
Double p52Shc/p46Shc Rat Knockout Demonstrates Severe Gait Abnormalities Accompanied by Dilated Cardiomyopathy



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
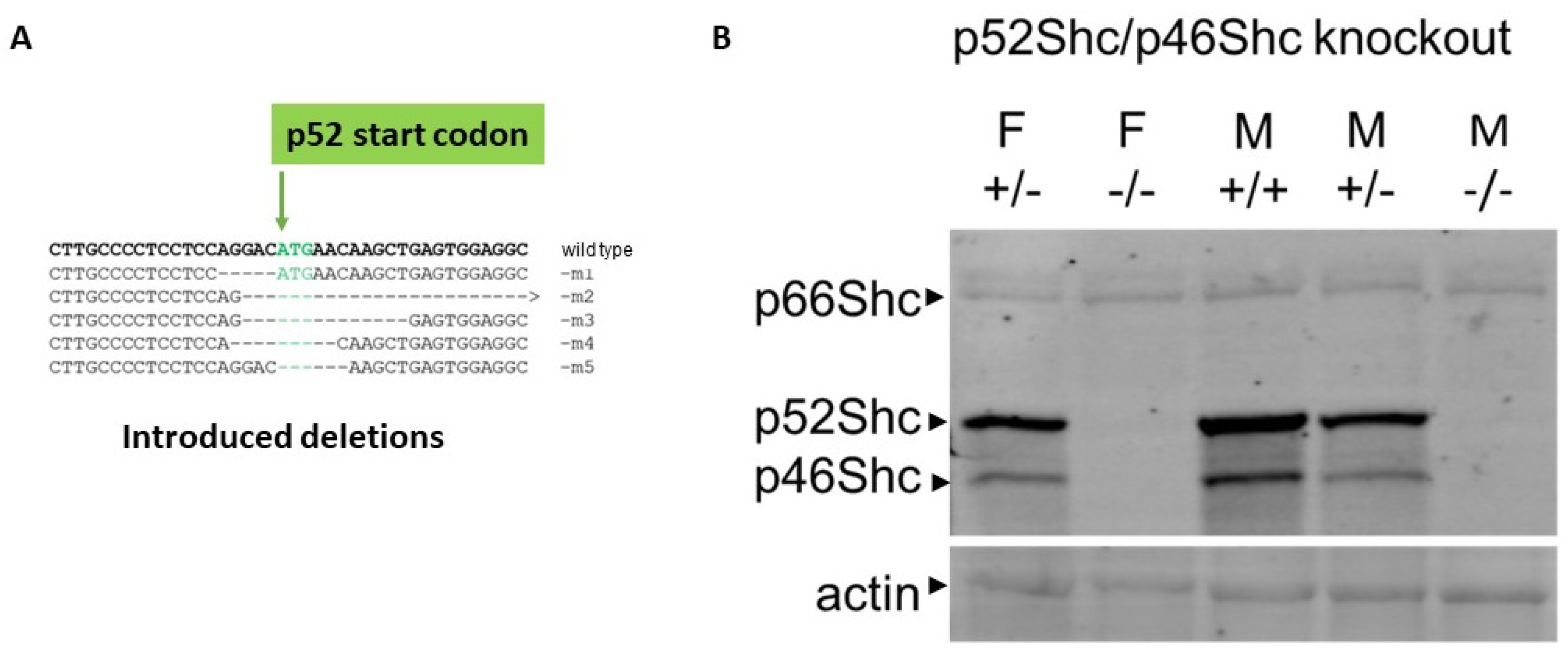
2.1. Generation of Rat Strains Deficient in Shc Isoforms
2.2. p52Shc/p46Shc-KO Rats Demonstrate Severe Gait Abnormalities
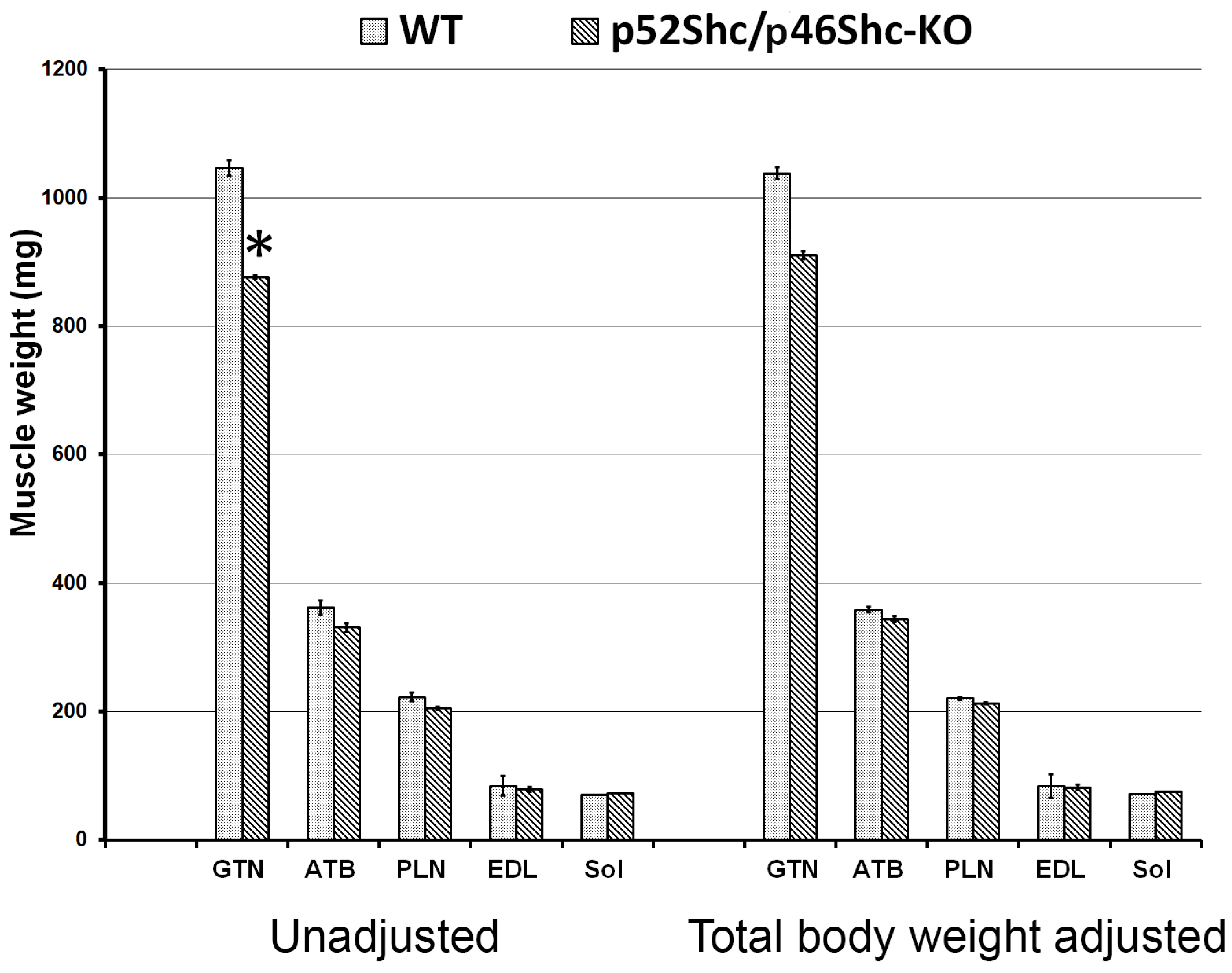
2.3. Total Body and Muscle Mass of p52Shc/p46Shc-KO Rats
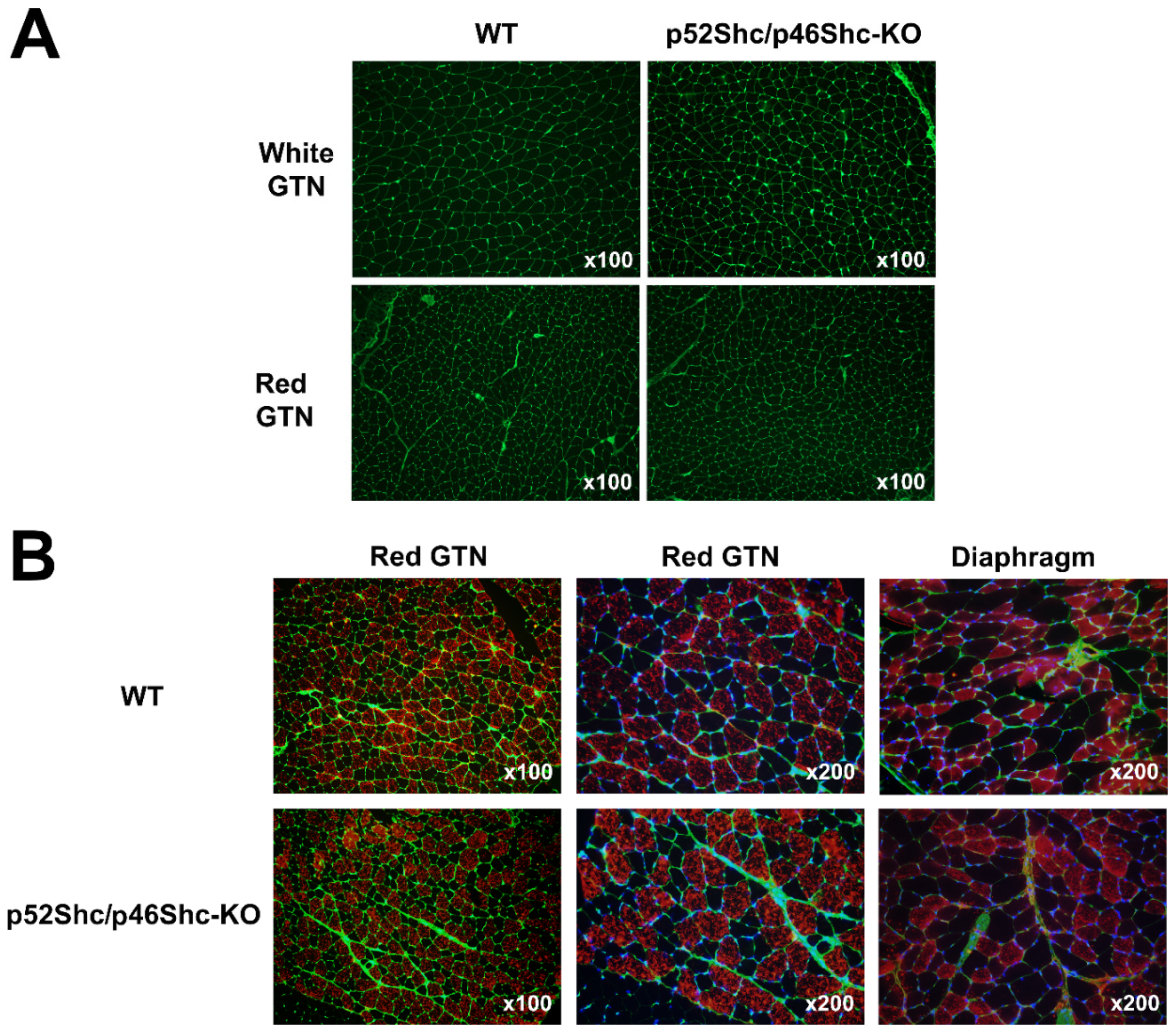
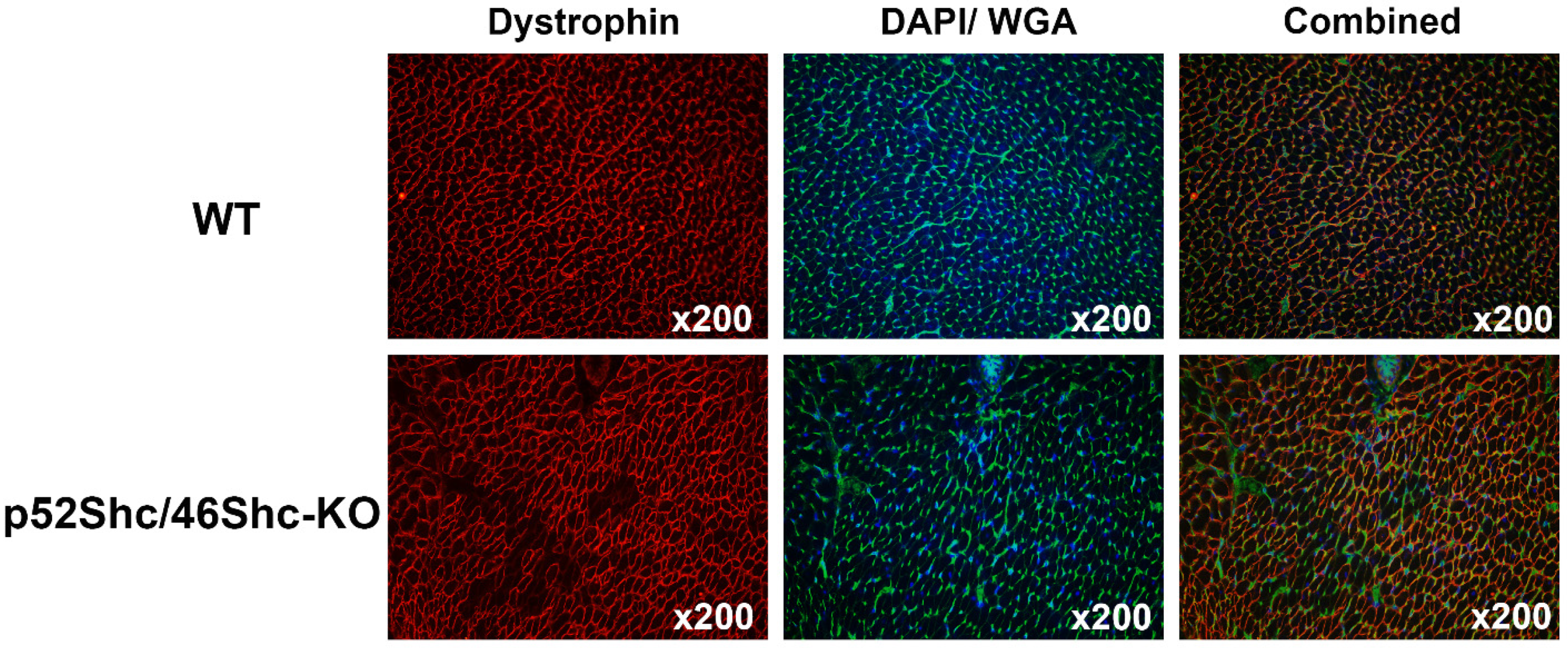
2.4. Histological Analysis of Skeletal Muscle of p52Shc/46Shc-KO Rats
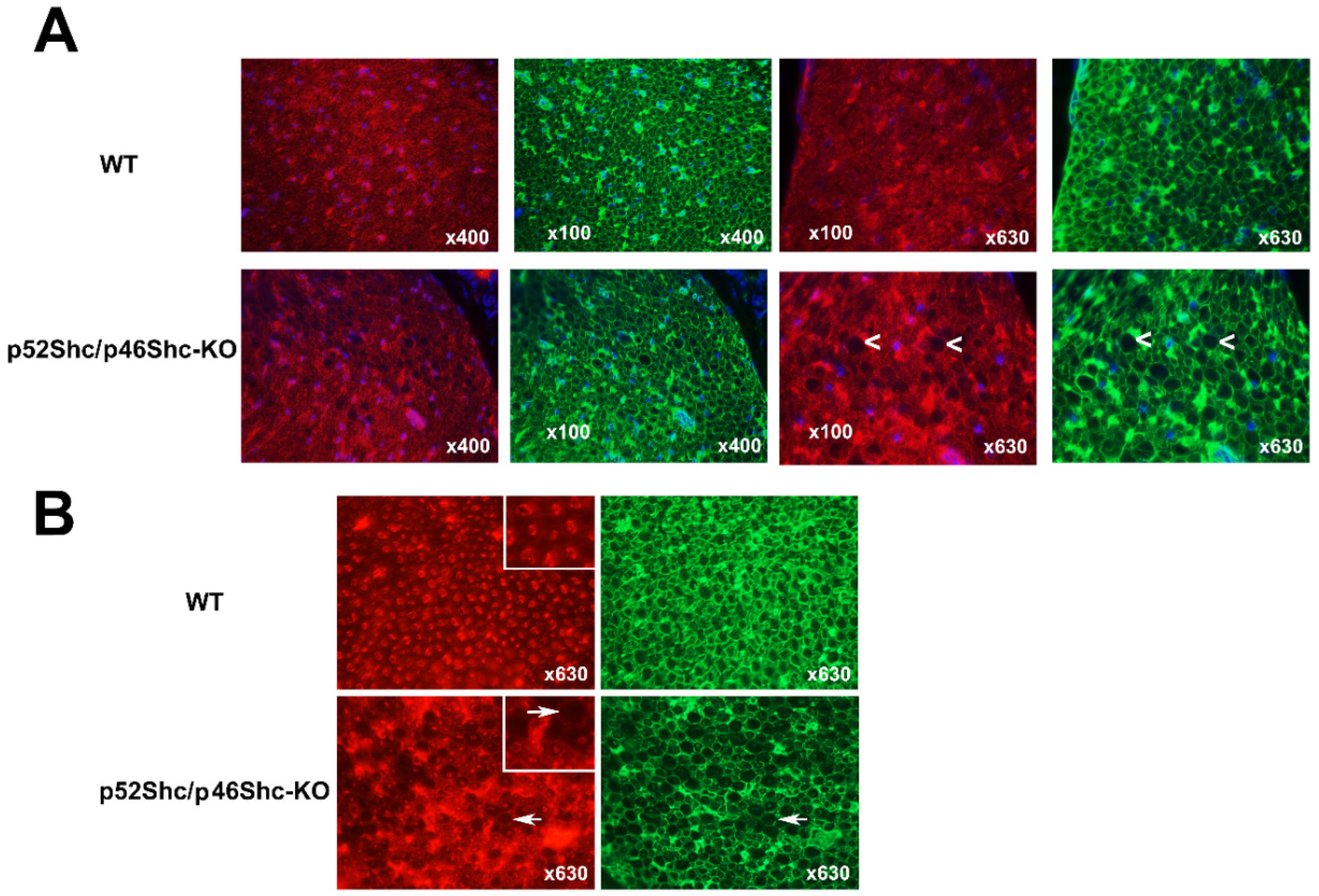
2.5. Analysis of p52Shc/46Shc-KO Sciatic Nerve Revealed Abnormalities
2.6. Analysis of p52Shc/p46Shc-KO Cardiac Muscle Revealed Features of Cardiomyopathy
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Harvesting Muscle Samples
4.3. Gait Analysis
4.4. Histochemical and Immunohistochemical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wills, M.K.B.; Jones, N. Teaching an old dogma new tricks: Twenty years of Shc adaptor signalling. Biochem. J. 2012, 447, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, R.; Henderson, J.T.; O’Bryan, J.P.; Elia, A.J.; Saxton, T.M.; Pawson, T. The Mammalian ShcB and ShcC Phosphotyrosine Docking Proteins Function in the Maturation of Sensory and Sympathetic Neurons. Neuron 2000, 28, 819–833. [Google Scholar] [CrossRef] [Green Version]
