
Effects of Aqueous Dispersions of C60, C70 and Gd@C82 Fullerenes on Genes Involved in Oxidative Stress and Anti-Inflammatory Pathways

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Preparation and Characterization of Aqueous Fullerene Dispersions
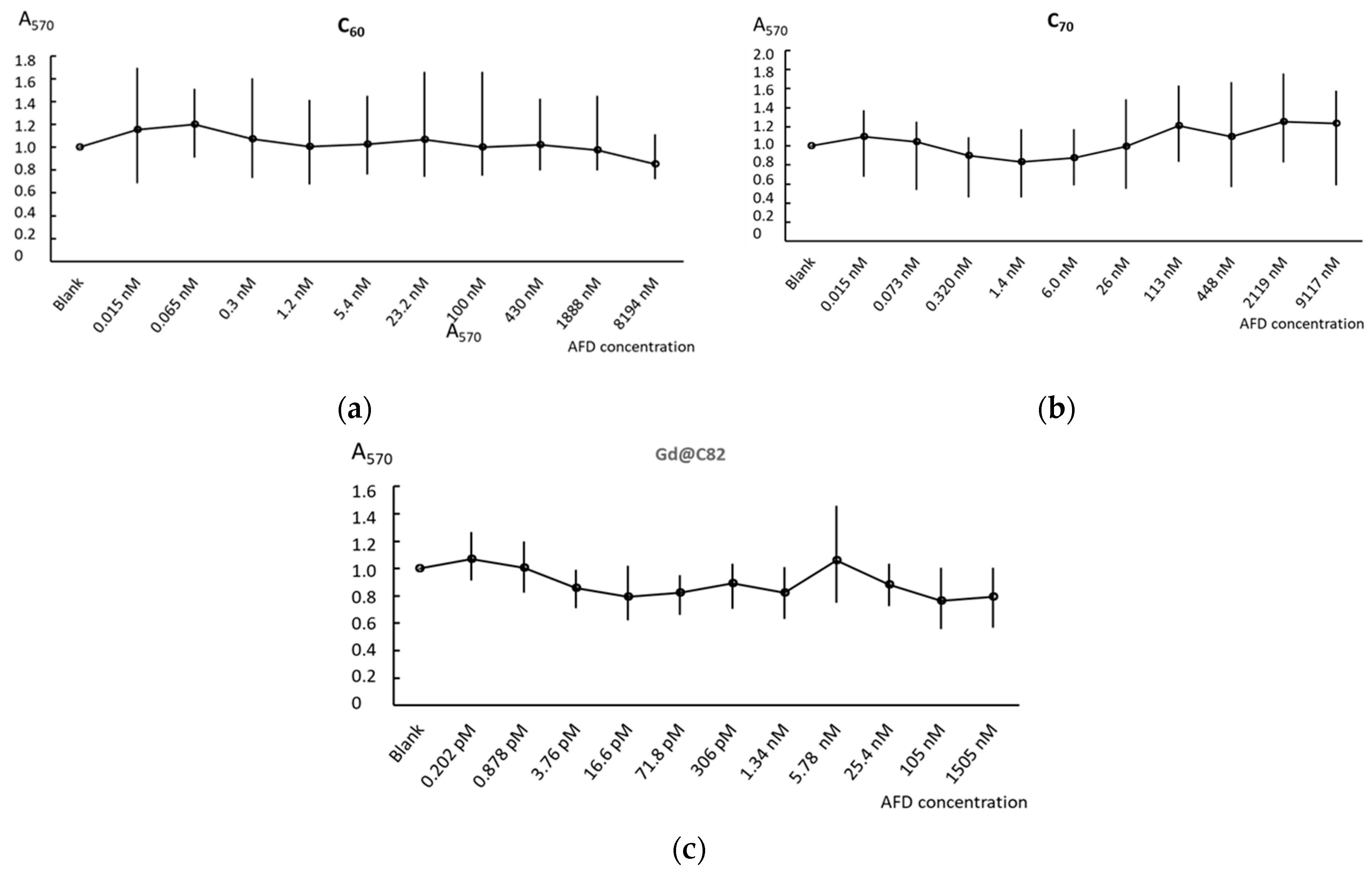
2.2. Cell Viability
2.3. Mitochondrial Potential
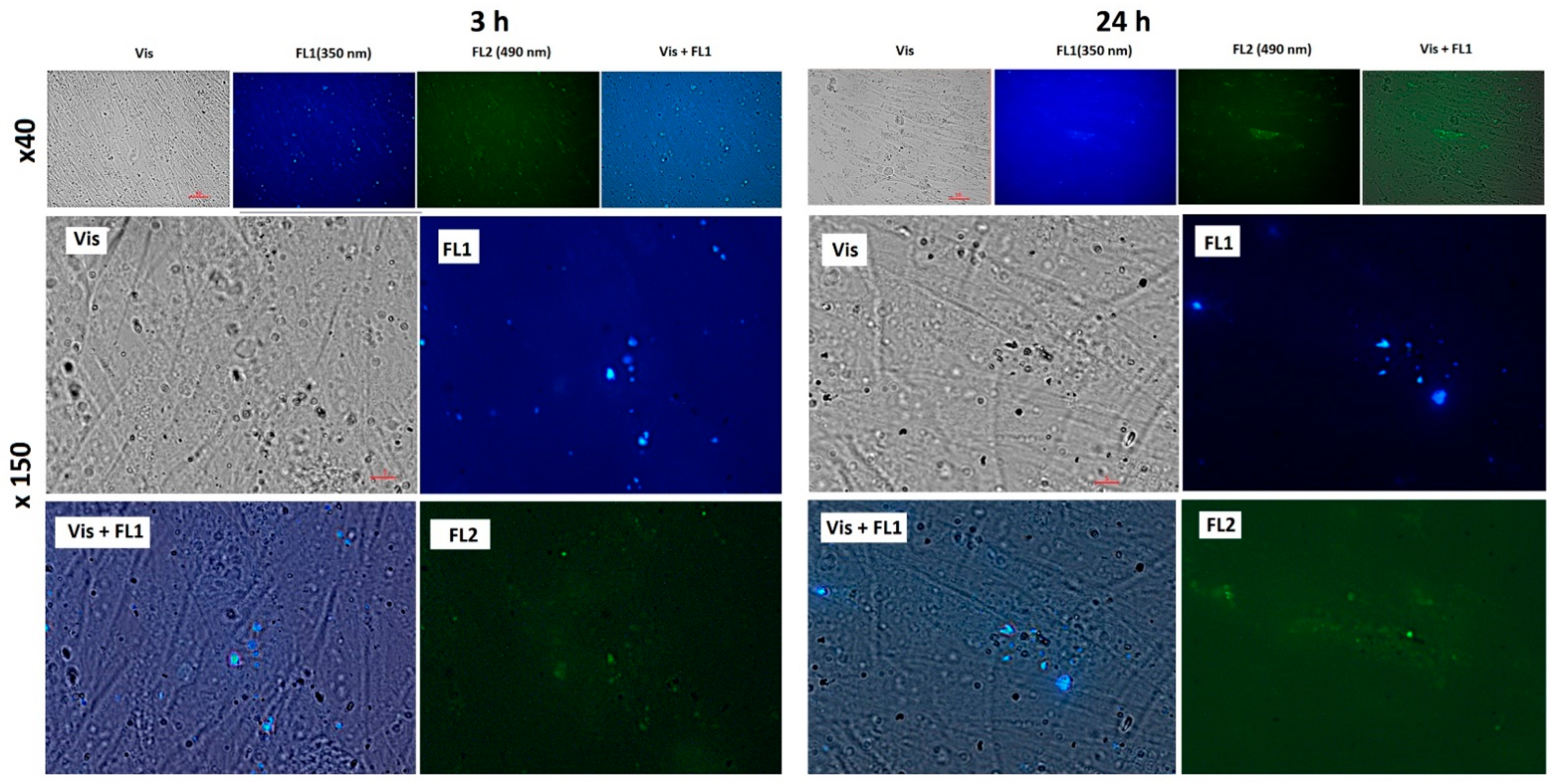
2.4. Localization of Fullerenes inside Human Fetal Lung Fibroblasts
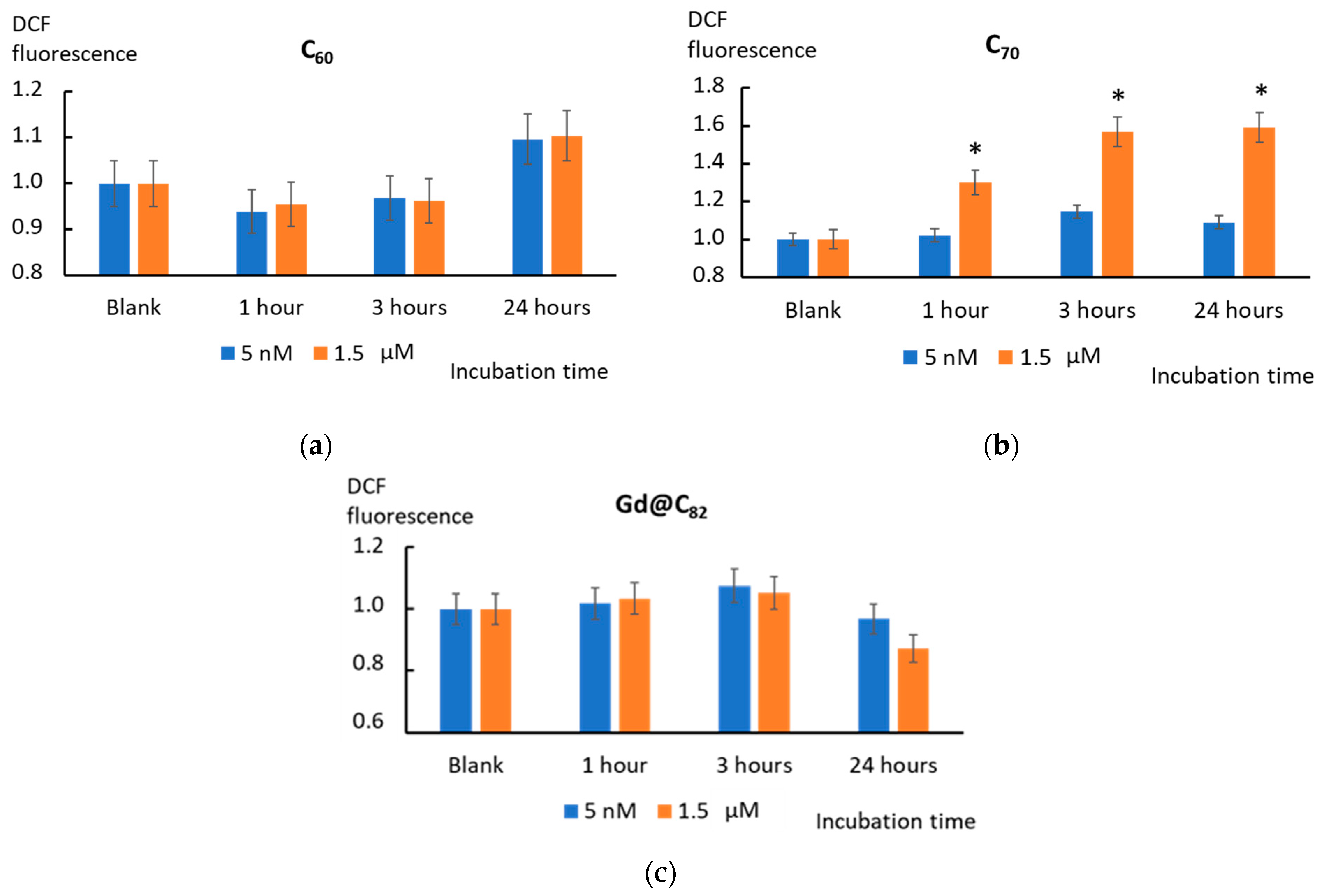
2.5. Intracellular ROS
2.6. NOX4 Expression
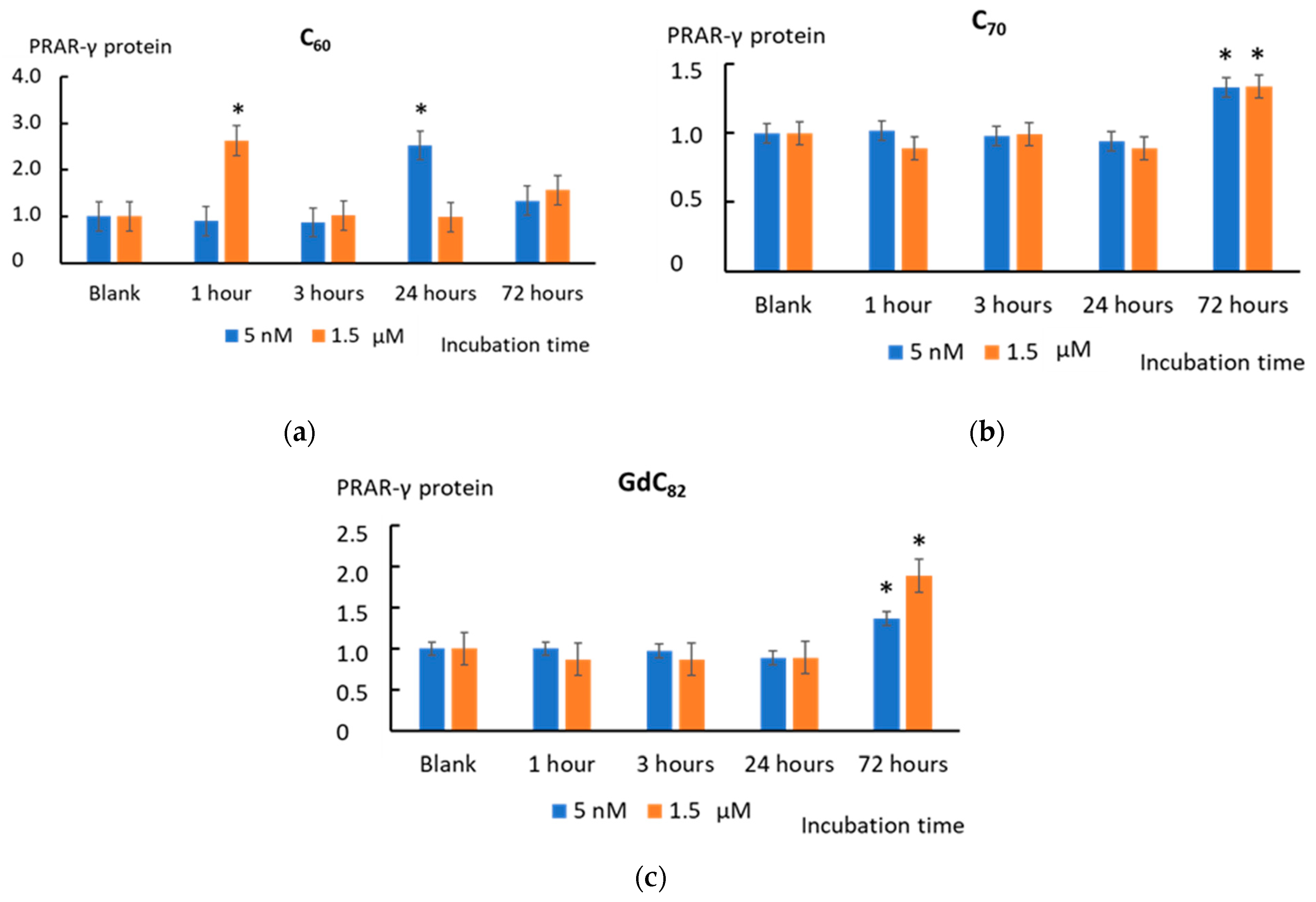
2.7. NFκB Pathway and PRAR-γ
2.8. NRF2 Pathway
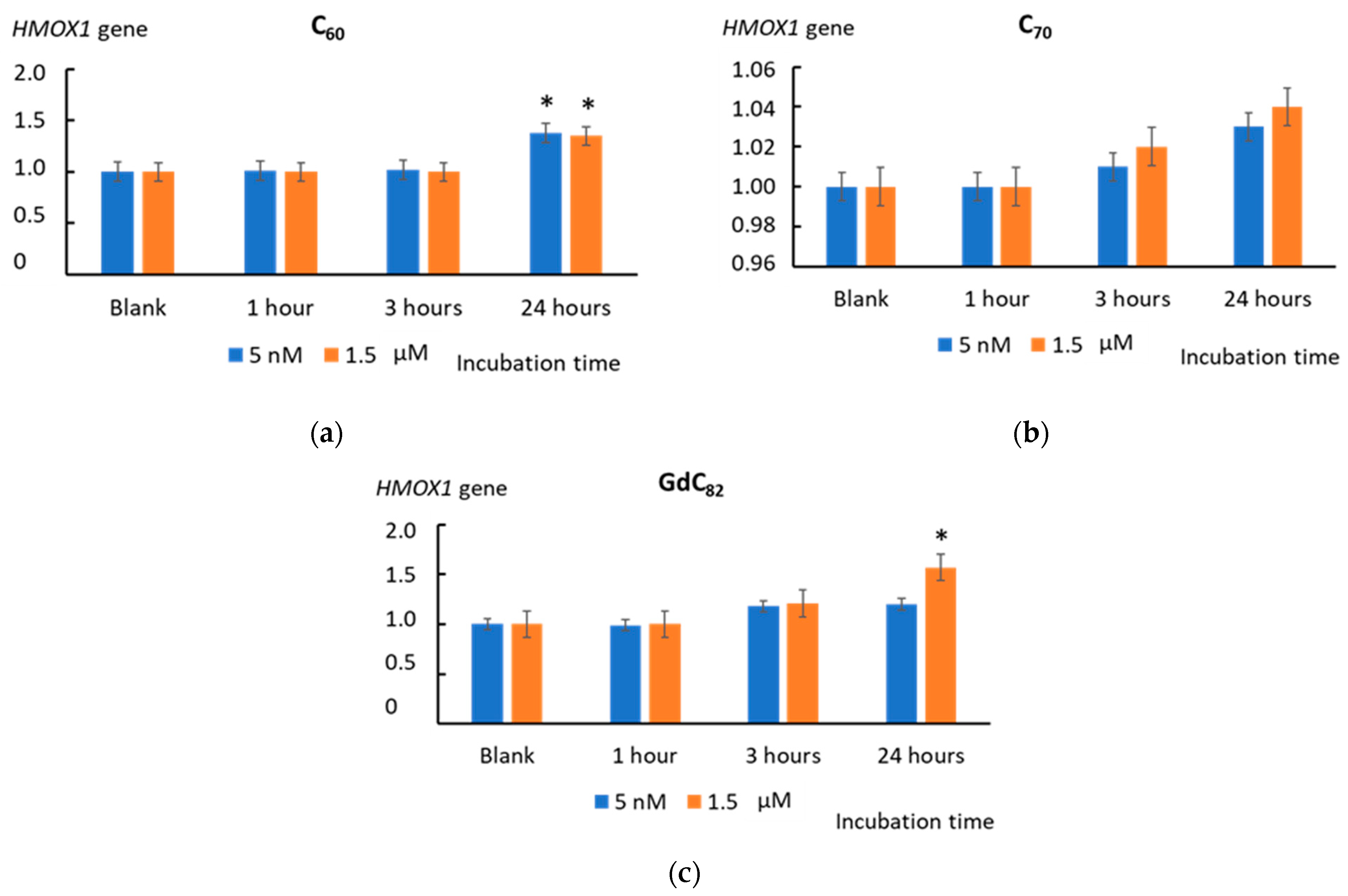
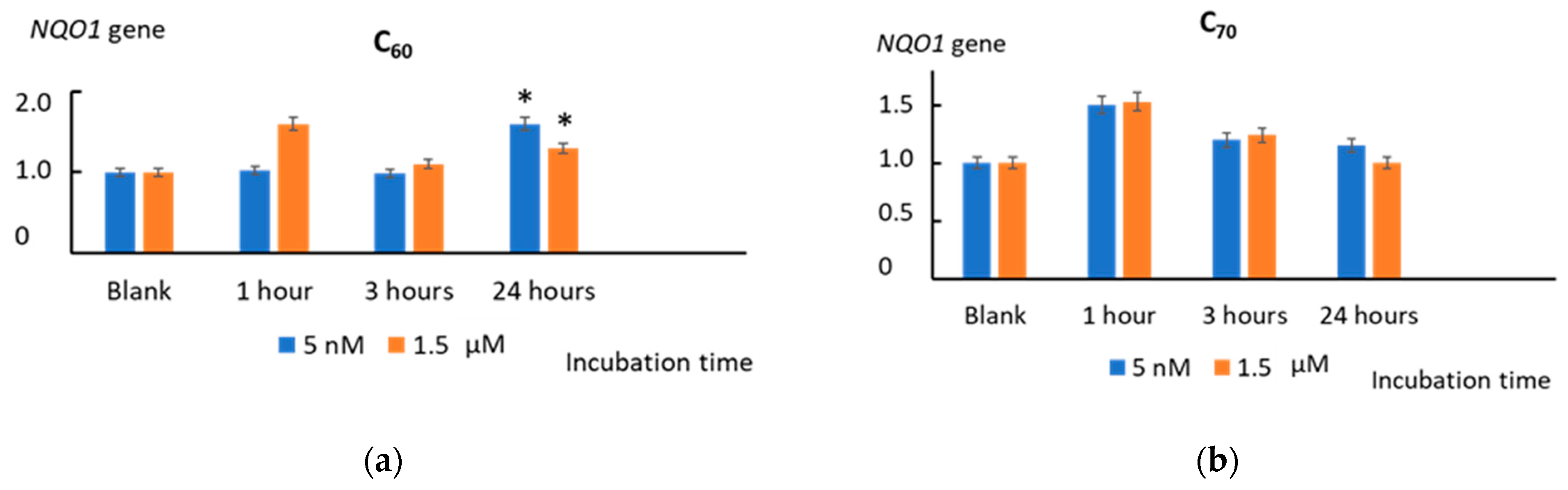
2.9. Heme Oxygenase 1 and NAD(P)H Quinone Dehydrogenase 1
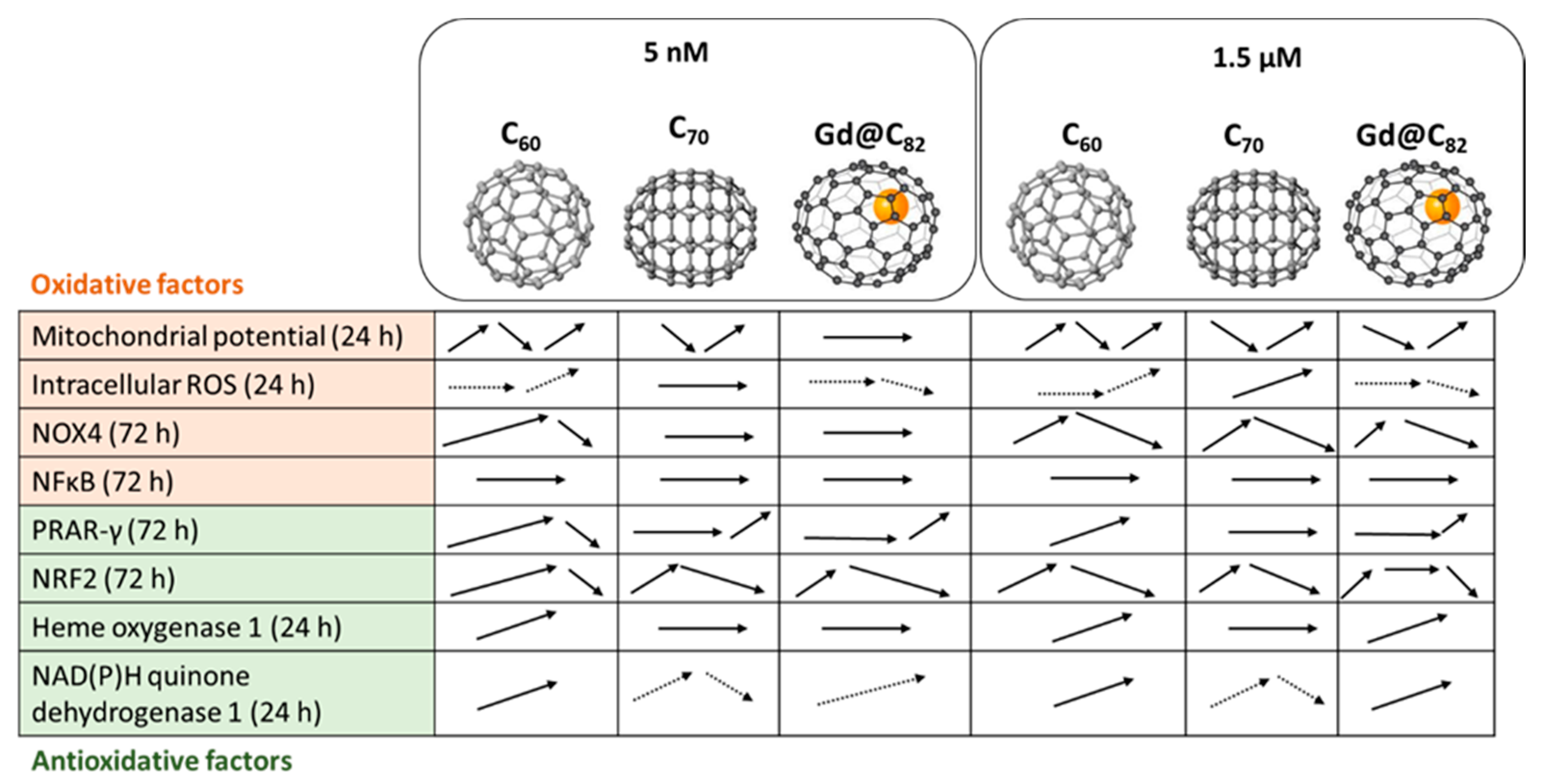
3. Discussion
4. Materials and Methods
4.1. AFD Preparation and Characterization
4.1.1. Reagents
4.1.2. Preparation of Aqueous Fullerene Dispersions by Direct Ultrasound Probe Sonication
4.1.3. Characterization
4.2. Cell Culture
4.3. MTT Assay and Mitotracker Test
4.4. Reactive Oxygen Species Assays
4.5. Quantification of mRNA Levels
4.6. Fluorescence Microscopy
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamashita, T.; Yamashita, K.; Nabeshi, H.; Yoshikawa, T.; Yoshioka, Y.; Tsunoda, S.I.; Tsutsumi, Y. Carbon Nanomaterials: Efficacy and Safety for Nanomedicine. Materials 2012, 5, 350–363. [Google Scholar] [CrossRef]
- Dellinger, A.; Zhou, Z.; Connor, J.; Madhankumar, A.B.; Pamujula, S.; Sayes, C.M.; Kepley, C.L. Application of fullerenes in nanomedicine: An update. Nanomedicine 2013, 8, 1191–1208. [Google Scholar] [CrossRef] [Green Version]
- Partha, R.; Conyers, J.L. Biomedical applications of functionalized fullerene-based nanomaterials. Int. J. Nanomed. 2009, 4, 261–275. [Google Scholar]
- Tagmatarchis, N.; Shinohara, H. Fullerenes in medicinal chemistry and their biological applications. Mini Rev. Med. Chem. 2001, 1, 339–348. [Google Scholar] [CrossRef]
