
Molecular Insights into the Oxygen-Sensing Pathway and Erythropoietin Expression Regulation in Erythropoiesis



Abstract
:1. Introduction
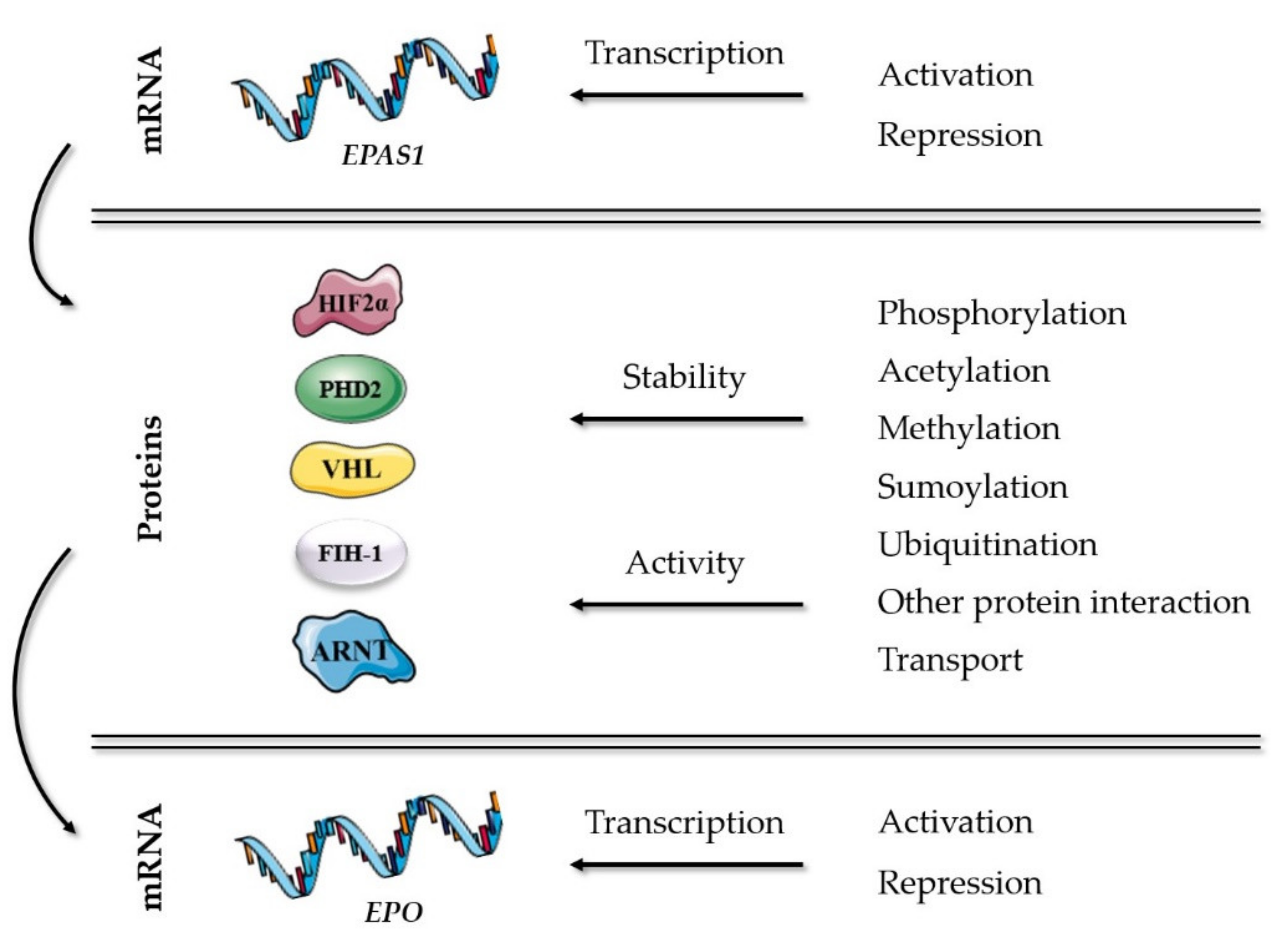
2. Main Players in the Oxygen-Sensing Pathway and EPO Expression Regulation
2.1. Regulation of HIF2α Transcription
2.2. Protein Interactions and Modifications of the Main Players in the Oxygen-Sensing Pathway
2.2.1. Post-Translational Modifications
2.2.2. Other Protein Interactions
2.2.3. Nuclear Transport of the Main Players in the Oxygen-Sensing Pathway
2.3. Regulation of EPO Transcription

{kind=link}
| Gene Symbol | Gene ID | Protein Name | Category | Mechanism | Ref. |
|---|---|---|---|---|---|
| Regulation of HIF2α transcription | |||||
| E2F1# | 1869 | Transcription factor E2F1 | 1 | Transcriptional activation of EPAS1 | [24] |
| OTUD7B# | 56957 | OTU domain-containing protein 7B (Cezanne) | 2 | Transcriptional activation of EPAS1 | [24] |
| ZC3H12A# | 80149 | MCP-induced protein 1 (MCPIP1) | 1 | Negative transcriptional regulation of EPAS1 | [25] |
| SP1# | 6667 | Transcription factor Sp1 | 1 | Transcriptional activation of EPAS1 | [26] |
| SP3# | 6670 | Transcription factor Sp3 | 1 | Transcriptional activation of EPAS1 | [26] |
| PI3K# | Phosphatidylinositol 3-kinase | 2 | Transcriptional activation of EPAS1 | [27] | |
| MTOR# | 2475 | Serine/threonine-protein kinase mTOR | 2 | Transcriptional activation of EPAS1 | [27] |
| DNMT1# | 1786 | DNA (cytosine-5)-methyltransferase 1 (DNMT1) | 1 | Negative transcriptional regulation of EPAS1 | [29] |
| HIF2α protein modifications and protein interactions | |||||
| HIF1AN | 55662 | Hypoxia-inducible factor 1-alpha inhibitor (FIH-1) | 1 | Hydroxylation | [22] |
| CSNK2A1 | 1457 | Casein kinase II subunit alpha (CKIIα) | 1 | Phosphorylation | [30] |
| CSNK1D# | 1453 | Casein kinase I isoform delta (CKIδ) | 1 | Phosphorylation and nuclear transport | [32] |
| MAPK 1/3# | 5594/5595 | Mitogen-activated protein kinase 1/3 (MAPK 1/3) | 1 | Phosphorylation and nuclear transport | [31] |
| SIRT1# | 23411 | NAD-dependent protein deacetylase sirtuin-1 (hSIRT1) | 1 | Deacetylation | [33] |
| SENP1# | 29843 | Sentrin-specific protease 1 | 1 | Desumoylation | [40] |
| RNF4# | 6047 | E3 ubiquitin-protein ligase RNF4 | 1 | Ubiquitination | [40] |
| USP8# | 9101 | Ubiquitin carboxyl-terminal hydrolase 8 (USP8) | 1 | Deubiquitination | [46] |
| USP29 | 57663 | Ubiquitin carboxyl-terminal hydrolase 29 (USP29) | 1 | Deubiquitination | [47] |
| IKBKG | 8517 | NF-kappa-B essential modulator (NEMO) | 1 | Other interaction proteins | [22] |
| KLHL20# | 27252 | Kelch-like protein 20 (KLHL20) | 1 | Other interaction proteins | [48] |
| SART1 (HAF) | 9092 | Hypoxia-associated factor (HAF) | 1 | Other interaction proteins | [49] |
| BHLHE41 (SHARP1) | 79365 | Class E basic helix-loop-helix protein 41 (bHLHe41) | 4 | Other interaction proteins | [50] |
| EIF3E (INT6) | 3646 | Eukaryotic translation initiation factor 3 subunit E (eIF3e) | 1 | Other interaction proteins | [52] |
| PHD2 protein modifications and protein interactions | |||||
| FKBP8 | 23770 | Peptidyl-prolyl cis-trans isomerase FKBP8 (PPIase FKBP8) | 1 | Other interaction proteins | [53] |
| OS-9 | 10956 | Protein OS-9 (OS-9) | Other interaction proteins | [54] | |
| CBFA2T3 (MTG16)# | 863 | Protein CBFA2T3 (CBFA2T3) | 1 | Other interaction proteins | [55] |
| PTGES3 (P23) | 10728 | Prostaglandin E synthase 3 (PTGES3) | 1 | Other interaction proteins | [55] |
| LIMD1 | 8994 | LIM domain-containing protein 1 | 1, 4 | Other interaction proteins | [57,58] |
| VHL protein modifications and protein interactions | |||||
| LIMD1 | 8994 | LIM domain-containing protein 1 | 1, 4 | Other interaction proteins | [58] |
| DNMT3A | 1788 | DNA (cytosine-5)-methyltransferase 3A (DNMT3a) | 1 | MethylationNegative transcriptional regulation of VHL | [35] |
| DNMT3B | 1789 | DNA (cytosine-5)-methyltransferase 3B (DNMT3b) | 1 | MethylationNegative transcriptional regulation of VHL | [35] |
