GH/IGF-1 Abnormalities and Muscle Impairment: From Basic Research to Clinical Practice



Abstract
:1. Introduction
2. GH/IGF-1 Axis in Humans
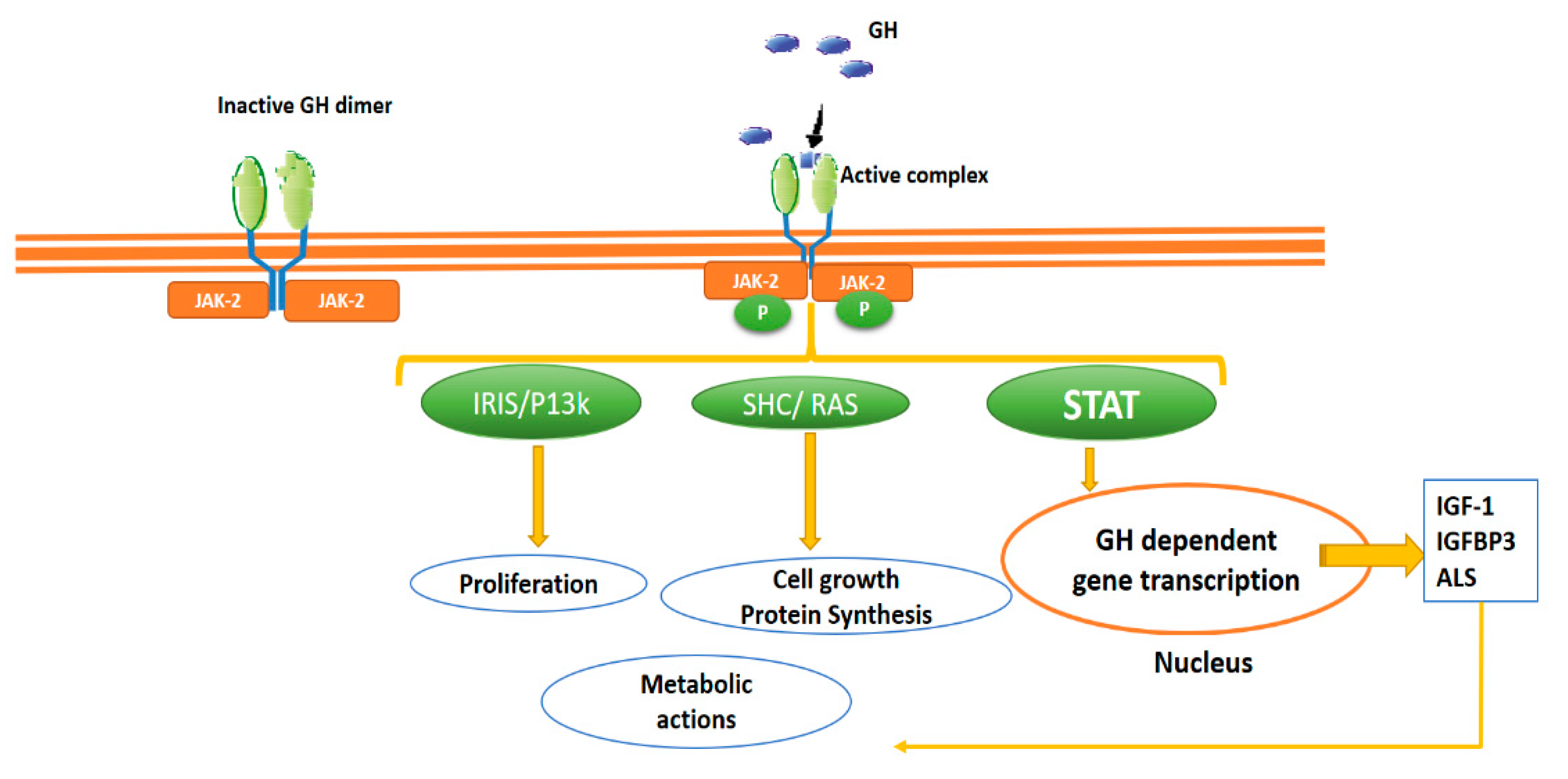
2.1. GH Circulation, GH Receptor and Intracellular Signalling
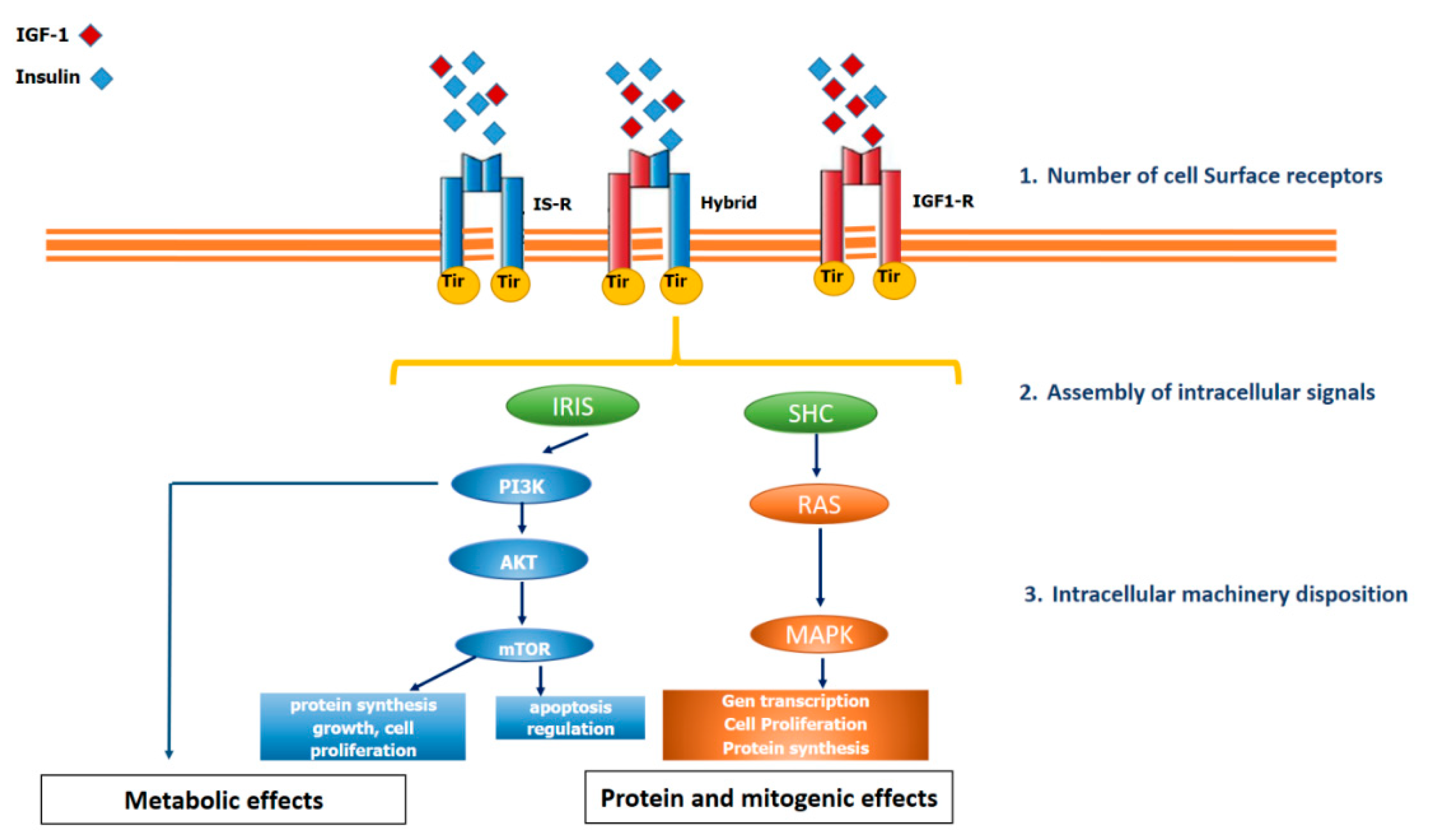
2.2. IGF-1 Circulation, IGF-1 Receptor and Intracellular Signalling
3. The Skeletal Muscle: A Central Link between Structural and Metabolic Function
4. GH/IGF-1 Actions in the Muscular System
4.1. GH
4.2. IGF-1
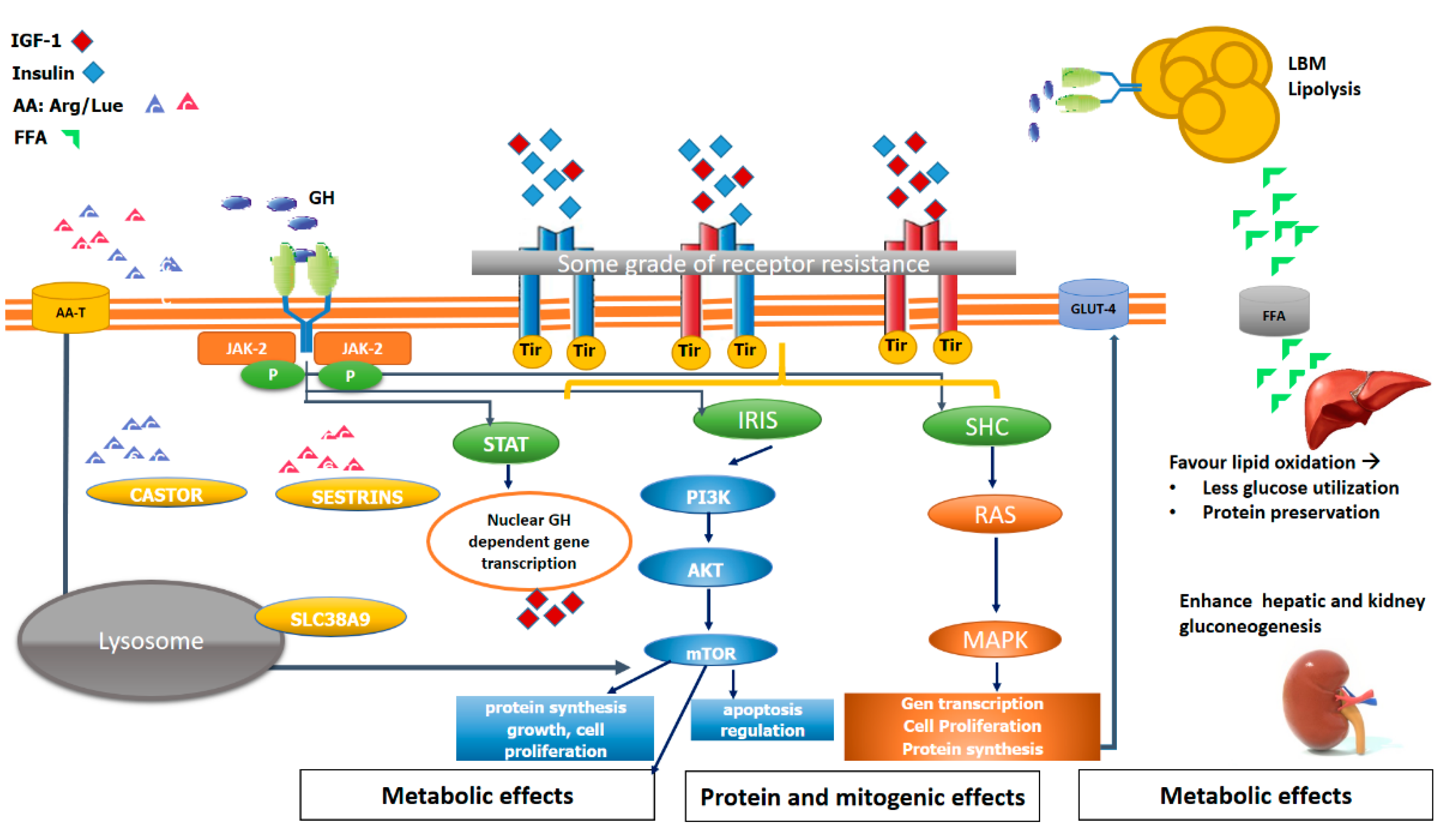
5. The Key of mTOR in Muscle Structural and Metabolic Function
6. Impact of GH/IGF-1 Axis Impairment on Muscle Metabolism and Function: Lessons from Basic Research
6.1. Primary Cell Cultures
6.2. Transgenic Mice
6.3. Experimental Research Focused on the GH/IGF-1 System Effects on Muscle
7. The Muscle in GH/IGF1 Axis Deficiency
8. Muscle Impairment in Human Acromegaly
9. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Colao, A.; Grasso, L.F.S.; Giustina, A.; Melmed, S.; Chanson, P.; Pereira, A.M.; Pivonello, R. Acromegaly. Nat. Rev. Dis. Primers 2019, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Killinger, Z.; Payer, J.; Lazúrová, I.; Imrich, R.; Homérová, Z.; Kužma, M.; Rovenský, J. Arthropathy in acromegaly. Rheum. Dis. Clin. North. Am. 2010, 36, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Melmed, S.; Casanueva, F.F.; Klibanski, A.; Bronstein, M.D.; Chanson, P.; Lamberts, S.W.; Strasburger, C.J.; Wass, J.A.H.; Giustina, A. A consensus on the diagnosis and treatment of acromegaly complications. Pituitary 2013, 16, 294–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassenaar, M.J.E.; Biermasz, N.R.; Kloppenburg, M.; van der Klaauw, A.A.; Tiemensma, J.; Smit, J.W.A.; Pereira, A.M.; Roelfsema, F.; Kroon, H.M.; Romijn, J.A. Clinical osteoarthritis predicts physical and psychological QoL in acromegaly patients. Growth Horm. IGF Res. 2010, 20, 226–233. [Google Scholar] [CrossRef]
- Füchtbauer, L.; Olsson, D.S.; Bengtsson, B.-Å.; Norrman, L.-L.; Sunnerhagen, K.S.; Johannsson, G. Muscle strength in patients with acromegaly at diagnosis and during long-term follow-up. Eur. J. Endocrinol. 2017, 177, 217–226. [Google Scholar] [CrossRef]
- Lima, T.R.L.; Kasuki, L.; Gadelha, M.; Lopes, A.J. Physical exercise improves functional capacity and quality of life in patients with acromegaly: A 12-week follow-up study. Endocrine 2019, 66, 301–309. [Google Scholar] [CrossRef]
- Rajasoorya, C.; Holdaway, I.M.; Wrightson, P.; Scott, D.J.; Ibbertson, H.K. Determinants of clinical outcome and survival in acromegaly. Clin. Endocrinol. 1994, 41, 95–102. [Google Scholar] [CrossRef]
- Orme, S.M.; McNally, R.J.; Cartwright, R.A.; Belchetz, P.E. Mortality and cancer incidence in acromegaly: A retrospective cohort study. United Kingdom Acromegaly Study Group. J. Clin. Endocrinol. Metab. 1998, 83, 2730–2734. [Google Scholar] [CrossRef] [Green Version]
- Holdaway, I.M.; Rajasoorya, R.C.; Gamble, G.D. Factors influencing mortality in acromegaly. J. Clin. Endocrinol. Metab. 2004, 89, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Mercado, M.; Gonzalez, B.; Vargas, G.; Ramirez, C.; de los Monteros, A.L.E.; Sosa, E.; Jervis, P.; Roldan, P.; Mendoza, V.; López-Félix, B.; et al. Successful mortality reduction and control of co-morbidities in patients with acromegaly followed at a highly specialised multidisciplinary clinic. J. Clin. Endocrinol. Metab. 2014, 99, 4438–4446. [Google Scholar] [CrossRef] [Green Version]
- Sardella, C.; Cappellani, D.; Urbani, C.; Manetti, L.; Marconcini, G.; Tomisti, L.; Lupi, I.; Rossi, G.; Scattina, I.; Lombardi, M.; et al. Disease activity and lifestyle influence co-morbidities and cardiovascular events in patients with acromegaly. Eur. J. Endocrinol. 2016, 175, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.M. Quality of life in acromegaly. Neuroendocrinology 2006, 83, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Giustina, A.; Bevan, J.S.; Bronstein, M.D.; Casanueva, F.F.; Chanson, P.; Petersenn, S.; Thanh, X.-M.T.; Sert, C.; Houchard, A.; Guillemin, I.; et al. SAGIT®: Clinician-reported outcome instrument for managing acromegaly in clinical practice—development and results from a pilot study. Pituitary 2016, 19, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Lely, A.J.; Gomez, R.; Pleil, A.; Badia, X.; Brue, T.; Buchfelder, M.; Burman, P.; Clemmons, D.; Ghigo, E.; Jørgensen, J.O.L.; et al. Development of ACRODAT®, a new software medical device to assess disease activity in patients with acromegaly. Pituitary 2017, 20, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Olarescu, N.C.; Gunawardane, K.; Hansen, T.K.; Møller, N.; Jørgensen, J.O.L. Normal Physiology of Growth Hormone in Adults. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dungan, K., Grossman, A., Hershman, J.M., Hofland, H.J., Kaltsas, G., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Waters, M.J. The growth hormone receptor. Growth Horm. IGF Res. 2016, 28, 6–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, M.J.; Hoang, H.N.; Fairlie, D.P.; Pelekanos, R.A.; Brown, R.J. New insights into growth hormone action. J. Mol. Endocrinol. 2006, 36, 1–7. [Google Scholar] [CrossRef]