- Wright, K.D.; Staruschenko, A.; Sorokin, A. Role of adaptor protein p66Shc in renal pathologies. Am. J. Physiol. Physiol. 2018, 314, F143–F153. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, E.; Mele, S.; Salcini, A.E.; Pelicci, G.; Lai, K.V.; Superti-Furga, G.; Pawson, T.; Di Fiore, P.P.; Lanfrancone, L.; Pelicci, P.G. Opposite effects of the p52shc/p46shc and p66shc splicing isoforms on the EGF receptor-MAP kinase-fos signalling pathway. EMBO J. 1997, 16, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Tomilov, A.; Tomilova, N.; Shan, Y.; Hagopian, K.; Bettaieb, A.; Kim, K.; Pelicci, P.G.; Haj, F.; Ramsey, J.; Cortopassi, G. p46Shc Inhibits Thiolase and Lipid Oxidation in Mitochondria. J. Biol. Chem. 2016, 291, 12575–12585. [Google Scholar] [CrossRef] [Green Version]
- Purdom, S.; Chen, Q.M. p66Shc: At the crossroad of oxidative stress and the genetics of aging. Trends Mol. Med. 2003, 9, 206–210. [Google Scholar] [CrossRef]
- Downward, J. Control of ras activation. Cancer Surv. 1996, 27, 87–100. [Google Scholar]