| UBE2S | 27338 | Ubiquitin-conjugating enzyme E2 S (E2-EPF) | 1, 4 | UbiquitinationDegradation of VHL | [45] |
| PIAS4 | 51588 | E3 SUMO-protein ligase PIAS4 (PIASY) | 1, 4 | Sumoylation | [39] |
| CDH1 | 999 | Cadherin 1 | 1 | UbiquitylationDegradation of VHL | [43] |
| WSB1 | 26118 | WD repeat and SOCS box-containing protein 1 (WSB1) | 1 | UbiquitinationDegradation of VHL | [44] |
| SMURF1 | 57154 | E3 ubiquitin-protein ligase SMURF1 (hSMURF1) | 1 | UbiquitinationDegradation of VHL | [42] |
| USP9X | 8239 | Probable ubiquitin carboxyl-terminal hydrolase FAF-X | 2, 4 | DeubiquitylationDegradation of VHL | [42] |
| ELOB | 6923 | Elongin-B (ELOB) | 2 | Other interaction proteinsUbiquitination of HIFα | [59] |
| ELOC | 6921 | Elongin-C (ELOC) | 1 | Other interaction proteinsUbiquitination of HIFα | [59] |
| CUL2 | 8453 | Cullin-2 (CUL-2) | 2 | Other interaction proteinsUbiquitination of HIFα | [59] |
| RBX1 | 9978 | E3 ubiquitin-protein ligase RBX1 | 2 | Other interaction proteinsUbiquitination of HIFα | [59] |
| SAT2 | 112483 | Thialysine N-epsilon-acetyltransferase (SSAT-2) | 1, 4 | Other interaction proteins | [60] |
| TCP1 | 6950 | T-complex protein 1 subunit alpha (TRiC) | 1 | Other interaction proteins | [61] |
| ID2# | 3398 | DNA-binding protein inhibitor ID-2 (ID2) | 1 | Other interaction proteins | [62] |
| FIH protein modifications and protein interactions | |||||
| SIAH1 | 6477 | E3 ubiquitin-protein ligase SIAH1 | 2 | Ubiquitination | [22] |
| NFKBIA | 4792 | NF-kappa-B inhibitor alpha | 2 | Other interaction proteins | [22] |
| NFKB1 | 4790 | Nuclear factor NF-kappa-B p105 subunit | 2 | Other interaction proteins | [22] |
| NOTCH1/2/3 | 4851/4853/4854 | Neurogenic locus notch homolog protein 1/2/3 (Notch 1/2/3) | 2 | Other interaction proteins | [22] |
| ASB4 | 51666 | Ankyrin repeat and SOCS box protein 4 (ASB-4) | 2 | Other interaction proteins | [22] |
| CUX1 | 1523 | Homeobox protein cut-like 1 | 2 | DNA protein partner | [63] |
| PRKCZ | 5590 | Protein kinase C zeta type | 3 | Other interaction proteins | [63] |
| ARNT protein modifications and protein interactions | |||||
| UBE2I# | 7329 | SUMO-conjugating enzyme UBC9 | 2 | Sumoylation | [36] |
| PIAS1# | 8554 | E3 SUMO-protein ligase PIAS1 | 2 | Sumoylation | [36] |
| AHR# | 196 | Aryl hydrocarbon receptor (AHR) | 2 | DNA protein partner | [64] |
| TACC3 | 10460 | Transforming acidic coiled-coil-containing protein 3 | 2 | DNA protein partnerCo-activator of HIF2 complex | [65] |
| Nuclear transport of key players of oxygen-sensing pathway | |||||
| KPNA2# | 3838 | Importin subunit alpha-1 (Importinα) | 1 | Nuclear import of HIF2α, ARNT | [68] |
| KPNB1# | 3837 | Importin subunit beta-1 (Importinβ) | 1 | Nuclear transport of HIF2α, ARNT | [68] |
| XPO1 | 7514 | Exportin-1 (CRM1) | 1 | Nuclear export of HIF2α, PHD2 | [31,66] |
| EEF1A1 | 1915 | Elongation factor 1-alpha 1 (eEF1A) | 1 | Nuclear export of VHL | [69] |
| Regulation of EPO transcription | |||||
| EP300 | 2033 | Histone acetyltransferase p300 (p300) | 1 | Transcriptional activation of EPO (co-activator) | [15,71] |
| CREBBP | 1387 | CREB-binding protein (CBP) | 1 | Transcriptional activation of EPO (co-activator) | [15] |
| HNF4A | 3172 | Hepatocyte nuclear factor-4α (HNF4α) | 1 | Transcriptional activation of EPO | [72] |
| RXRA | 6256 | Retinoid X receptor alpha (RXRα) | 1 | Transcriptional activation of EPO | [73] |
| NCOA3 (SRC3)# | 8202 | Nuclear receptor coactivator 3 (NCoA-3) | 1, 4 | DNA protein partner | [71] |
| RELA | 5970 | Transcription factor p65 (NFKB3) | 1, 4 | Transcriptional activation of EPO | [74,75] |
| GATA2 | 2624 | Endothelial transcription factor GATA-2 (GATA-2) | 1 | Negative transcriptional regulation of EPO | [77] |
| GATA3 | 2625 | Trans-acting T-cell-specific transcription factor GATA-3 (GATA-3) | 1 | Negative transcriptional regulation of EPO | [77] |
| MECP2 | 4204 | Methyl- CpG binding protein 2 (MeCP2) | 1 | Negative transcriptional regulation of EPO | [78] |