- O’Sullivan, L.A.; Liongue, C.; Lewis, R.S.; Stephenson, S.E.M.; Ward, A.C. Cytokine receptor signaling through the Jak-Stat-Socs pathway in disease. Mol. Immunol. 2007, 44, 2497–2506. [Google Scholar] [CrossRef]
- Kelly, P.A.; Djiane, J.; Postel-Vinay, M.C.; Edery, M. The prolactin/growth hormone receptor family. Endocr. Rev. 1991, 12, 235–251. [Google Scholar] [CrossRef]
- Kim, J.J.; Accili, D. Signalling through IGF-I and insulin receptors: Where is the specificity? Growth Horm. IGF Res. 2002, 12, 84–90. [Google Scholar] [CrossRef]
- Laviola, L.; Natalicchio, A.; Giorgino, F. The IGF-I signaling pathway. Curr. Pharm. Des. 2007, 13, 663–669. [Google Scholar] [CrossRef]
- Hakuno, F.; Takahashi, S.-I. IGF1 receptor signaling pathways. J. Mol. Endocrinol 2018, 61, T69–T86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dave, H.D.; Shook, M.; Varacallo, M. Anatomy, Skeletal Muscle. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Baskin, K.K.; Winders, B.R.; Olson, E.N. Muscle as a “mediator” of systemic metabolism. Cell Metab 2015, 21, 237–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, N.; Takahashi, H.; Yee, G.M.; Kitajima, Y.; Katagiri, S.; Kojima, M.; Anzai, K.; Eguchi, Y.; Hamilton, J.A. Influence of muscle fiber type composition on early fat accumulation under high-fat diet challenge. PLoS ONE 2017, 12, e0182430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKrell, J.G.; Arias, E.B.; Cartee, G.D. Fiber Type–Specific Differences in Glucose Uptake by Single Fibers From Skeletal Muscles of 9- and 25-Month-Old Rats. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 1286–1294. [Google Scholar] [CrossRef] [Green Version]
- McCuller, C.; Jessu, R.; Callahan, A.L. Physiology, Skeletal Muscle. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Scott, W.; Stevens, J.; Binder–Macleod, S.A. Human Skeletal Muscle Fiber Type Classifications. Phys. Ther. 2001, 81, 1810–1816. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Tripathy, D. Skeletal Muscle Insulin Resistance Is the Primary Defect in Type 2 Diabetes. Diabetes Care 2009, 32, S157–S163. [Google Scholar] [CrossRef] [Green Version]
- Perrini, S.; Laviola, L.; Carreira, M.C.; Cignarelli, A.; Natalicchio, A.; Giorgino, F. The GH/IGF1 axis and signaling pathways in the muscle and bone: Mechanisms underlying age-related skeletal muscle wasting and osteoporosis. J. Endocrinol. 2010, 205, 201–210. [Google Scholar] [CrossRef]
- Møller, N.; Jørgensen, J.O.L. Effects of growth hormone on glucose, lipid, and protein metabolism in human subjects. Endocr. Rev. 2009, 30, 152–177. [Google Scholar] [CrossRef] [Green Version]
- Yakar, S.; Liu, J.-L.; Stannard, B.; Butler, A.; Accili, D.; Sauer, B.; LeRoith, D. Normal growth and development in the absence of hepatic insulin-like growth factor I. Proc. Natl. Acad. Sci. USA 1999, 96, 7324–7329. [Google Scholar] [CrossRef] [Green Version]
- Le Roith, D.; Bondy, C.; Yakar, S.; Liu, J.L.; Butler, A. The somatomedin hypothesis: 2001. Endocr. Rev. 2001, 22, 53–74. [Google Scholar] [CrossRef]
- Bramnert, M.; Segerlantz, M.; Laurila, E.; Daugaard, J.R.; Manhem, P.; Groop, L. Growth hormone replacement therapy induces insulin resistance by activating the glucose-fatty acid cycle. J. Clin. Endocrinol. Metab. 2003, 88, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Szego, C.M.; White, A. The influence of purified growth hormone on fasting metabolism. J. Clin. Endocrinol. Metab. 1948, 8, 594. [Google Scholar] [PubMed]
- Fryburg, D.A.; Barrett, E.J. Growth hormone acutely stimulates skeletal muscle but not whole-body protein synthesis in humans. Metab. Clin. Exp. 1993, 42, 1223–1227. [Google Scholar] [CrossRef]
- Copeland, K.C.; Nair, K.S. Acute growth hormone effects on amino acid and lipid metabolism. J. Clin. Endocrinol. Metab. 1994, 78, 1040–1047. [Google Scholar] [CrossRef]