- Maeda, Y.; Tsuchiya, T.; Lu, X.; Takano, Y.; Akasaka, T.; Nagase, S. Current progress on the chemical functionalization and supramolecular chemistry of M@C82. Nanoscale 2011, 3, 2421–2429. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Raza, K. C60-fullerenes as Drug Delivery Carriers for Anticancer Agents: Promises and Hurdles. Pharm. Nanotechnol. 2017, 5, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Xu, X.; Zhang, X.; Hu, L. Development of calixarenes, cyclodextrins and fullerenes as new platforms for anti-HIV drug design: An overview. Mini Rev. Med. Chem. 2013, 13, 1160–1165. [Google Scholar] [CrossRef] [PubMed]
- Ghiassi, K.B.; Olmstead, M.M.; Balch, A.L. Gadolinium-containing endohedral fullerenes: Structures and function as magnetic resonance imaging (MRI) agents. Dalton Trans. 2014, 43, 7346–7358. [Google Scholar] [CrossRef]
- Sachkova, A.S.; Kovel, E.S.; Churilov, G.N.; Guseynov, O.A.; Bondar, A.A.; Dubinina, I.A.; Kudryasheva, N.S. On mechanism of antioxidant effect of fullerenols. Biochem. Biophys. Rep. 2017, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Injac, R.; Prijatelj, M.; Strukelj, B. Fullerenol nanoparticles: Toxicity and antioxidant activity. Methods Mol. Biol. 2013, 1028, 75–100. [Google Scholar] [CrossRef] [PubMed]
- Dugan, L.L.; Lovett, E.G.; Quick, K.L.; Lotharius, J.; Lin, T.T.; O’Malley, K.L. Fullerene-based antioxidants and neurodegenerative disorders. Parkinsonism Relat. Disord. 2001, 7, 243–246. [Google Scholar] [CrossRef]
- Zay, S.Y.; Zavodovskyi, D.A.; Bogutska, K.I.; Nozdrenko, D.N.; Prylutskyy, Y.I. Prospects of C60 Fullerene Application as a Mean of Prevention and Correction of Ischemic-Reperfusion Injury in the Skeletal Muscle Tissue. Fiziol. Zh. 2016, 62, 66–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Cui, Q.; Li, X.J.; Jin, L. The applications of buckminsterfullerene C60 and derivatives in orthopaedic research. Connect. Tissue Res. 2014, 55, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.M.; Fang, S.F.; Lin, S.Z.; Chou, C.K.; Luh, T.Y.; Ho, L.T. Local carboxyfullerene protects cortical infarction in rat brain. Neurosci. Res. 2002, 43, 317–321. [Google Scholar] [CrossRef]