| p300 protein modifications and protein interactions | |||||
| SIRT1 | 23411 | NAD-dependent protein deacetylase sirtuin-1 (hSIRT1) | 2 | Deaceltylation | [34] |
| UBE2I | 7329 | SUMO-conjugating enzyme UBC9 | 2 | Sumoylation | [37] |
| SENP3 | 26168 | Sentrin-specific protease 3 | 2 | Desumoylation | [41] |
| CBP protein modifications and protein interactions | |||||
| UBE2I | 7329 | SUMO-conjugating enzyme UBC9 | 2 | Sumoylation | [38] |
3. Diseases and Putative Treatments
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haase, V.H. Regulation of Erythropoiesis by Hypoxia-Inducible Factors. Blood Rev. 2013, 27, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Shih, H.M.; Wu, C.J.; Lin, S.L. Physiology and Pathophysiology of Renal Erythropoietin-Producing Cells. J. Formos. Med. Assoc. 2018, 117, 955–963. [Google Scholar] [CrossRef]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING V10: Protein-Protein Interaction Networks, Integrated over the Tree of Life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Consortium, T.U. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Iny Stein, T.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinf. 2016, 54, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a Knowledge-Based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- NCBI Resource Coordinators. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef] [Green Version]
- Tweedie, S.; Braschi, B.; Gray, K.; Jones, T.E.M.; Seal, R.L.; Yates, B.; Bruford, E.A. Genenames.Org: The HGNC and VGNC Resources in 2021. Nucleic Acids Res. 2021, 49, D939–D946. [Google Scholar] [CrossRef] [PubMed]
- Jelkmann, W. Erythropoietin: Structure, Control of Production, and Function. Physiol. Rev. 1992, 72, 449–489. [Google Scholar] [CrossRef]
- Suzuki, N.; Yamamoto, M. Roles of Renal Erythropoietin-Producing (REP) Cells in the Maintenance of Systemic Oxygen Homeostasis. Pflugers Arch. Eur. J. Physiol. 2016, 468, 3–12. [Google Scholar] [CrossRef]
- Kuhrt, D.; Wojchowski, D.M. Emerging EPO and EPO Receptor Regulators and Signal Transducers. Blood 2015, 125, 3536–3541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L. The Genomics and Genetics of Oxygen Homeostasis. Annu. Rev. Genom. Hum. Genet. 2020, 21, 183–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fandrey, J. Oxygen-Dependent and Tissue-Specific Regulation of Erythropoietin Gene Expression. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286. [Google Scholar] [CrossRef]
- Dengler, V.L.; Galbraith, M.; Espinosa, J.M. Transcriptional Regulation by Hypoxia Inducible Factors. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, V. The VHL Tumor Suppressor: Master Regulator of HIF. Curr. Pharm. Des. 2009, 15, 3895–3903. [Google Scholar] [CrossRef] [Green Version]
- Bunn, H.F. Erythropoietin. Cold Spring Harb. Perspect. Med. 2013, 3, a011619. [Google Scholar] [CrossRef] [Green Version]
- Souma, T.; Nezu, M.; Nakano, D.; Yamazaki, S.; Hirano, I.; Sekine, H.; Dan, T.; Takeda, K.; Fong, G.H.; Nishiyama, A.; et al. Erythropoietin Synthesis in Renal Myofibroblasts Is Restored by Activation of Hypoxia Signaling. J. Am. Soc. Nephrol. 2016, 27, 428–438. [Google Scholar] [CrossRef]
- Saito, T.; Kawaguchi, H. HIF-2α as a Possible Therapeutic Target of Osteoarthritis. Osteoarthr. Cartil. 2010, 18, 1552–1556. [Google Scholar] [CrossRef] [Green Version]
- Stolze, I.; Berchner-Pfannschmidt, U.; Freitag, P.; Wotzlaw, C.; Rössler, J.; Frede, S.; Acker, H.; Fandrey, J. Hypoxia-Inducible Erythropoietin Gene Expression in Human Neuroblastoma Cells. Blood 2002, 100, 2623–2628. [Google Scholar] [CrossRef]
- Suzuki, N.; Gradin, K.; Poellinger, L.; Yamamoto, M. Regulation of Hypoxia-Inducible Gene Expression after HIF Activation. Exp. Cell Res. 2017, 356, 182–186. [Google Scholar] [CrossRef]
- Lisy, K.; Peet, D.J. Turn Me on: Regulating HIF Transcriptional Activity. Cell Death Differ. 2008, 15, 642–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelkmann, W. Regulation of Erythropoietin Production. J. Physiol. 2011, 589, 1251–1258. [Google Scholar] [CrossRef]