- Moller, N.; Vendelbo, M.H.; Kampmann, U.; Christensen, B.; Madsen, M.; Norrelund, H.; Jorgensen, J.O. Growth hormone and protein metabolism. Clin. Nutr. 2009, 28, 597–603. [Google Scholar] [CrossRef]
- Fryburg, D.A.; Gelfand, R.A.; Barrett, E.J. Growth hormone acutely stimulates forearm muscle protein synthesis in normal humans. Am. J. Physiol. 1991, 260, E499–E504. [Google Scholar] [CrossRef]
- Fryburg, D.A.; Louard, R.J.; Gerow, K.E.; Gelfand, R.A.; Barrett, E.J. Growth hormone stimulates skeletal muscle protein synthesis and antagonises insulin’s antiproteolytic action in humans. Diabetes 1992, 41, 424–429. [Google Scholar] [CrossRef]
- Buijs, M.M.; Romijn, J.A.; Burggraaf, J.; De Kam, M.L.; Cohen, A.F.; Frölich, M.; Stellaard, F.; Meinders, A.E.; Pijl, H. Growth hormone blunts protein oxidation and promotes protein turnover to a similar extent in abdominally obese and normal-weight women. J. Clin. Endocrinol. Metab. 2002, 87, 5668–5674. [Google Scholar] [CrossRef] [Green Version]
- Gibney, J.; Wolthers, T.; Johannsson, G.; Umpleby, A.M.; Ho, K.K.Y. Growth hormone and testosterone interact positively to enhance protein and energy metabolism in hypopituitary men. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E266–E271. [Google Scholar] [CrossRef] [Green Version]
- Short, K.R.; Moller, N.; Bigelow, M.L.; Coenen-Schimke, J.; Nair, K.S. Enhancement of muscle mitochondrial function by growth hormone. J. Clin. Endocrinol. Metab. 2008, 93, 597–604. [Google Scholar] [CrossRef] [Green Version]
- Yarasheski, K.E.; Campbell, J.A.; Smith, K.; Rennie, M.J.; Holloszy, J.O.; Bier, D.M. Effect of growth hormone and resistance exercise on muscle growth in young men. Am. J. Physiol. 1992, 262, E261–E267. [Google Scholar] [CrossRef] [PubMed]
- Russell-Jones, D.L.; Weissberger, A.J.; Bowes, S.B.; Kelly, J.M.; Thomason, M.; Umpleby, A.M.; Jones, R.H.; Sönksen, P.H. The effects of growth hormone on protein metabolism in adult growth hormone deficient patients. Clin. Endocrinol. 1993, 38, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Jørgensen, J.O.L.; Hartmund, T.; Nørrelund, H.; Nair, K.S.; Christiansen, J.S.; Møller, N. Effects of lowering circulating free fatty acid levels on protein metabolism in adult growth hormone deficient patients. Growth Horm. IGF Res. 2002, 12, 425–433. [Google Scholar] [CrossRef]
- Garibotto, G. Muscle amino acid metabolism and the control of muscle protein turnover in patients with chronic renal failure. Nutrition 1999, 15, 145–155. [Google Scholar] [CrossRef]
- Nørrelund, H.; Nair, K.S.; Jørgensen, J.O.L.; Christiansen, J.S.; Møller, N. The Protein-Retaining Effects of Growth Hormone During Fasting Involve Inhibition of Muscle-Protein Breakdown. Diabetes 2001, 50, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Taaffe, D.R.; Jin, I.H.; Vu, T.H.; Hoffman, A.R.; Marcus, R. Lack of effect of recombinant human growth hormone (GH) on muscle morphology and GH-insulin-like growth factor expression in resistance-trained elderly men. J. Clin. Endocrinol. Metab. 1996, 81, 421–425. [Google Scholar] [CrossRef]
- Taaffe, D.R.; Pruitt, L.; Reim, J.; Hintz, R.L.; Butterfield, G.; Hoffman, A.R.; Marcus, R. Effect of recombinant human growth hormone on the muscle strength response to resistance exercise in elderly men. J. Clin. Endocrinol. Metab. 1994, 79, 1361–1366. [Google Scholar] [CrossRef]
- Anderson, L.J.; Tamayose, J.M.; Garcia, J.M. Use of growth hormone, IGF-I, and insulin for anabolic purpose: Pharmacological basis, methods of detection, and adverse effects. Mol. Cell Endocrinol. 2018, 464, 65–74. [Google Scholar] [CrossRef]
- Mendias, C.L.; Enselman, E.R.S.; Olszewski, A.M.; Gumucio, J.P.; Edon, D.L.; Konnaris, M.A.; Carpenter, J.E.; Awan, T.M.; Jacobson, J.A.; Gagnier, J.J.; et al. The Use of Recombinant Human Growth Hormone to Protect Against Muscle Weakness in Patients Undergoing Anterior Cruciate Ligament Reconstruction: A Pilot, Randomized Placebo-Controlled Trial. Am. J. Sports Med. 2020, 48, 1916–1928. [Google Scholar] [CrossRef]
- Goldspink, G. Loss of muscle strength during aging studied at the gene level. Rejuvenation Res. 2007, 10, 397–405. [Google Scholar] [CrossRef]
- Yoshida, T.; Delafontaine, P. Mechanisms of IGF-1-Mediated Regulation of Skeletal Muscle Hypertrophy and Atrophy. Cells 2020, 9, 1970. [Google Scholar] [CrossRef] [PubMed]
- Majorczyk, M.; Smoląg, D. Effect of physical activity on IGF-1 and IGFBP levels in the context of civilisation diseases prevention. Rocz Panstw Zakl Hig 2016, 67, 105–111. [Google Scholar] [PubMed]
- Kruk, J. Physical activity in the prevention of the most frequent chronic diseases: An analysis of the recent evidence. Asian Pac. J. Cancer Prev. 2007, 8, 325–338. [Google Scholar] [PubMed]