- Chistyakov, V.A.; Smirnova, Y.O.; Prazdnova, E.V.; Soldatov, A.V. Possible mechanisms of fullerene C60 antioxidant action. Biomed. Res. Int. 2013, 2013, 821498. [Google Scholar] [CrossRef] [Green Version]
- Galvan, Y.P.; Alperovich, I.; Zolotukhin, P.; Prazdnova, E.; Mazanko, M.; Belanova, A.; Chistyakov, V. Fullerenes as Anti-Aging Antioxidants. Curr. Aging Sci. 2017, 10, 56–67. [Google Scholar] [CrossRef]
- Li, J.; Chen, L.; Yan, L.; Gu, Z.; Chen, Z.; Zhang, A.; Zhao, F. A Novel Drug Design Strategy: An Inspiration from Encaging Tumor by Metallofullerenol Gd@C82(OH)22. Molecules 2019, 24, 2387. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Liang, X.; Chen, X.; Zhao, Y. Biological characterizations of [Gd@C82(OH)22]n nanoparticles as fullerene derivatives for cancer therapy. Integr. Biol. 2013, 5, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Ershova, E.S.; Sergeeva, V.; Chausheva, A.I.; Zheglo, D.G.; Nikitina, V.; Smirnova, T.D.; Kameneva, L.V.; Porokhovnik, L.N.; Kutsev, S.I.; Troshin, P.A.; et al. Toxic and DNA damaging effects of a functionalized fullerene in human embryonic lung fibroblasts. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2016, 805, 46–57. [Google Scholar] [CrossRef]
- Ershova, E.S.; Sergeeva, V.A.; Tabakov, V.J.; Kameneva, L.A.; Porokhovnik, L.N.; Voronov, I.I.; Khakina, E.A.; Troshin, P.A.; Kutsev, S.I.; Veiko, N.N.; et al. Functionalized Fullerene Increases NF-kappaB Activity and Blocks Genotoxic Effect of Oxidative Stress in Serum-Starving Human Embryo Lung Diploid Fibroblasts. Oxid. Med. Cell. Longev. 2016, 2016, 9895245. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.; Morimoto, Y.; Ogami, A.; Myojyo, T.; Tanaka, I.; Shimada, M.; Wang, W.N.; Endoh, S.; Uchida, K.; Nakazato, T.; et al. Gene expression profiles in rat lung after inhalation exposure to C60 fullerene particles. Toxicology 2009, 258, 47–55. [Google Scholar] [CrossRef]