- Moniz, S.; Bandarra, D.; Biddlestone, J.; Campbell, K.J.; Komander, D.; Bremm, A.; Rocha, S. Cezanne Regulates E2F1-Dependent HIF2α Expression. J. Cell Sci. 2015, 128, 3082–3093. [Google Scholar] [CrossRef] [Green Version]
- Ligęza, J.; Marona, P.; Gach, N.; Lipert, B.; Miekus, K.; Wilk, W.; Jaszczyński, J.; Stelmach, A.; Loboda, A.; Dulak, J.; et al. MCPIP1 Contributes to Clear Cell Renal Cell Carcinomas Development. Angiogenesis 2017, 20. [Google Scholar] [CrossRef] [Green Version]
- Wada, T.; Shimba, S.; Tezuka, M. Transcriptional Regulation of the Hypoxia Inducible Factor-2α (HIF-2α) Gene during Adipose Differentiation in 3T3-L1 Cells. Biol. Pharm. Bull. 2006, 29, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohlin, S.; Hamidian, A.; Von Stedingk, K.; Bridges, E.; Wigerup, C.; Bexell, D.; Påhlman, S. PI3K-MTORC2 but Not PI3K-MTORC1 Regulates Transcription of HIF2A/EPAS1and Vascularization in Neuroblastoma. Cancer Res. 2015, 75, 4617–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamidian, A.; Vaapil, M.; von Stedingk, K.; Fujita, T.; Persson, C.U.; Eriksson, P.; Veerla, S.; De Preter, K.; Speleman, F.; Fujii, H.; et al. Promoter-Associated Proteins of EPAS1 Identified by EnChIP-MS—A Putative Role of HDX as a Negative Regulator. Biochem. Biophys. Res. Commun. 2018, 499, 291–298. [Google Scholar] [CrossRef]
- Xu, X.H.; Bao, Y.; Wang, X.; Yan, F.; Guo, S.; Ma, Y.; Xu, D.; Jin, L.; Xu, J.; Wang, J. Hypoxic-Stabilized EPAS1 Proteins Transactivate DNMT1 and Cause Promoter Hypermethylation and Transcription Inhibition of EPAS1 in Non-Small Cell Lung Cancer. FASEB J. 2018, 32, 6694–6705. [Google Scholar] [CrossRef] [Green Version]
- Gradin, K.; Takasaki, C.; Fujii-kuriyama, Y.; Sogawa, K. The Transcriptional Activation Function of the HIF-like Factor Requires Phosphorylation at a Conserved Threonine. J. Biol. Chem. 2002, 277, 23508–23514. [Google Scholar] [CrossRef] [Green Version]
- Gkotinakou, I.M.; Befani, C.; Simos, G.; Liakos, P. ERK1/2 Phosphorylates HIF-2α and Regulates Its Activity by Controlling Its CRM1-Dependent Nuclear Shuttling. J. Cell Sci. 2019, 132, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pangou, E.; Befani, C.; Mylonis, I.; Samiotaki, M.; Panayotou, G.; Simos, G.; Liakos, P. HIF-2α Phosphorylation by CK1δ Promotes Erythropoietin Secretion in Liver Cancer Cells under Hypoxia. J. Cell Sci. 2016, 129, 4213–4226. [Google Scholar] [CrossRef] [Green Version]
- Dioum, E.M.; Chen, R.; Alexander, M.S.; Zhang, Q.; Hogg, R.T.; Gerard, R.D.; Garcia, J.A. Regulation of Hypoxia-Inducible Factor 2α Signaling by the Stress-Responsive Deacetylase Sirtuin 1. Science 2009, 324, 1289–1293. [Google Scholar] [CrossRef]
- Bouras, T.; Fu, M.; Sauve, A.A.; Wang, F.; Quong, A.A.; Perkins, N.D.; Hay, R.T.; Gu, W.; Pestell, R.G. SIRT1 Deacetylation and Repression of P300 Involves Lysine Residues 1020/1024 within the Cell Cycle Regulatory Domain 1. J. Biol. Chem. 2005, 280, 10264–10276. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Zhu, M.; Song, K.; Wuren, T.; Yan, J.; Ge, R.-L.; Ji, L.; Cui, S. VHL Gene Methylation Contributes to Excessive Erythrocytosis in Chronic Mountain Sickness Rat Model by Upregulating the HIF-2α/EPO Pathway. Life Sci. 2021, 266, 118873. [Google Scholar] [CrossRef] [PubMed]
- Tojo, M.; Matsuzaki, K.; Minami, T.; Honda, Y.; Yasuda, H.; Chiba, T.; Saya, H.; Fujii-Kuriyama, Y.; Nakao, M. The Aryl Hydrocarbon Receptor Nuclear Transporter Is Modulated by the SUMO-1 Conjugation System. J. Biol. Chem. 2002, 277, 46576–46585. [Google Scholar] [CrossRef] [Green Version]
- Girdwood, D.; Bumpass, D.; Vaughan, O.A.; Thain, A.; Anderson, L.A.; Snowden, A.W.; Garcia-Wilson, E.; Perkins, N.D.; Hay, R.T. P300 Transcriptional Repression Is Mediated by SUMO Modification. Mol. Cell 2003, 11, 1043–1054. [Google Scholar] [CrossRef]