- Daughaday, W.H.; Hall, K.; Raben, M.S.; Salmon, W.D.; van den Brande, J.L.; van Wyk, J.J. Somatomedin: Proposed designation for sulphation factor. Nature 1972, 235, 107. [Google Scholar] [CrossRef] [PubMed]
- DeVol, D.L.; Rotwein, P.; Sadow, J.L.; Novakofski, J.; Bechtel, P.J. Activation of insulin-like growth factor gene expression during work-induced skeletal muscle growth. Am. J. Physiol. 1990, 259, E89–E95. [Google Scholar] [CrossRef]
- Niwa, M.; Sato, S.; Saito, Y.; Uchiyama, F.; Ono, H.; Yamashita, M.; Kitaguchi, T.; Shiga, Y.; Notani, J.; Yamada, H. Chemical synthesis, cloning, and expression of genes for human somatomedin C (insulin-like growth factor I) and 59Val-somatomedin C. Ann. N Y Acad. Sci. 1986, 469, 31–52. [Google Scholar] [CrossRef]
- Guler, H.P.; Zapf, J.; Froesch, E.R. Short-term metabolic effects of recombinant human insulin-like growth factor I in healthy adults. N Engl. J. Med. 1987, 317, 137–140. [Google Scholar] [CrossRef]
- Laron, Z.; Klinger, B.; Silbergeld, A.; Lewin, R.; Erster, B.; Gil-Ad, I. Intravenous administration of recombinant IGF-I lowers serum GHRH and TSH. Acta Endocrinol. 1990, 123, 378–382. [Google Scholar] [CrossRef]
- Klinger, B.; Garty, M.; Laron, Z. Elimination Characteristics of Intravenously Administered rIGF-I in Laron-Type Dwarfs. DPD 1990, 15, 196–199. [Google Scholar] [CrossRef]
- Laron, Z. Insulin-like growth factor 1 (IGF-1): A growth hormone. Mol. Pathol. 2001, 54, 311–316. [Google Scholar] [CrossRef]
- Laron, Z.; Klinger, B.; Jensen, L.T.; Erster, B. Biochemical and hormonal changes induced by one week of administration of rIGF-I to patients with Laron type dwarfism. Clin. Endocrinol. 1991, 35, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, D.R.; Smith-Banks, A.; Underwood, L.E. Reversal of diet-induced catabolism by infusion of recombinant insulin-like growth factor-I in humans. J. Clin. Endocrinol. Metab. 1992, 75, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Turkalj, I.; Keller, U.; Ninnis, R.; Vosmeer, S.; Stauffacher, W. Effect of increasing doses of recombinant human insulin-like growth factor-I on glucose, lipid, and leucine metabolism in man. J. Clin. Endocrinol. Metab. 1992, 75, 1186–1191. [Google Scholar] [CrossRef] [PubMed]
- Russell-Jones, D.L.; Umpleby, A.M.; Hennessy, T.R.; Bowes, S.B.; Shojaee-Moradie, F.; Hopkins, K.D.; Jackson, N.C.; Kelly, J.M.; Jones, R.H.; Sönksen, P.H. Use of a leucine clamp to demonstrate that IGF-I actively stimulates protein synthesis in normal humans. Am. J. Physiol. 1994, 267, E591–E598. [Google Scholar] [CrossRef]
- Kupfer, S.R.; Underwood, L.E.; Baxter, R.C.; Clemmons, D.R. Enhancement of the anabolic effects of growth hormone and insulin-like growth factor I by use of both agents simultaneously. J. Clin. Investig. 1993, 91, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Laager, R.; Ninnis, R.; Keller, U. Comparison of the effects of recombinant human insulin-like growth factor-I and insulin on glucose and leucine kinetics in humans. J. Clin. Investig. 1993, 92, 1903–1909. [Google Scholar] [CrossRef] [Green Version]
- Rutter, M.M.; Wong, B.L.; Collins, J.J.; Sawnani, H.; Taylor, M.D.; Horn, P.S.; Backeljauw, P.F. Recombinant human insulin-like growth factor-1 therapy for 6 months improves growth but not motor function in boys with Duchenne muscular dystrophy. Muscle Nerve 2020, 61, 623–631. [Google Scholar] [CrossRef]
- Guha, N.; Nevitt, S.P.; Francis, M.; Woodland, J.A.; Böhning, D.; Sönksen, P.H.; Holt, R.I.G. The Effects of Recombinant Human Insulin-Like Growth Factor-I/Insulin-Like Growth Factor Binding Protein-3 Administration on Body Composition and Physical Fitness in Recreational Athletes. J. Clin. Endocrinol. Metab. 2015, 100, 3126–3131. [Google Scholar] [CrossRef] [Green Version]
- Barclay, R.D.; Burd, N.A.; Tyler, C.; Tillin, N.A.; Mackenzie, R.W. The Role of the IGF-1 Signaling Cascade in Muscle Protein Synthesis and Anabolic Resistance in Aging Skeletal Muscle. Front. Nutr. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Janssen, J.A.M.J.L. Mechanisms of putative IGF-I receptor resistance in active acromegaly. Growth Horm. IGF Res. 2020, 52, 101319. [Google Scholar] [CrossRef]