- Jovanovic, B.; Ji, T.; Palic, D. Gene expression of zebrafish embryos exposed to titanium dioxide nanoparticles and hydroxylated fullerenes. Ecotoxicol. Environ. Saf. 2011, 74, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi-Tago, M.; Nagata, T.; Tago, K.; Tsukada, M.; Tanaka, K.; Nakamura, S.; Mashino, T.; Kasahara, T. Fullerene derivative prevents cellular transformation induced by JAK2 V617F mutant through inhibiting c-Jun N-terminal kinase pathway. Cell. Signal. 2012, 24, 2024–2034. [Google Scholar] [CrossRef]
- Etem, E.O.; Bal, R.; Akagac, A.E.; Kuloglu, T.; Tuzcu, M.; Andrievsky, G.V.; Buran, I.; Nedzvetsky, V.S.; Baydas, G. The effects of hydrated C60 fullerene on gene expression profile of TRPM2 and TRPM7 in hyperhomocysteinemic mice. J. Recept. Signal. Transduct. Res. 2014, 34, 317–324. [Google Scholar] [CrossRef]
- Ye, S.; Chen, M.; Jiang, Y.; Chen, M.; Zhou, T.; Wang, Y.; Hou, Z.; Ren, L. Polyhydroxylated fullerene attenuates oxidative stress-induced apoptosis via a fortifying Nrf2-regulated cellular antioxidant defence system. Int. J. Nanomed. 2014, 9, 2073–2087. [Google Scholar] [CrossRef] [Green Version]
- Hao, T.; Zhou, J.; Lu, S.; Yang, B.; Wang, Y.; Fang, W.; Jiang, X.; Lin, Q.; Li, J.; Wang, C. Fullerene mediates proliferation and cardiomyogenic differentiation of adipose-derived stem cells via modulation of MAPK pathway and cardiac protein expression. Int. J. Nanomed. 2016, 11, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Nie, X.; Tang, J.; Liu, Y.; Cai, R.; Miao, Q.; Zhao, Y.; Chen, C. Fullerenol inhibits the crosstalk between bone marrow-derived mesenchymal stem cells and tumor cells by regulating MAPK signaling. Nanomedicine 2017, 13, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Sollogoub, M.; Zhang, Y. Biological applications of hydrophilic C60 derivatives (hC60s)-A structural perspective. Eur. J. Med. Chem. 2016, 115, 438–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovel, E.S.; Sachkova, A.S.; Vnukova, N.G.; Churilov, G.N.; Knyazeva, E.M.; Kudryasheva, N.S. Antioxidant Activity and Toxicity of Fullerenols via Bioluminescence Signaling: Role of Oxygen Substituents. Int. J. Mol. Sci. 2019, 20, 2324. [Google Scholar] [CrossRef] [Green Version]