- Park, S.; Stanfield, R.L.; Martinez-Yamout, M.A.; Dyson, H.J.; Wilson, I.A.; Wright, P.E. Role of the CBP Catalytic Core in Intramolecular SUMOylation and Control of Histone H3 Acetylation. Proc. Natl. Acad. Sci. USA 2017, 114, E5335–E5342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Verma, S.C.; Kumar, P.; Ma, M.; Robertson, E.S. Hypoxia Inactivates the VHL Tumor Suppressor through PIASy-Mediated SUMO Modification. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hagen, M.; Overmeer, R.M.; Abolvardi, S.S.; Vertegaal, A.C.O. RNF4 and VHL Regulate the Proteasomal Degradation of SUMO-Conjugated Hypoxia-Inducible Factor-2α. Nucleic Acids Res. 2009, 38, 1922–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Han, Y.; Wang, Y.; Sun, X.; Yan, S.; Yeh, E.T.H.; Chen, Y.; Cang, H.; Li, H.; Shi, G.; et al. SENP3 Is Responsible for HIF-1 Transactivation under Mild Oxidative Stress via P300 de-SUMOylation. EMBO J. 2009, 28, 2748–2762. [Google Scholar] [CrossRef] [Green Version]
- Kubaichuk, K.; Kietzmann, T. Involvement of E3 Ligases and Deubiquitinases in the Control of HIF-α Subunit Abundance. Cells 2019, 8, 598. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Xin, H.; Eckert, D.T.; Brown, J.A.; Gnarra, J.R. Hypoxia and Cell Cycle Regulation of the von Hippel-Lindau Tumor Suppressor. Oncogene 2011, 30, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Lee, S.B.; Jang, J.; Yi, S.Y.; Kim, S.H.; Han, S.A.; Lee, J.M.; Tong, S.Y.; Vincelette, N.D.; Gao, B.; et al. WSB1 Promotes Tumor Metastasis by Inducing PVHL Degradation. Genes Dev. 2015, 29, 2244–2257. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.H.; Kim, D.G.; Yu, D.Y.; Kang, H.M.; Noh, K.H.; Kim, D.S.; Park, D.; Chang, T.K.; Im, D.S.; Jung, C.R. Stabilization of E2-EPF UCP Protein Is Implicated in Hepatitis B Virus-Associated Hepatocellular Carcinoma Progression. Cell. Mol. Life Sci. 2019, 76, 2647–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troilo, A.; Alexander, I.; Muehl, S.; Jaramillo, D.; Knobeloch, K.; Krek, W. HIF1α Deubiquitination by USP8 Is Essential for Ciliogenesis in Normoxia. EMBO Rep. 2014, 15, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schober, A.; Martin-Barros, I.; Martin-Mateos, T.; Perez-Andres, E.; Carlevaris, O.; Pozo, S.; Cortazar, A.; Aransay, A.; Carracedo, A.; Mayor, U.; et al. USP29 Is a Novel Non-Canonical Hypoxia Inducible Factor-α Activator. bioRxiv 2020. [Google Scholar] [CrossRef]
- Higashimura, Y.; Terai, T.; Yamaji, R.; Mitani, T.; Ogawa, M.; Harada, N.; Inui, H.; Nakano, Y. Kelch-like 20 up-Regulates the Expression of Hypoxia-Inducible Factor-2α through Hypoxia- and von Hippel-Lindau Tumor Suppressor Protein-Independent Regulatory Mechanisms. Biochem. Biophys. Res. Commun. 2011, 413, 201–205. [Google Scholar] [CrossRef]
- Hashimoto, T.; Shibasaki, F. Hypoxia-Inducible Factor as an Angiogenic Master Switch. Front. Pediatr. 2015, 3, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Montagner, M.; Enzo, E.; Forcato, M.; Zanconato, F.; Parenti, A.; Rampazzo, E.; Basso, G.; Leo, G.; Rosato, A.; Bicciato, S.; et al. SHARP1 Suppresses Breast Cancer Metastasis by Promoting Degradation of Hypoxia-Inducible Factors. Nature 2012, 487, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Camps, C.; Petousi, N.; Bento, C.; Cario, H.; Copley, R.R.; McMullin, M.F.; Van Wijk, R.; Ratcliffe, P.J.; Robbins, P.A.; Taylor, J.C. Gene Panel Sequencing Improves the Diagnostic Work-up of Patients with Idiopathic Erythrocytosis and Identifies New Mutations. Haematologica 2016, 101, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Uchida, K.; Endler, A.; Shibasak, F. Mammalian Tumor Suppressor Int6 Specifically Targets Hypoxia Inducible Factor 2 Alpha for Degrada- Tion by Hypoxia- and PVHL-Independent Regulation. Chem. J. Biol. 2007, 282, 12707–12716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, S.; Nesper, J.; Hasgall, P.A.; Wirthner, R.; Nytko, K.J.; Edlich, F.; Katschinski, D.M.; Stiehl, D.P.; Wenger, R.H.; Camenisch, G. The Peptidyl Prolyl Cis/Trans Isomerase FKBP38 Determines Hypoxia-Inducible Transcription Factor Prolyl-4-Hydroxylase PHD2 Protein Stability. Mol. Cell. Biol. 2007, 27, 3758–3768. [Google Scholar] [CrossRef] [Green Version]