- Foschi, M.; Chari, S.; Dunn, M.J.; Sorokin, A. Biphasic activation of p21ras by endothelin-1 sequentially activates the ERK cascade and phosphatidylinositol 3-kinase. EMBO J. 1997, 16, 6439–6451. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.; Palygin, O.; Rufanova, V.A.; Chong, A.; Lazar, J.; Jacob, H.J.; Mattson, D.; Roman, R.J.; Williams, J.M.; Cowley, A.W.; et al. p66Shc regulates renal vascular tone in hypertension-induced nephropathy. J. Clin. Investig. 2016, 126, 2533–2546. [Google Scholar] [CrossRef]
- Migliaccio, E.; Giorgio, M.; Mele, S.; Pelicci, G.; Reboldi, P.; Pandolfi, P.P.; Lanfrancone, L.; Pelicci, P.G. The p66shc adaptor protein controls oxidative stress response and life span in mammals. Nat. Cell Biol. 1999, 402, 309–313. [Google Scholar] [CrossRef]
- Wright, K.D.; Miller, B.S.; El-Meanawy, S.; Tsaih, S.-W.; Banerjee, A.; Geurts, A.M.; Sheinin, Y.; Sun, Y.; Kalyanaraman, B.; Rui, H.; et al. The p52 isoform of SHC1 is a key driver of breast cancer initiation. Breast Cancer Res. 2019, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Edstrom, E.; Altun, M.; Ulfhake, B. Differential regulation of Shc adaptor proteins in skeletal muscle, spinal cord and forebrain of aged rats with sensorimotor impairment. Aging Cell 2003, 2, 47–57. [Google Scholar] [CrossRef]
- Baker, J.M. Gait Disorders. Am. J. Med. 2018, 131, 602–607. [Google Scholar] [CrossRef] [PubMed]