- Labille, J.; Masion, A.; Ziarelli, F.; Rose, J.; Brant, J.; Villieras, F.; Pelletier, M.; Borschneck, D.; Wiesner, M.R.; Bottero, J.Y. Hydration and dispersion of C60 in aqueous systems: The nature of water-fullerene interactions. Langmuir 2009, 25, 11232–11235. [Google Scholar] [CrossRef] [PubMed]
- Mikheev, I.V.; Pirogova, M.O.; Usoltseva, L.O.; Uzhel, A.S.; Bolotnik, T.A.; Kareev, I.E.; Bubnov, V.P.; Lukonina, N.S.; Volkov, D.S.; Goryunkov, A.A.; et al. Green and rapid preparation of long-term stable aqueous dispersions of fullerenes and endohedral fullerenes: The pros and cons of an ultrasonic probe. Ultrason. Sonochem. 2021, 73, 105533. [Google Scholar] [CrossRef]
- Mosharova, I.V.; Dallakyan, G.A.; Mikheev, I.V.; Il’inskii, V.V.; Akulova, A.Y. Changes in the Quantitative and Functional Characteristics of Bacterioplankton under the Influence of Aqueous Unmodified Fullerene capital ES, Cyrillic60 Dispersions. Dokl. Biochem. Biophys. 2019, 487, 256–259. [Google Scholar] [CrossRef]
- Mikheev, I.V.; Sozarukova, M.M.; Proskurnina, E.V.; Kareev, I.E.; Proskurnin, M.A. Non-Functionalized Fullerenes and Endofullerenes in Aqueous Dispersions as Superoxide Scavengers. Molecules 2020, 25, 2506. [Google Scholar] [CrossRef]
- Bubnov, V.P.; Laukhina, E.E.; Kareev, I.E.; Koltover, V.K.; Prokhorova, T.G.; Yagubskii, E.B.; Kozmin, Y.P. Endohedral Metallofullerenes: A Convenient Gram-Scale Preparation. Chem. Mater. 2002, 14, 1004–1008. [Google Scholar] [CrossRef]
- Ko, W.B.; Heo, J.Y.; Nam, J.H.; Lee, K.B. Synthesis of a water-soluble fullerene [C60] under ultrasonication. Ultrasonics 2004, 41, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Taurozzi, J.S.; Hackley, V.A.; Wiesner, M. Preparation of nanoparticle dispersions from powdered material using ultrasonic disruption. NIST Spec. Publ. 2012, 1200, 1200–1202. [Google Scholar]
- Braun, T. (Ed.) Nuclear and Radiation Chemical Approaches to Fullerene Science; Springer: Dordrecht, The Netherlands, 2000; Volume 1, p. 203. [Google Scholar]