- Baek, J.H.; Mahon, P.C.; Oh, J.; Kelly, B.; Krishnamachary, B.; Pearson, M.; Chan, D.A.; Giaccia, A.J.; Semenza, G.L. OS-9 Interacts with Hypoxia-Inducible Factor 1α and Prolyl Hydroxylases to Promote Oxygen-Dependent Degradation of HIF-1α. Mol. Cell 2005, 17, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Gullberg, U.; Olsson, I.; Ajore, R. Myeloid Translocation Gene-16 Co-Repressor Promotes Degradation of Hypoxia-Inducible Factor 1. PLoS ONE 2015, 10, e0123725. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Li, L.S.; Heaton-Johnson, K.J.; Arsenault, P.R.; Master, S.R.; Lee, F.S. Prolyl Hydroxylase Domain Protein 2 (PHD2) Binds a Pro-Xaa-Leu-Glu Motif, Linking It to the Heat Shock Protein 90 Pathway. J. Biol. Chem. 2013, 288, 9662–9674. [Google Scholar] [CrossRef] [Green Version]
- Minervini, G.; Quaglia, F.; Tosatto, S.C.E. Computational Analysis of Prolyl Hydroxylase Domain-Containing Protein 2 (PHD2) Mutations Promoting Polycythemia Insurgence in Humans. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Foxler, D.E.; Bridge, K.S.; James, V.; Webb, T.M.; Mee, M.; Wong, S.C.K.; Feng, Y.; Constantin-Teodosiu, D.; Petursdottir, T.E.; Bjornsson, J.; et al. The LIMD1 Protein Bridges an Association between the Prolyl Hydroxylases and VHL to Repress HIF-1 Activity. Nat. Cell Biol. 2012, 14, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Cardote, T.A.F.; Gadd, M.S.; Ciulli, A. Crystal Structure of the Cul2-Rbx1-EloBC-VHL Ubiquitin Ligase Complex. Structure 2017, 25, 901–911. [Google Scholar] [CrossRef] [Green Version]
- Baek, J.H.; Liu, Y.V.; McDonald, K.R.; Wesley, J.B.; Hubbi, M.E.; Byun, H.; Semenza, G.L. Spermidine/Spermine-N1-Acetyltransferase 2 Is an Essential Component of the Ubiquitin Ligase Complex That Regulates Hypoxia-Inducible Factor 1alpha. J. Biol. Chem. 2007, 282, 23572–23580. [Google Scholar] [CrossRef] [Green Version]
- Gossage, L.; Eisen, T.; Maher, E.R. VHL, the Story of a Tumour Suppressor Gene. Nat. Rev. Cancer 2015, 15, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Frattini, V.; Bansal, M.; Castano, A.; Sherman, D.; Hutchinson, K.; Bruce, J.; Califano, A.; Liu, G.; Cardozo, T.; et al. An ID2-Dependent Mechanism for VHL Inactivation in Cancer. Nature 2016, 529, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, E.; Dutta, S.; Lau, J.S.; Jiang, S.; Datta, K.; Mukhopadhyay, D. Protein Kinase C-Mediated Modulation of FIH-1 Expression by the Homeodomain Protein CDP/Cut/Cux. Mol. Cell. Biol. 2007, 27, 7345–7353. [Google Scholar] [CrossRef] [Green Version]
- Asai, H.; Hirata, J.; Hirano, A.; Hirai, K.; Seki, S.; Watanabe-Akanuma, M. Activation of Aryl Hydrocarbon Receptor Mediates Suppression of Hypoxia-Inducible Factor-Dependent Erythropoietin Expression by Indoxyl Sulfate. Am. J. Physiol. Cell Physiol. 2016, 310, C142–C150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partch, C.L.; Gardner, K.H. Coactivators Necessary for Transcriptional Output of the Hypoxia Inducible Factor, HIF, Are Directly Recruited by ARNT PAS-B. Proc. Natl. Acad. Sci. USA 2011, 108, 7739–7744. [Google Scholar] [CrossRef] [Green Version]
- Depping, R.; Jelkmann, W.; Kosyna, F.K. Nuclear-Cytoplasmatic Shuttling of Proteins in Control of Cellular Oxygen Sensing. J. Mol. Med. 2015, 93, 599–608. [Google Scholar] [CrossRef]
- Tran, E.J.; King, M.C.; Corbett, A.H. Macromolecular Transport between the Nucleus and the Cytoplasm: Advances in Mechanism and Emerging Links to Disease. Biochim. Biophys. Acta 2014, 1843, 2784–2795. [Google Scholar] [CrossRef] [Green Version]