- Sabatini, D.M. Twenty-five years of mTOR: Uncovering the link from nutrients to growth. Proc. Natl. Acad. Sci. USA 2017, 114, 11818–11825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laplante, M.; Sabatini, D.M. mTOR Signaling in Growth Control and Disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moro, T.; Ebert, S.M.; Adams, C.M.; Rasmussen, B.B. Amino Acid Sensing in Skeletal Muscle. Trends Endocrinol. Metab. 2016, 27, 796–806. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Tsun, Z.-Y.; Wolfson, R.L.; Shen, K.; Wyant, G.A.; Plovanich, M.E.; Yuan, E.D.; Jones, T.D.; Chantranupong, L.; Comb, W.; et al. Lysosomal amino acid transporter SLC38A9 signals arginine sufficiency to mTORC1. Science 2015, 347, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfson, R.L.; Chantranupong, L.; Saxton, R.A.; Shen, K.; Scaria, S.M.; Cantor, J.R.; Sabatini, D.M. Sestrin2 is a leucine sensor for the mTORC1 pathway. Science 2016, 351, 43–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxton, R.A.; Chantranupong, L.; Knockenhauer, K.E.; Schwartz, T.U.; Sabatini, D.M. Mechanism of arginine sensing by CASTOR1 upstream of mTORC1. Nature 2016, 536, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Biagetti, B.; Herance, J.R.; Ferrer, R.; Aulinas, A.; Palomino-Schätzlein, M.; Mesa, J.; Castaño, J.P.; Luque, R.M.; Simó, R. Metabolic Fingerprint of Acromegaly and its Potential Usefulness in Clinical Practice. J. Clin. Med. 2019, 8, 1549. [Google Scholar] [CrossRef] [Green Version]
- Chikani, V.; Ho, K.K.Y. Action of GH on skeletal muscle function: Molecular and metabolic mechanisms. J. Mol. Endocrinol. 2014, 52, R107–R123. [Google Scholar] [CrossRef] [Green Version]
- Bartke, A. Growth hormone and aging. Rev. Endocr. Metab. Disord 2020. [Google Scholar] [CrossRef]
- Hembree, J.R.; Hathaway, M.R.; Dayton, W.R. Isolation and culture of fetal porcine myogenic cells and the effect of insulin, IGF-I, and sera on protein turnover in porcine myotube cultures. J. Anim. Sci. 1991, 69, 3241–3250. [Google Scholar] [CrossRef]
- Foulstone, E.J.; Huser, C.; Crown, A.L.; Holly, J.M.P.; Stewart, C.E.H. Differential signalling mechanisms predisposing primary human skeletal muscle cells to altered proliferation and differentiation: Roles of IGF-I and TNFalpha. Exp. Cell Res. 2004, 294, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Sawa, R.; Nishida, H.; Yamamoto, Y.; Wake, I.; Kai, N.; Kikkawa, U.; Okimura, Y. Growth hormone and Insulin-like growth factor-I (IGF-I) modulate the expression of L-type amino acid transporters in the muscles of spontaneous dwarf rats and L6 and C2C12 myocytes. Growth Horm. IGF Res. 2018, 42–43, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Mavalli, M.D.; DiGirolamo, D.J.; Fan, Y.; Riddle, R.C.; Campbell, K.S.; van Groen, T.; Frank, S.J.; Sperling, M.A.; Esser, K.A.; Bamman, M.M.; et al. Distinct growth hormone receptor signaling modes regulate skeletal muscle development and insulin sensitivity in mice. J. Clin. Investig. 2010, 120, 4007–4020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- List, E.O.; Basu, R.; Duran-Ortiz, S.; Krejsa, J.; Jensen, E.A. Mouse models of growth hormone deficiency. Rev. Endocr Metab Disord 2020. [Google Scholar] [CrossRef] [PubMed]
- Gahete, M.D.; Luque, R.M.; Castaño, J.P. Models of GH deficiency in animal studies. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Brinster, R.L.; Hammer, R.E.; Trumbauer, M.E.; Rosenfeld, M.G.; Birnberg, N.C.; Evans, R.M. Dramatic growth of mice that develop from eggs microinjected with metallothionein-growth hormone fusion genes. Nature 1982, 300, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Luque, R.M. Bidirectional Interrelationship between Growth Hormone and Metabolism; BioScientifica: Bristol, UK, 2019. [Google Scholar]
- Carter, C.S.; Ramsey, M.M.; Ingram, R.L.; Cashion, A.B.; Cefalu, W.T.; Wang, Z.Q.; Sonntag, W.E. Models of growth hormone and IGF-1 deficiency: Applications to studies of aging processes and life-span determination. J. Gerontol. A Biol. Sci. Med. Sci. 2002, 57, B177–B188. [Google Scholar] [CrossRef]
- Kopchick, J.J.; List, E.O.; Kelder, B.; Gosney, E.S.; Berryman, D.E. Evaluation of growth hormone (GH) action in mice: Discovery of GH receptor antagonists and clinical indications. Mol. Cell Endocrinol. 2014, 386, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Dal, J.; List, E.O.; Jørgensen, J.O.L.; Berryman, D.E. Glucose and Fat Metabolism in Acromegaly: From Mice Models to Patient Care. Neuroendocrinology 2016, 103, 96–105. [Google Scholar] [CrossRef]
- Liu, J.P.; Baker, J.; Perkins, A.S.; Robertson, E.J.; Efstratiadis, A. Mice carrying null mutations of the genes encoding insulin-like growth factor I (Igf-1) and type 1 IGF receptor (Igf1r). Cell 1993, 75, 59–72. [Google Scholar] [CrossRef]
