K.; Metryka, E.; Safranow, K.; Chlubek, D. Influence of acetylcholinesterase inhibitors used in Alzheimer’s Disease treatment on the activity of antioxidant enzymes and the concentration of glutathione in THP-1 macrophages under fluoride-induced oxidative stress. Int. J. Environ. Res. Public Health 2019, 16, 10. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flow Cytometry | ||
|---|---|---|
| CD14 Expression | CD68 Expression | |
| % Positive Cells | ||
| THP1 | 0.8 | 2.9 |
| THP1 + MTA Repair HP | 0.7 | 2.7 |
| THP1 + PMA | 0.1 ns | 45.2 ** |
| THP1 + PMA + MTA Repair HP | 0.1 ns | 43.5 ## |
| Preparation | Producer | Composition |
|---|---|---|
| MTA RepairHP | Angelus, Londrina, Brasil | Powder: Tricalcium Silicate 3CaO·SiO2, Dicalcium Silicate 2CaO·SiO2, Tricalcium Aluminate 3CaO·Al2O3, Calcium Oxide CaO, Calcium Tungstate CAWO4 Liquid: Water and Plasticizer |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barczak, K.; Palczewska-Komsa, M.; Lipski, M.; Chlubek, D.; Buczkowska-Radlińska, J.; Baranowska-Bosiacka, I. The Influence of New Silicate Cement Mineral Trioxide Aggregate (MTA Repair HP) on Metalloproteinase MMP-2 and MMP-9 Expression in Cultured THP-1 Macrophages. Int. J. Mol. Sci. 2021, 22, 295. https://doi.org/10.3390/ijms22010295
Barczak K, Palczewska-Komsa M, Lipski M, Chlubek D, Buczkowska-Radlińska J, Baranowska-Bosiacka I. The Influence of New Silicate Cement Mineral Trioxide Aggregate (MTA Repair HP) on Metalloproteinase MMP-2 and MMP-9 Expression in Cultured THP-1 Macrophages. International Journal of Molecular Sciences. 2021; 22(1):295. https://doi.org/10.3390/ijms22010295
Chicago/Turabian StyleBarczak, Katarzyna, Mirona Palczewska-Komsa, Mariusz Lipski, Dariusz Chlubek, Jadwiga Buczkowska-Radlińska, and Irena Baranowska-Bosiacka. 2021. "The Influence of New Silicate Cement Mineral Trioxide Aggregate (MTA Repair HP) on Metalloproteinase MMP-2 and MMP-9 Expression in Cultured THP-1 Macrophages" International Journal of Molecular Sciences 22, no. 1: 295. https://doi.org/10.3390/ijms22010295
APA StyleBarczak, K., Palczewska-Komsa, M., Lipski, M., Chlubek, D., Buczkowska-Radlińska, J., & Baranowska-Bosiacka, I. (2021). The Influence of New Silicate Cement Mineral Trioxide Aggregate (MTA Repair HP) on Metalloproteinase MMP-2 and MMP-9 Expression in Cultured THP-1 Macrophages. International Journal of Molecular Sciences, 22(1), 295. https://doi.org/10.3390/ijms22010295