- American Geriatrics Society; British Geriatrics Society; American Academy of Orthopaedic Surgeons Panel on Falls Prevention. Guideline for the prevention of falls in older persons. J. Am. Geriatr. Soc. 2001, 49, 664–672. [Google Scholar] [CrossRef]
- Prado, F.P.; Dos Santos, D.O.; Blefari, V.; Silva, C.A.; Machado, J.; Kettelhut, Í.d.C.; Ramos, S.G.; Baruffi, M.D.; Salgado, H.C.; Prado, C.M. Early dystrophin loss is coincident with the transition of compensated cardiac hypertrophy to heart failure. PLoS ONE 2017, 12, e0189469. [Google Scholar] [CrossRef] [Green Version]
- Sammani, A.; Baas, A.; Asselbergs, F.; Riele, A.T. Diagnosis and Risk Prediction of Dilated Cardiomyopathy in the Era of Big Data and Genomics. J. Clin. Med. 2021, 10, 921. [Google Scholar] [CrossRef] [PubMed]
- Vanderlaan, R.D.; Hardy, W.R.; Kabir, M.G.; Pasculescu, A.; Jones, N.; Detombe, P.P.; Backx, P.H.; Pawson, T. The ShcA Phosphotyrosine Docking Protein Uses Distinct Mechanisms to Regulate Myocyte and Global Heart Function. Circ. Res. 2011, 108, 184–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, W.R.; Li, L.; Wang, Z.; Sedy, J.; Fawcett, J.; Frank, E.; Kucera, J.; Pawson, T. Combinatorial ShcA Docking Interactions Support Diversity in Tissue Morphogenesis. Science 2007, 317, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Badorff, C.; Knowlton, K.U. Dystrophin disruption in enterovirus-induced myocarditis and dilated cardiomyopathy: From bench to bedside. Med. Microbiol. Immunol. 