- Li, S.; Crenshaw, E.B.; Rawson, E.J.; Simmons, D.M.; Swanson, L.W.; Rosenfeld, M.G. Dwarf locus mutants lacking three pituitary cell types result from mutations in the POU-domain gene pit-1. Nature 1990, 347, 528–533. [Google Scholar] [CrossRef]
- Ayling, C.M.; Moreland, B.H.; Zanelli, J.M.; Schulster, D. Human growth hormone treatment of hypophysectomised rats increases the proportion of type-1 fibres in skeletal muscle. J. Endocrinol. 1989, 123, 429–435. [Google Scholar] [CrossRef]
- Loughna, P.T.; Bates, P.C. Interactions between growth hormone and nutrition in hypophysectomised rats: Skeletal muscle myosin heavy chain mRNA levels. Biochem. Biophys. Res. Commun. 1994, 198, 97–102. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Sakuma, K.; Morita, I.; Soya, H.; Takeda, H.; Katsuta, S. Changes in fibre types in rat soleus and plantaris muscles following hypophysectomy and compensatory overload. Acta Physiol. Scand. 1996, 158, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Everitt, A.V.; Terry, V.; Phillips, M.J.; Kerry, H.M.; Shorey, C.D. Morphometric analysis of gastrocnemius muscle fiber size and fiber proportions in the hypophysectomised rat after prolonged administration of growth hormone or thyroxine. Growth Dev. Aging 1996, 60, 85–93. [Google Scholar] [PubMed]
- Nielsen, R.H.; Clausen, N.M.; Schjerling, P.; Larsen, J.O.; Martinussen, T.; List, E.O.; Kopchick, J.J.; Kjaer, M.; Heinemeier, K.M. Chronic alterations in growth hormone/insulin-like growth factor-I signaling lead to changes in mouse tendon structure. Matrix Biol. 2014, 34, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cianforlini, M.; Grassi, M.; Coppa, V.; Manzotti, S.; Orlando, F.; Mattioli-Belmonte, M.; Gigante, A. Skeletal muscle repair in a rat muscle injury model: The role of growth hormone (GH) injection. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8566–8572. [Google Scholar] [CrossRef]
- Schoenle, E.; Zapf, J.; Humbel, R.E.; Froesch, E.R. Insulin-like growth factor I stimulates growth in hypophysectomised rats. Nature 1982, 296, 252–253. [Google Scholar] [CrossRef]
- Alatzoglou, K.S.; Webb, E.A.; Le Tissier, P.; Dattani, M.T. Isolated growth hormone deficiency (GHD) in childhood and adolescence: Recent advances. Endocr. Rev. 2014, 35, 376–432. [Google Scholar] [CrossRef]
- Nadler, H.L.; Neumann, L.L.; Gershberg, H. HYPOGLYCEMIA, GROWTH RETARDATION, AND PROBABLE ISOLATED GROWTH HORMONE DEFICIENCY IN A 1-YEAR-OLD CHILD. J. Pediatr. 1963, 63, 977–983. [Google Scholar] [CrossRef]
- Boguszewski, M.C.S. Growth hormone deficiency and replacement in children. Rev. Endocr. Metab. Disord 2020. [Google Scholar] [CrossRef] [PubMed]
- Boguszewski, C.L. Individual sensitivity to growth hormone replacement in adults. Rev. Endocr. Metab. Disord 2020. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, J.O.L.; Juul, A. THERAPY OF ENDOCRINE DISEASE: Growth hormone replacement therapy in adults: 30 years of personal clinical experience. Eur J. Endocrinol. 2018, 179, R47–R56. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Guimarães, A.L.; Aguiar-Oliveira, M.H.; Salvatori, R.; Carvalho, V.O.; Alvim-Pereira, F.; Daniel, C.R.A.; Brasileiro, G.A.M.; Santana-Ribeiro, A.A.; Santos-Carvalho, H.A.; Oliveira, C.R.P.; et al. Adult individuals with congenital, untreated, severe isolated growth hormone deficiency have satisfactory muscular function. Endocrine 2019, 63, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Díez, J.J.; Sangiao-Alvarellos, S.; Cordido, F. Treatment with Growth Hormone for Adults with Growth Hormone Deficiency Syndrome: Benefits and Risks. Int. J. Mol. Sci. 2018, 19, 893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Barkan, A.L. Growth hormone therapy in adults with growth hormone deficiency: A critical assessment of the literature. Pituitary 2020, 23, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Laron, Z.; Kauli, R.; Lapkina, L.; Werner, H. IGF-I deficiency, longevity and cancer protection of patients with Laron syndrome. Mutat. Res. Rev. Mutat. Res. 2017, 772, 123–133. [Google Scholar] [CrossRef]
- Guevara-Aguirre, J.; Rosenbloom, A.L. Obesity, diabetes and cancer: Insight into the relationship from a cohort with growth hormone receptor deficiency. Diabetologia 2015, 58, 37–42. [Google Scholar] [CrossRef]
- Laron, Z.; Kauli, R. Fifty seven years of follow-up of the Israeli cohort of Laron Syndrome patients-From discovery to treatment. Growth Horm IGF Res. 2016, 28, 53–56. [Google Scholar] [CrossRef]
- Laron, Z. LESSONS FROM 50 YEARS OF STUDY OF LARON SYNDROME. Endocr. Pract. 2015, 21, 1395–1402. [Google Scholar] [CrossRef]