- Treubig, J.M.; Brown, P.R. Analysis of C60 and C70 fullerenes using high-performance liquid chromatography–Fourier transform infrared spectroscopy. J. Chromatogr. A 2002, 960, 135–142. [Google Scholar] [CrossRef]
- Wang, J.; Li, Z.; Li, S.; Qi, W.; Liu, P.; Liu, F.; Ye, Y.; Wu, L.; Wang, L.; Wu, W. Adsorption of Cu(II) on Oxidized Multi-Walled Carbon Nanotubes in the Presence of Hydroxylated and Carboxylated Fullerenes. PLoS ONE 2013, 8, e72475. [Google Scholar] [CrossRef]
- Cataldo, F.; Iglesias-Groth, S.; Manchado, A. Low and High Temperature Infrared Spectroscopy of C60 and C70 Fullerenes. Fuller. Nanotub. Carbon Nanostructures 2010, 18, 224–235. [Google Scholar] [CrossRef]
- Krause, M.; Kuran, P.; Kirbach, U.; Dunsch, L. Raman and infrared spectra of Tm@C82 and Gd@C82. Carbon 1999, 37, 113–115. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Suzuki, T.; Ishii, H.; Nakae, D.; Ogata, A. Cytotoxic effects of hydroxylated fullerenes on isolated rat hepatocytes via mitochondrial dysfunction. Arch. Toxicol. 2011, 85, 1429–1440. [Google Scholar] [CrossRef]
- Dellinger, A.; Zhou, Z.; Norton, S.K.; Lenk, R.; Conrad, D.; Kepley, C.L. Uptake and distribution of fullerenes in human mast cells. Nanomedicine 2010, 6, 575–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghadam, Z.M.; Henneke, P.; Kolter, J. From Flies to Men: ROS and the NADPH Oxidase in Phagocytes. Front. Cell Dev. Biol. 2021, 9, 628991. [Google Scholar] [CrossRef]
- Scirpo, R.; Fiorotto, R.; Villani, A.; Amenduni, M.; Spirli, C.; Strazzabosco, M. Stimulation of nuclear receptor peroxisome proliferator-activated receptor-gamma limits NF-kappaB-dependent inflammation in mouse cystic fibrosis biliary epithelium. Hepatology 2015, 62, 1551–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricote, M.; Glass, C.K. PPARs and molecular mechanisms of transrepression. Biochim. Biophys. Acta 2007, 1771, 926–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Scaduto, R.C.; Grotyohann, L.W. Measurement of Mitochondrial Membrane Potential Using Fluorescent Rhodamine Derivatives. Biophys. J. 1999, 76, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Creed, S.; McKenzie, M. Measurement of Mitochondrial Membrane Potential with the Fluorescent Dye Tetramethylrhodamine Methyl Ester (TMRM). Methods Mol. Biol. 2019, 1928, 69–76. [Google Scholar] [CrossRef]