- Depping, R.; Steinhoff, A.; Schindler, S.G.; Friedrich, B.; Fagerlund, R.; Metzen, E.; Hartmann, E.; Köhler, M. Nuclear Translocation of Hypoxia-Inducible Factors (HIFs): Involvement of the Classical Importin α/β Pathway. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Khacho, M.; Mekhail, K.; Pilon-Larose, K.; Pause, A.; Cote, J.; Lee, S. EEF1A Is a Novel Component of the Mammalian Nuclear Protein Export Machinery. Mol. Biol. Cell 2008, 19, 5296–5308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, B.L.; Bunn, H.F. Regulation of the Erythropoietin Gene. Blood 1999, 94, 1864–1877. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, R.; Wu, X.; Hankinson, O. Roles of Coactivators in Hypoxic Induction of the Erythropoietin Gene. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galson, D.L.; Tsuchiya, T.; Tendler, D.S.; Huang, L.E.; Ren, Y.; Ogura, T.; Bunn, H.F. The Orphan Receptor Hepatic Nuclear Factor 4 Functions as a Transcriptional Activator for Tissue-Specific and Hypoxia-Specific Erythropoietin Gene Expression and Is Antagonized by EAR3/COUP-TF1. Mol. Cell. Biol. 1995, 15, 2135–2144. [Google Scholar] [CrossRef] [Green Version]
- Makita, T.; Hernandez-Hoyos, G.; Chen, T.H.P.; Wu, H.; Rothenberg, E.V.; Sucov, H.M. A Developmental Transition in Definitive Erythropoiesis: Erythropoietin Expression Is Sequentially Regulated by Retinoic Acid Receptors and HNF4. Genes Dev. 2001, 15, 889–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa, Y.G.; Chan, A.K.; Ibrahim, R.; Tang, Y.; Burow, M.E.; Alam, J.; Scandurro, A.B.; Beckman, B.S. NF-ΚB Plays a Key Role in Hypoxia-Inducible Factor-1-Regulated Erythropoietin Gene Expression. Exp. Hematol. 2002, 30, 1419–1427. [Google Scholar] [CrossRef]
- Vočanec, D.; Prijatelj, T.; Debeljak, N.; Kunej, T. Genetic Variants of Erythropoietin (EPO) and EPO Receptor Genes in Familial Erythrocytosis. Int. J. Lab. Hematol. 2019, 41, 162–167. [Google Scholar] [CrossRef] [Green Version]
- Imagawa, S.; Yamamoto, M.; Miura, Y. Negative Regulation of the Erythropoietin Gene Expression by the GATA Transcription Factors. Blood 1997, 89, 1430–1439. [Google Scholar] [CrossRef]
- Obara, N.; Suzuki, N.; Kim, K.; Nagasawa, T.; Imagawa, S.; Yamamoto, M. Repression via the GATA Box Is Essential for Tissue-Specific Erythropoietin Gene Expression. Blood 2008, 111, 5223–5232. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Blachard, K.L. Erratum: DNA Methylation Represses the Expression of the Human Erythropoietin Gene by Two Different Mechanisms. Blood 2000, 95, 111–119. [Google Scholar] [CrossRef]
- Hodges, V.M.; Rainey, S.; Lappin, T.R.; Maxwell, A.P. Pathophysiology of Anemia and Erythrocytosis. Crit. Rev. Oncol. Hematol. 2007, 64, 139–158. [Google Scholar] [CrossRef]
- Kim, A.R.; Ulirsch, J.C.; Wilmes, S.; Unal, E.; Moraga, I.; Karakukcu, M.; Yuan, D.; Kazerounian, S.; Abdulhay, N.J.; King, D.S.; et al. Functional Selectivity in Cytokine Signaling Revealed Through a Pathogenic EPO Mutation. Cell 2017, 168, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- Bento, C.; Cario, H.; Gardie, B.; Sylvie, H.; McMullin, M.F. Congenital Erythrocytosis and Hereditary Thrombocytosis. In Clinical Presentation, Diagnosis, Treatment and Follow-up: A Practical Guide with Clinical Cases; European Cooperation in Science & Technology: Brussels, Belgium, 2015; pp. 41–61. [Google Scholar]
- Lappin, T.R.; Lee, F.S. Update on Mutations in the HIF: EPO Pathway and Their Role in Erythrocytosis. Blood Rev. 2019, 37, 100590. [Google Scholar] [CrossRef]
- Zatyka, M.; Morrissey, C.; Kuzmin, I.; Lerman, M.I.; Latif, F.; Richards, F.M.; Maher, E.R. Genetic and Functional Analysis of the von Hippel-Lindau (VHL) Tumour Suppressor Gene Promoter. J. Med. Genet. 2002, 39, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yang, H. The Roles of VHL-Dependent Ubiquitination in Signaling and Cancer. Front. Oncol. 2012, 2, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L. Signal Transduction to Hypoxia-Inducible Factor 1. Biochem. Pharmacol. 2002, 64, 993–998. [Google Scholar] [CrossRef]
- Fallah, J.; Rini, B.I. HIF Inhibitors: Status of Current Clinical Development. Curr. Oncol. Rep. 2019, 21, 1–10. [Google Scholar] [CrossRef]