- Laron, Z. Insulin-like growth factor-I treatment of children with Laron syndrome (primary growth hormone insensitivity). Pediatr. Endocrinol. Rev. 2008, 5, 766–771. [Google Scholar] [PubMed]
- Miller, A.; Doll, H.; David, J.; Wass, J. Impact of musculoskeletal disease on quality of life in long-standing acromegaly. Eur. J. Endocrinol. 2008, 158, 587–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadelha, M.R.; Kasuki, L.; Lim, D.S.T.; Fleseriu, M. Systemic Complications of Acromegaly and the Impact of the Current Treatment Landscape: An Update. Endocr. Rev. 2019, 40, 268–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodmansee, W.W.; Gordon, M.B.; Molitch, M.E.; Ioachimescu, A.G.; Carver, D.W.; Mirakhur, B.; Cox, D.; Salvatori, R. Screening for comorbid conditions in patients enrolled in the SODA registry: A 2-year observational analysis. Endocrine 2018, 61, 105–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastaglia, F.L.; Barwick, D.D.; Hall, R. MYOPATHY IN ACROMEGALY. Lancet 1970, 296, 907–909. [Google Scholar] [CrossRef]
- Nagulesparen, M.; Trickey, R.; Davies, M.J.; Jenkins, J.S. Muscle changes in acromegaly. BMJ 1976, 2, 914–915. [Google Scholar] [CrossRef] [Green Version]
- Szendroedi, J.; Zwettler, E.; Schmid, A.I.; Chmelik, M.; Pacini, G.; Kacerovsky, G.; Smekal, G.; Nowotny, P.; Wagner, O.; Schnack, C.; et al. Reduced basal ATP synthetic flux of skeletal muscle in patients with previous acromegaly. PLoS ONE 2008, 3, e3958. [Google Scholar] [CrossRef]
- Elbialy, A.; Asakawa, S.; Watabe, S.; Kinoshita, S. A Zebrafish Acromegaly Model Elevates DNA Damage and Impairs DNA Repair Pathways. Biology 2018, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Brummer, R.-J.M.; Lönns, L.; Kvist, H.; Grangård, U.; Bengtsson, B.-A.; Sjöström, L. Adipose tissue and muscle volume determination by computed tomography in acromegaly, before and 1 year after adenomectomy. Eur. J. Clin. Investig. 1993, 23, 199–205. [Google Scholar] [CrossRef]
- Freda, P.U.; Shen, W.; Reyes-Vidal, C.M.; Geer, E.B.; Arias-Mendoza, F.; Gallagher, D.; Heymsfield, S.B. Skeletal muscle mass in acromegaly assessed by magnetic resonance imaging and dual-photon x-ray absorptiometry. J. Clin. Endocrinol. Metab. 2009, 94, 2880–2886. [Google Scholar] [CrossRef]
- Ozturk Gokce, B.; Gogus, F.; Bolayir, B.; Tecer, D.; Gokce, O.; Eroglu Altinova, A.; Balos Toruner, F.; Akturk, M. The evaluation of the tendon and muscle changes of lower extremity in patients with acromegaly. Pituitary 2020, 23, 338–346. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author | Subjects | Design | GH Dose | Effects | Conclusions |
|---|---|---|---|---|---|
| Fryberg [39] | 7 healthy | Brachial artery infusion no placebo | 0.014 μg/kg/min To rise locally not systemic | Rd PHe  BCAAs  release | Locally infused GH stimulates skeletal muscle protein synthesis. |
| Fryberg [40] | 7 healthy | Brachial artery infusion 3 h GH and then 3 h GH + insulin/no placebo | GH 0.014 μg/kg/min Insulin 0.02 mU/kg/min To rise locally not systemic | 3 h Rd 6 h Rd Ra  BCAAs | GH blunted the action of insulin to suppress proteolysis |
| Yarasheski [44] | 18 healthy man | Resistance training GH/placebo | GH 40 μg/d for 12 weeks | protein synthesis Leu. Ox | Similar muscle size, strength and protein synthesis |
| Fryburt and Barret [36] | 8 healthy | Systemic brachial GH infusion/no placebo | 0.06 μg/kg/min for 6 h systemic IGF-1 | Rd Ra | Acute stimulation of muscle but not whole-body protein synthesis |
| Russell-Jones [45] | 18 adults GHD | Double-blind, placebo-controlled trial | 0.018 IU/kg/day for 1 month followed by 0.036 IU/kg/day for 1 month | Leu Ra in either the placebo or GH-treated In GH group Leu Rd | GH action in adults with GHD is due to an increase in protein synthesis. |
| Copeland [37] | 15 healthy | Infusion/SSA +/− GH/control | 2 μg/kg/h for 3.5 h | Ra leu Ox | Acute GH did not affect muscle protein synthesis despite enhanced protein synthesis in non-muscle tissue |
| Garibotto [47] | 6 Dialysis | Prospective cross over trial 6-week run and 6-week washout/no control | 5 mg 3 times a week for 6 weeks. | Rd BCAAs | Increased muscle protein synthesis and decreased negative muscle protein balance. |
| Norrelund [48] | 8 healthy basal or after 40 h fasting | Systemic infusion/pancreatic clamp/controlled | 4.5 IU (1) basal (2) after 40 h of fasting (3) after 40 h of fasting + SSAs (4) after 40 h of fasting with SSAs + GH replacement. | GH suppression Ra ----------- GH replacement Ra BCAAs | Suppression of GH during fasting leads to a 50% increase in urea-nitrogen excretion and increased proteolysis. |