- Boltalina, O.; Ioffe, I.; Sorokin, I.; Sidorov, L.N. Electron Affinity of Some Endohedral Lanthanide Fullerenes. J. Phys. Chem. A 1997, 101, 9561–9563. [Google Scholar] [CrossRef]
- Ptasinska, S.; Echt, O.; Denifl, S.; Stano, M.; Sulzer, P.; Zappa, F.; Stamatovic, A.; Scheier, P.; Mark, T.D. Electron attachment to higher fullerenes and to Sc3N@C80. J. Phys. Chem. A 2006, 110, 8451–8456. [Google Scholar] [CrossRef]
- Almeida, M.; Ambrogini, E.; Han, L.; Manolagas, S.C.; Jilka, R.L. Increased lipid oxidation causes oxidative stress, increased peroxisome proliferator-activated receptor-gamma expression, and diminished pro-osteogenic Wnt signaling in the skeleton. J. Biol. Chem. 2009, 284, 27438–27448. [Google Scholar] [CrossRef] [Green Version]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and-independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xiang, Y. Molecular and cellular basis for the unique functioning of Nrf1, an indispensable transcription factor for maintaining cell homoeostasis and organ integrity. Biochem. J. 2016, 473, 961–1000. [Google Scholar] [CrossRef]
- O’Connell, M.A.; Hayes, J.D. The Keap1/Nrf2 pathway in health and disease: From the bench to the clinic. Biochem. Soc. Trans. 2015, 43, 687–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikheev, I.V.; Khimich, E.S.; Rebrikova, A.T.; Volkov, D.S.; Proskurnin, M.A.; Korobov, M.V. Quasi-equilibrium distribution of pristine fullerenes C60 and C70 in a water–toluene system. Carbon 2017, 111, 191–197. [Google Scholar] [CrossRef]
- Mikheev, I.V.; Usoltseva, L.O.; Ivshukov, D.A.; Volkov, D.S.; Korobov, M.V.; Proskurnin, M.A. Approach to the assessment of size-dependent thermal properties of disperse solutions: Time-resolved photothermal lensing of aqueous pristine fullerenes C60 and C70. J. Phys. Chem. C 2016, 120, 28270–28287. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fullerene | c, mM | Particle Size after a 0.22 μm Filter, nm | ζ-potential, mV | PDI |