- Klein, M.; Kaestner, L.; Bogdanova, A.Y.; Minetti, G.; Rudloff, S.; Lundby, C.; Makhro, A.; Seiler, E.; van Cromvoirt, A.; Fenk, S.; et al. Absence of Neocytolysis in Humans Returning from a 3-Week High-Altitude Sojourn. Acta Physiol. 2021, 1–11. [Google Scholar] [CrossRef]
- Mairbäurl, H. Neocytolysis: How to Get Rid of the Extra Erythrocytes Formed by Stress Erythropoiesis upon Descent from High Altitude. Front. Physiol. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Chen, W.; Hill, H.; Christie, A.; Kim, M.S.; Holloman, E.; Pavia-Jimenez, A.; Homayoun, F.; Ma, Y.; Patel, N.; Yell, P.; et al. Targeting Renal Cell Carcinoma with a HIF-2 Antagonist. Nature 2016, 539, 112–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bei, Y. On-Target Efficacy of a HIF2α Antagonist in Preclinical Kidney Cancer Models. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Fath, D.M.; Kong, X.; Liang, D.; Lin, Z.; Chou, A.; Jiang, Y.; Fang, J.; Caro, J.; Sang, N. Histone Deacetylase Inhibitors Repress the Transactivation Potential of Hypoxia-Inducible Factors Independently of Direct Acetylation of HIF-α. J. Biol. Chem. 2006, 281, 13612–13619. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, M.C.; Zhang, D.L.; Ollivierre, W.H.; Noguchi, A.; Springer, D.A.; Linehan, W.M.; Rouault, T.A. Therapeutic Inhibition of HIF-2α Reverses Polycythemia and Pulmonary Hypertension in Murine Models of Human Diseases. Blood 2021, 137, 2509–2519. [Google Scholar] [CrossRef] [PubMed]
- Kaelin, W.G.; Ratcliffe, P.J.; Semenza, G.L. Pathways for Oxygen Regulation and Homeostasis: The 2016 Albert Lasker Basic Medical Research Award. JAMA J. Am. Med. Assoc. 2016, 316, 1252–1253. [Google Scholar] [CrossRef] [PubMed]
- Rainville, N.; Jachimowicz, E.; Wojchowski, D.M. Targeting EPO and EPO RECEPTOR PATHWAYS in Anemia and Dysregulated Erythropoiesis. Expert Opin. Ther. Targets 2016, 20, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Weidemann, A.; Johnson, R.S. Nonrenal Regulation of EPO Synthesis. Kidney Int. 2009, 75, 682–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| Gene Symbol | Gene ID | Protein Name | Mechanism |
|---|---|---|---|
| EPAS1 | 2034 | Hypoxia-inducible factor 2-alpha (HIF2α) | Transcriptional activation of EPO [17] |
| HIF1A | 3091 | Hypoxia-inducible factor 1-alpha (HIF1α) | Transcriptional activation [16] |
| HIF3A | 64344 | Hypoxia-inducible factor 3-alpha (HIF3α) | Transcriptional repression [16] |
| EGLN1 | 54583 | HIF-prolyl hydroxylase 2 (PHD2) | HIF2α hydroxylation [18] |
| VHL | 7428 | von Hippel-Lindau disease tumor suppressor protein (VHL) | HIF2α ubiquitination [16] |
| ARNT | 405 | Aryl hydrocarbon receptor nuclear translocator (ARNT) | HIF2α DNA protein partner Transcriptional activation of EPO [15] |
| ARNTL# | 406 | Aryl hydrocarbon receptor nuclear translocator-like protein 1 (ARNTL) | HIFα DNA protein partner [19] |
| ARNT2# | 9915 | Aryl hydrocarbon receptor nuclear translocator 2 (ARNT2) | HIFα DNA protein partner [20] |
| EPO | 2056 | Erythropoietin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomc, J.; Debeljak, N. Molecular Insights into the Oxygen-Sensing Pathway and Erythropoietin Expression Regulation in Erythropoiesis. Int. J. Mol. Sci. 2021, 22, 7074. https://doi.org/10.3390/ijms22137074
Tomc J, Debeljak N. Molecular Insights into the Oxygen-Sensing Pathway and Erythropoietin Expression Regulation in Erythropoiesis. International Journal of Molecular Sciences. 2021; 22(13):7074. https://doi.org/10.3390/ijms22137074
Chicago/Turabian StyleTomc, Jana, and Nataša Debeljak. 2021. "Molecular Insights into the Oxygen-Sensing Pathway and Erythropoietin Expression Regulation in Erythropoiesis" International Journal of Molecular Sciences 22, no. 13: 7074. https://doi.org/10.3390/ijms22137074
APA StyleTomc, J., & Debeljak, N. (2021). Molecular Insights into the Oxygen-Sensing Pathway and Erythropoietin Expression Regulation in Erythropoiesis. International Journal of Molecular Sciences, 22(13), 7074. https://doi.org/10.3390/ijms22137074