| Nielsen [46] | 7 GHD | Subcutaneous GH/controlled | GH replacement +/− acipimox | Ra Rd | Lowering FFA with Acipimox increased whole body and forearm protein breakdown, and protein synthesis |
| Short [43] | 9 healthy | Randomised crossover design GH/FS | GH (150 g/h; 2,1 ± 0.1 g/kg h) | Ra Rd | After GH synthetic rate of muscle proteins |
| Buijs [41] | 6 NW 6 OB | Crossover design placebo | A 1-h iv infusion of SSAs and GH (12 mU/kg/h or placebo | Leu Ox | Administration of GH blunted the rise in Leu Ox similarly in both NW and OB |
| Gibney [42] | 12 hypopit. man | Open-label randomised crossover testosterone and GH | 0.5 mg daily 6 weeks | No effect in protein breakdown, suppressing protein ox and stimulating protein synthesis |
Decrease Increase, not change. Ins: insulin, Leu: leucine, Ox: oxidation, BCAAs: branched-chain amino acids, Phe: phenylalanine, NW: normal weight, OB: obese, wk week. Units: Kg: kilogram, h: hours, IU: international units, min: minutes, mg: milligram, mU: primary units, μg: microgram. Rd: (Phe/Leu disappearance rate) represents protein synthesis. Ra: (rate of appearance of Phe/Leu) represents proteolysis. Hypopit: hypopituitarism. Ox—oxidation.| Author | Subjects | Design | Dose | Muscle Anabolism | Muscle Function | Conclusions |
|---|---|---|---|---|---|---|
| Clemmons [65] | 6 healthy | 2 weeks calorically restricted | rhGH: 0.05 mg/kg *6 d or iv infusion of rhIGF-I 12 μg/kg IBW *16 h | With IGF-1 serum urea nitrogen the reduction was greater than with GH | NA | IGF-1 reversed the catabolism caused by a restriction of the diet. |
| Turkalj [66] | 19 healthy man | Acute randomised rhIGF-1 ascending doses vs. saline. | IGF-I doses 5, 7.5, 15 and 30 μg/kg/h (n = 4) and saline control (n = 3). | Dose-dependent decrease of leucine oxidation. | NA | GF-1 proteolysis IGF-1 protein synthesis |
| Russell-Jones [67] | 5 healthy adequate substrate supply condition), | Random order of both IGF-1 or insulin 1 week apart + AA infusion | 3-h IV infusion of IGF-1: 43.7 pmol*kg/min or Insulin 3.4 pmol/kg/min | IGF-1RaRd Insulin RaRd | NA | IGF-1 increases protein synthesis in contrast to insulin, which acts to reduce proteolysis. |
| Kupfer [68] | 7 healthy | 2 weeks calorically restricted (20 kcal/kg IBW per d), with 1 g protein/kg IBW. | IV infusion of rhIGF-1 12 μg/kg IBW *16 h *5 d or rhIGF-1 (same doses above) + rhGH: 0.05 mg/kg *5 d | Nitrogen retention was 2.4-fold greater in combination | NA | Combination of GH and IGF-I was substantially more anabolic than either GH or IGF-I alone. |
| Lagger [69] | 24 healthy male | Randomised and paired three groups IGF-1-insulin | High doses (30 μg/kg/h IGF-I or 0.23 nmol/kg/h insulin); low doses (5 μg/kg/h IGF-I or 0.04 nmol/kg/h insulin) | RaRd ------------ Any change compared to control | NA | High doses of IGF-1 compared to insulin decreased more proteolysis. Unexpected inhibition of protein synthesis |
| Rutter [70] | Boys with DMD 21 control 17 IGF1 | Randomised, rhIGF-1 vs. placebo. 6 month | GC-treated + IGF-1 (n = 17) vs. controls (GC-therapy only n = 21) | NA | Did not observe a change in functional motor outcomes | Boys with DMD. 6 months of rhIGF-1 therapy did not change motor function but did improve linear growth. |
| Nishan Guha [71] | Fifty-six recreational athletes | Randomised, double-blind, placebo-controlled | Low dose rhIGF-I/rhIGFBP-3 (30 mg/d), or high dose rhIGF-I/rhIGFBP-3 (60 mg/d) for 28 d | NA | Significant increase in maximal oxygen consumption | rhIGF-I/rhIGFBP-3 administration for 28 days improved aerobic performance in recreational athletes, with no effects on body composition. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biagetti, B.; Simó, R. GH/IGF-1 Abnormalities and Muscle Impairment: From Basic Research to Clinical Practice. Int. J. Mol. Sci. 2021, 22, 415. https://doi.org/10.3390/ijms22010415
Biagetti B, Simó R. GH/IGF-1 Abnormalities and Muscle Impairment: From Basic Research to Clinical Practice. International Journal of Molecular Sciences. 2021; 22(1):415. https://doi.org/10.3390/ijms22010415
Chicago/Turabian StyleBiagetti, Betina, and Rafael Simó. 2021. "GH/IGF-1 Abnormalities and Muscle Impairment: From Basic Research to Clinical Practice" International Journal of Molecular Sciences 22, no. 1: 415. https://doi.org/10.3390/ijms22010415
APA StyleBiagetti, B., & Simó, R. (2021). GH/IGF-1 Abnormalities and Muscle Impairment: From Basic Research to Clinical Practice. International Journal of Molecular Sciences, 22(1), 415. https://doi.org/10.3390/ijms22010415