2004, 193, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Mlih, M.; Host, L.; Martin, S.; Niederhoffer, N.; Monassier, L.; Terrand, J.; Messaddeq, N.; Radke, M.; Gotthardt, M.; Bruban, V.; et al. The Src Homology and Collagen A (ShcA) Adaptor Protein Is Required for the Spatial Organization of the Costamere/Z-disk Network during Heart Development. J. Biol. Chem. 2015, 290, 2419–2430. [Google Scholar] [CrossRef] [Green Version]
- Macefield, V.G.; Norcliffe-Kaufmann, L.; Gutiérrez, J.; Axelrod, F.B.; Kaufmann, H. Can loss of muscle spindle afferents explain the ataxic gait in Riley-Day syndrome? Brain 2011, 134, 3198–3208. [Google Scholar] [CrossRef] [Green Version]
- Rask-Andersen, M.; Karlsson, T.; Ek, W.E.; Johansson, Å. Genome-wide association study of body fat distribution identifies adiposity loci and sex-specific genetic effects. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.-M.V.; Pawson, T. The ShcA phosphotyrosine docking protein sensitizes cardiovascular signaling in the mouse embryo. Genes Dev. 2000, 14, 1132–1145. [Google Scholar] [PubMed]
- Kozak, M. Regulation of translation via mRNA structure in prokaryotes and eukaryotes. Gene 2005, 361, 13–37. [Google Scholar] [CrossRef]
- Mehrotra, P.; Ullah, M.; Collett, J.A.; Myers, S.L.; Dwinell, M.R.; Geurts, A.M.; Basile, D.P. Mutation of RORγT reveals a role for Th17 cells in both injury and recovery from renal ischemia-reperfusion injury. Am. J. Physiol. Physiol. 2020, 319, F796–F808. [Google Scholar] [CrossRef] [PubMed]
- Spires, D.; Ilatovskaya, D.V.; Levchenko, V.; North, P.E.; Geurts, A.M.; Palygin, O.; Staruschenko, A. Protective role of Trpc6 knockout in the progression of diabetic kidney disease. Am. J. Physiol. Physiol. 2018, 315, F1091–F1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehrenbach, D.J.; Abais-Battad, J.M.; Dasinger, J.H.; Lund, H.; Keppel, T.; Zemaj, J.; Cherian-Shaw, M.; Gundry, R.L.; Geurts, A.M.; Dwinell, M.R.; et al. Sexual Dimorphic Role of CD14 (Cluster of Differentiation 14) in Salt-Sensitive Hypertension and Renal Injury. Hypertension 2021, 77, 228–240. [Google Scholar] [CrossRef]
- Geurts, A.M.; Cost, G.J.; Rémy, S.; Cui, X.; Tesson, L.; Usal, C.; Ménoret, S.; Jacob, H.J.; Anegon, I.; Buelow, R. Generation of Gene-Specific Mutated Rats Using Zinc-Finger Nucleases. Methods Mol. Biol. 2010, 597, 211–225. [Google Scholar] [PubMed]