|---|---|---|---|---|
| C60 | 0.083 1 | 110 ± 5 | −28.4 ± 0.2 | 0.109 ± 0.003 |
| C70 | 0.081 1 | 113 ± 2 | −29.5 ± 0.3 | 0.160 ± 0.008 |
| Gd@C82 | 0.022 2 | 95 ± 5 | −32.3 ± 0.3 | 0.123 ± 0.008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proskurnina, E.V.; Mikheev, I.V.; Savinova, E.A.; Ershova, E.S.; Veiko, N.N.; Kameneva, L.V.; Dolgikh, O.A.; Rodionov, I.V.; Proskurnin, M.A.; Kostyuk, S.V. Effects of Aqueous Dispersions of C60, C70 and Gd@C82 Fullerenes on Genes Involved in Oxidative Stress and Anti-Inflammatory Pathways. Int. J. Mol. Sci. 2021, 22, 6130. https://doi.org/10.3390/ijms22116130
Proskurnina EV, Mikheev IV, Savinova EA, Ershova ES, Veiko NN, Kameneva LV, Dolgikh OA, Rodionov IV, Proskurnin MA, Kostyuk SV. Effects of Aqueous Dispersions of C60, C70 and Gd@C82 Fullerenes on Genes Involved in Oxidative Stress and Anti-Inflammatory Pathways. International Journal of Molecular Sciences. 2021; 22(11):6130. https://doi.org/10.3390/ijms22116130
Chicago/Turabian StyleProskurnina, Elena V., Ivan V. Mikheev, Ekaterina A. Savinova, Elizaveta S. Ershova, Natalia N. Veiko, Larisa V. Kameneva, Olga A. Dolgikh, Ivan V. Rodionov, Mikhail A. Proskurnin, and Svetlana V. Kostyuk. 2021. "Effects of Aqueous Dispersions of C60, C70 and Gd@C82 Fullerenes on Genes Involved in Oxidative Stress and Anti-Inflammatory Pathways" International Journal of Molecular Sciences 22, no. 11: 6130. https://doi.org/10.3390/ijms22116130
APA StyleProskurnina, E. V., Mikheev, I. V., Savinova, E. A., Ershova, E. S., Veiko, N. N., Kameneva, L. V., Dolgikh, O. A., Rodionov, I. V., Proskurnin, M. A., & Kostyuk, S. V. (2021). Effects of Aqueous Dispersions of C60, C70 and Gd@C82 Fullerenes on Genes Involved in Oxidative Stress and Anti-Inflammatory Pathways. International Journal of Molecular Sciences, 22(11), 6130. https://doi.org/10.3390/ijms22116130