- Kostrominova, T.Y. Application of WGA lectin staining for visualization of the connective tissue in skeletal muscle, bone, and ligament/tendon studies. Microsc. Res. Tech. 2011, 74, 18–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, B.; Kostrominova, T.Y.; Geurts, A.M.; Sorokin, A. Double p52Shc/p46Shc Rat Knockout Demonstrates Severe Gait Abnormalities Accompanied by Dilated Cardiomyopathy. Int. J. Mol. Sci. 2021, 22, 5237. https://doi.org/10.3390/ijms22105237
Miller B, Kostrominova TY, Geurts AM, Sorokin A. Double p52Shc/p46Shc Rat Knockout Demonstrates Severe Gait Abnormalities Accompanied by Dilated Cardiomyopathy. International Journal of Molecular Sciences. 2021; 22(10):5237. https://doi.org/10.3390/ijms22105237
Chicago/Turabian StyleMiller, Bradley, Tatiana Y. Kostrominova, Aron M. Geurts, and Andrey Sorokin. 2021. "Double p52Shc/p46Shc Rat Knockout Demonstrates Severe Gait Abnormalities Accompanied by Dilated Cardiomyopathy" International Journal of Molecular Sciences 22, no. 10: 5237. https://doi.org/10.3390/ijms22105237
APA StyleMiller, B., Kostrominova, T. Y., Geurts, A. M., & Sorokin, A. (2021). Double p52Shc/p46Shc Rat Knockout Demonstrates Severe Gait Abnormalities Accompanied by Dilated Cardiomyopathy. International Journal of Molecular Sciences, 22(10), 5237. https://doi.org/10.3390/ijms22105237